1. Introduction
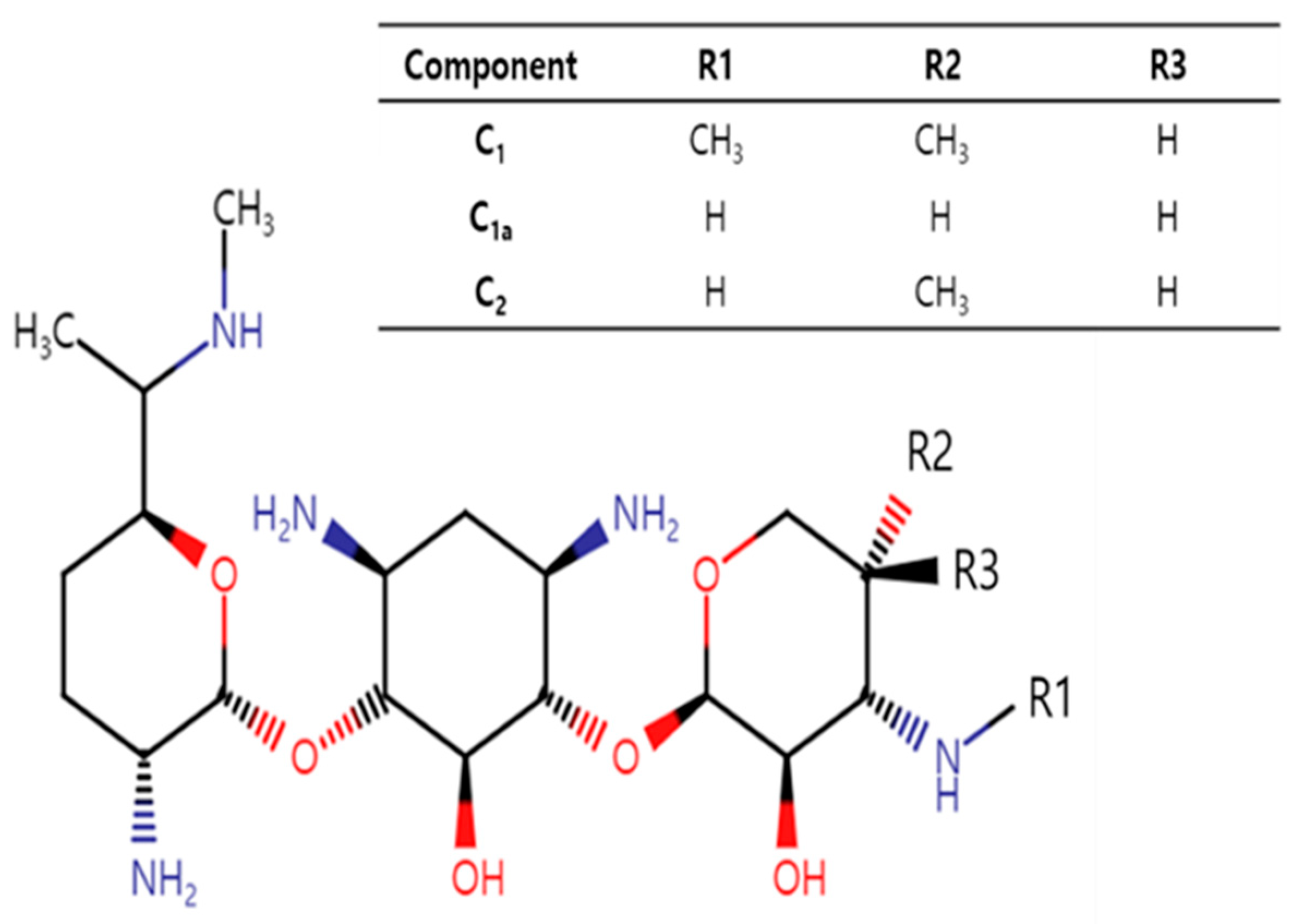
Gentamicin, a bactericidal aminoglycoside antibiotic, is a mixture of pharmacologically active components, mainly C
1, C
1a and C
2 and other minor components or impurities [
1,
2]. The major components differ mainly in their methylation patterns on the 2-amino-hexose ring (
Figure 1). Gentamicin C
1a is devoid of methyl groups on this ring, while both C
1 and C
2 have a methyl group at the 6′ position. C
1 further distinguishes itself by being N-methylated at this position, whereas C
2 retains free amines.
Gentamicin has broad-spectrum activity, showing effectiveness against a wide variety of serious bacterial infections in several animal species that are caused by susceptible Gram-negative and some Gram-positive aerobic bacteria [
3]. The mechanism of action of aminoglycosides involves several phases. These include binding of the drug to the components of bacterial cell membranes, resulting in displacement of divalent cations and increased membrane permeability. This allows for aminoglycoside entry and accumulation in the cell and to its primary intracellular target, the bacterial 30S ribosome, where it results in the mistranslation of proteins. This results in aminoglycoside accumulation, protein mistranslation, and synthesis inhibition, ultimately resulting in concentration-dependent bacterial killing [
4,
5,
6].
Although gentamicin has been well-established in a variety of clinical settings, its use is also associated with adverse effects, as seen with other aminoglycosides, including nephrotoxicity and ototoxicity [
4]. Human and animal studies have indicated that different components of gentamicin possess different potentials for toxicity, with some studies attributing the observed ototoxicity and nephrotoxicity of gentamicin mainly to the C
2 component [
7,
8,
9]. Various levels of the different components contained in different gentamicin formulations, as well as significant differences observed in some pharmacokinetic parameters between different gentamicin components, could have clinical or toxicological implications [
9,
10]. However, reported differences in the pharmacokinetics (PK) of gentamicin components are not consistent across species [
2,
10,
11,
12], emphasizing the importance of conducting PK studies of gentamicin components in species of interest such as piglets.
The PK of gentamicin has been studied in several species, including dogs [
1], horses [
2], chickens [
10], fish [
11], cats [
13], pigs [
14], sheep [
15], calves [
16], and buffalo calves [
17]. Other than a single PK study reported several decades ago that used the intravenous (IV) route of administration [
14], there are no gentamicin PK studies in piglets after the commonly used intramuscular (IM) or oral routes of administration.
Furthermore, most PK studies are conducted in healthy animals, and it is increasingly recognized that disease states—for example, bacterial infection and inflammation-mediated pathophysiological changes [
18]—could affect the pharmacokinetic profile of several drugs to an extent that entails dosage adjustments.
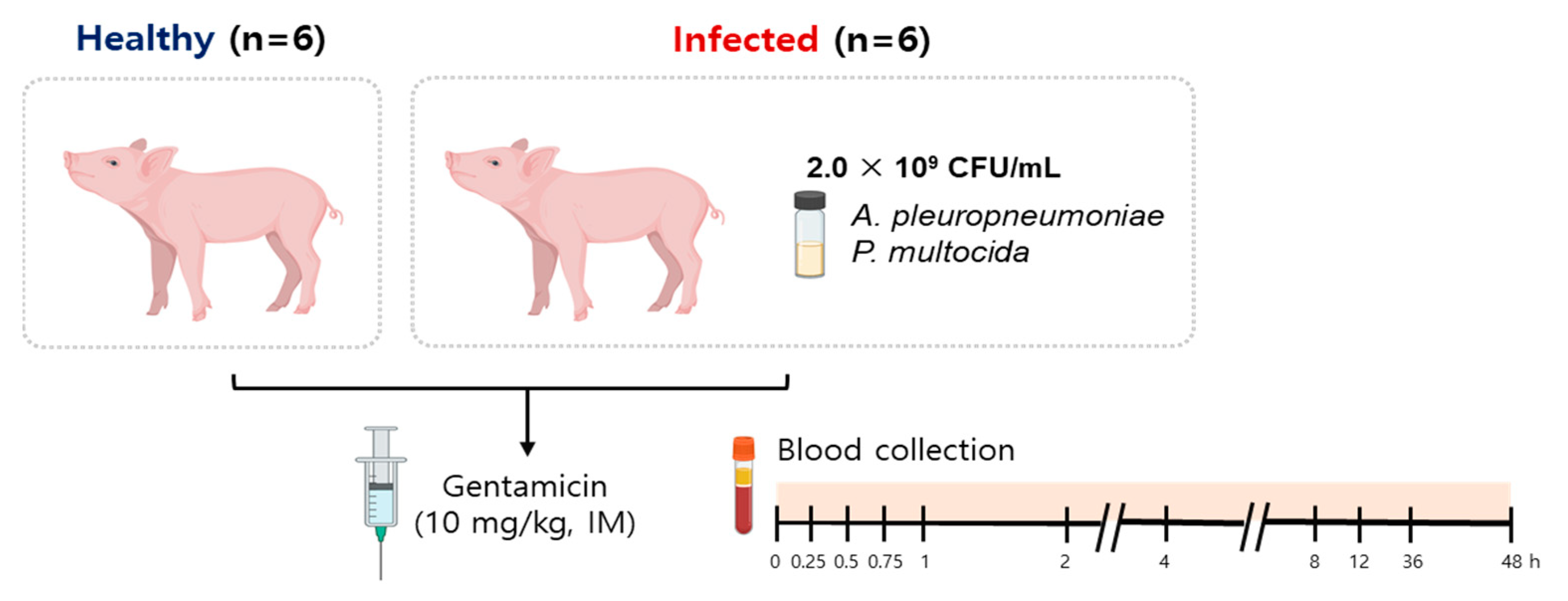
Therefore, we designed the current study to address the following main objectives. Our first objective was to characterize the pharmacokinetics of the gentamicin components C
1, C
1a and C
2 in piglets after the commonly used IM route of administration and fill the knowledge gap in this area. Our second objective was to assess whether the health status of the piglets had any modifying effects on the disposition kinetics of gentamicin and determine the magnitude of any such effects by comparing the gentamicin PKs between healthy and infected piglets. To do so, and because bacterial infections in piglets occur more often as co-infections rather than as single infections [
19], we developed and used a co-infection model induced by intranasal inoculation of
Actinobacillus pleuropneumoniae (AP) and
Pasteurella multocida (PM), which represent two important pathogens that are commonly associated with porcine respiratory disease complex (PRDC) [
20]. Finally, it was our aim to perform limited pharmacokinetic/pharmacodynamic (PK/PD) analyses using gentamicin PK information obtained from the healthy and infected pigs and published minimum inhibitory concentration (MIC) distribution data to gain further understanding of our PK data in the context of the susceptibility patterns of the pathogens. This could assist in making informed decisions during antibiotic and dosage regimen selections.
3. Discussion
Although gentamicin has long been used as a treatment for respiratory diseases, colibacillosis, peritonitis, urinary tract infection and sepsis in piglets [
14,
25], its intramuscular pharmacokinetics in piglets were largely unknown. Furthermore, while within human medicine, it is recognized that the PK of many compounds can be altered by the presence of inflammation or infection, far less is known about these relationships within the framework of veterinary medicine [
26]. Herein, we conducted a comparative PK study of gentamicin after a single intramuscular injection in healthy piglets and piglets co-infected with two important respiratory pathogens that are commonly associated with porcine respiratory disease complex (PRDC).
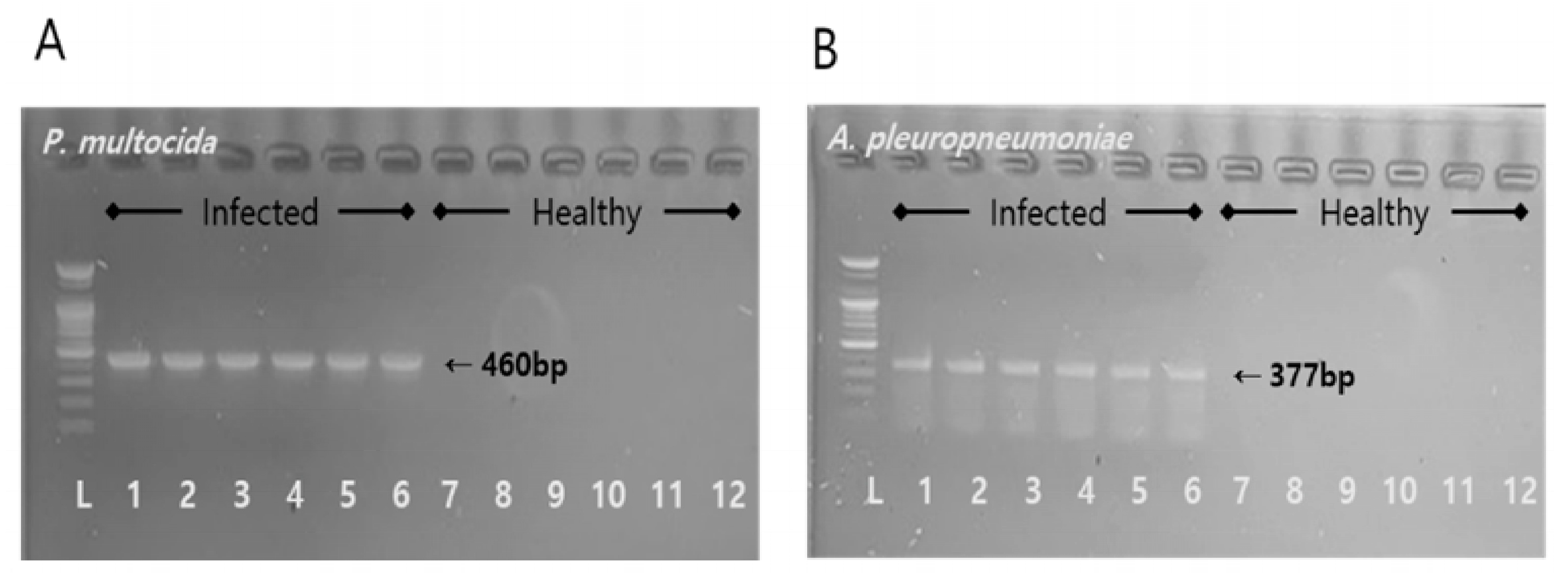
After co-infection of healthy piglets with AP and PM, clear clinical symptoms were observed. The infecting bacteria were identified via PCR, confirming the successful establishment of the co-infection model. A total of 12 h after the infection, the appearance of significant clinical signs in the piglets indicated successful establishment of the infection model. However, the observed variations in how the individual piglets responded to the infection could be explained by differences in microbial loads [
27], genetic predisposition [
28], and environmental conditions [
29]. The extent and timing of disease manifestation are directly affected by the microbial load, while an individual’s genetic background can determine their vulnerability or resilience to infection. Additionally, environmental influences, such as levels of stress and cleanliness, are key in shaping the immune response. These elements together lead to the varied responses seen in infectious diseases, underscoring the necessity of adopting a comprehensive approach in both research and the management of diseases.
The variability observed in the disease progression among the piglets infected with the
A. pleuropneumoniae and
P. multocida pathogens can be attributed to several critical factors, each contributing to the diverse outcomes seen in infectious disease models. One of the most significant factors is individual variations in the immune response [
30]. Each piglet’s immune system reacts differently to infection, influenced by genetic predispositions, previous exposure, and their overall health status at the time of infection. These variations dictate the severity and speed of disease progression, with some animals able to mount more effective responses than others. Another pivotal factor is the differences in pathogen load [
31]. The initial inoculum dose and the pathogen’s ability to replicate within the host vary significantly among individuals, directly impacting the disease’s progression rate. Factors such as the site of infection, the virulence of the pathogen strain, and the effectiveness of the host’s initial immune response can all influence the replication rate, leading to variability in the clinical outcomes among the infected piglets.
After intramuscular injection of gentamicin at a dose of 10 mg/kg in the healthy and co-infected piglets, the components of gentamicin (C
1, C
1a, C
2) were analyzed via validated LC/MS. Linearity validation is a crucial step in analytical method validation, ensuring that an analytical procedure demonstrates proportionality between the test results and the concentration of the analyte within a given range [
32]. A well-defined linear relationship confirms the method’s ability to produce results that are directly proportional to the concentration of the analyte in the sample. High R
2 values are indicative of the method’s strong predictability and consistent performance over the specified range. The slope values further emphasize this linearity. Ideally, a perfect linear method would have a slope of 1 [
33]. The obtained slopes, lying close to this ideal value, suggested that this method offered almost proportional response across the considered concentration range. The intercept values, which ideally should be close to zero when there is no inherent bias in the method at a zero concentration, show minor deviations, implying minimal systemic errors in the method. Furthermore, the LOD and LOQ provide insights into the method’s sensitivity [
34]. Lower LOD values signify that even trace amounts of the analyte can be detected, while the LOQ values suggest that the lowest concentration level that can be quantitatively determined with an acceptable precision and accuracy. In
Table 1, the presented results highlight the robust linearity of the methods for the analyzed components. The high R
2 values, ranging from 0.998 to 1.000, for both the intra-assay and inter-assay settings strongly indicated a near-perfect linear relationship between the observed results and the expected analyte concentrations for components C
1, C
1a and C
2. The strong linearity, combined with the sensitivity metrics (LOD and LOQ), underscored the method’s reliability and suitability for its intended purpose, ensuring accurate and consistent results across the analytical range.
Table 2 elaborates the intra-assay and inter-assay variations of the LC/MS method for the three gentamicin components (C
1, C
1a, C
2) at various nominal concentrations. For the gentamicin component C
1, the intra-assay accuracy percentages remained fairly close to the nominal values, with variations only ranging from 92.41% to 116.68%. Such values suggest that the method’s reliability in a single experimental run is commendable. Similarly, the inter-assay accuracy for C
1, which assesses the reproducibility across different experimental runs, showed values ranging from 96.24% to 113.64%. This tight range further confirms the method’s robustness and adaptability across different experimental conditions. For the gentamicin components C
1a and C
2, a similar trend in accuracy was observed. The intra-assay accuracies for both components closely matched their nominal concentrations, suggesting good reliability within a single experimental session. Yet, it was noted that the precision for C
1a in the intra-assay had a wider range, especially compared to C
1. This could indicate a slightly more variable consistency across repeated measurements for C
1a within the same run. The inter-assay results for C
1a and C
2 further confirmed the method’s reproducibility, with accuracy percentages staying within a narrow range of the nominal values. The precision values for these components in the inter-assay were also within acceptable limits, highlighting consistent results across different experimental conditions. The results from both the intra-assay and inter-assay measurements reflect a high degree of accuracy for the method, indicating its reliability in reflecting the true values. While the precision showed some variability, especially within the same experimental run, the ranges observed were within the acceptable limits for many applications. This reinforces the confidence in using this method for measuring gentamicin components in various experimental settings.
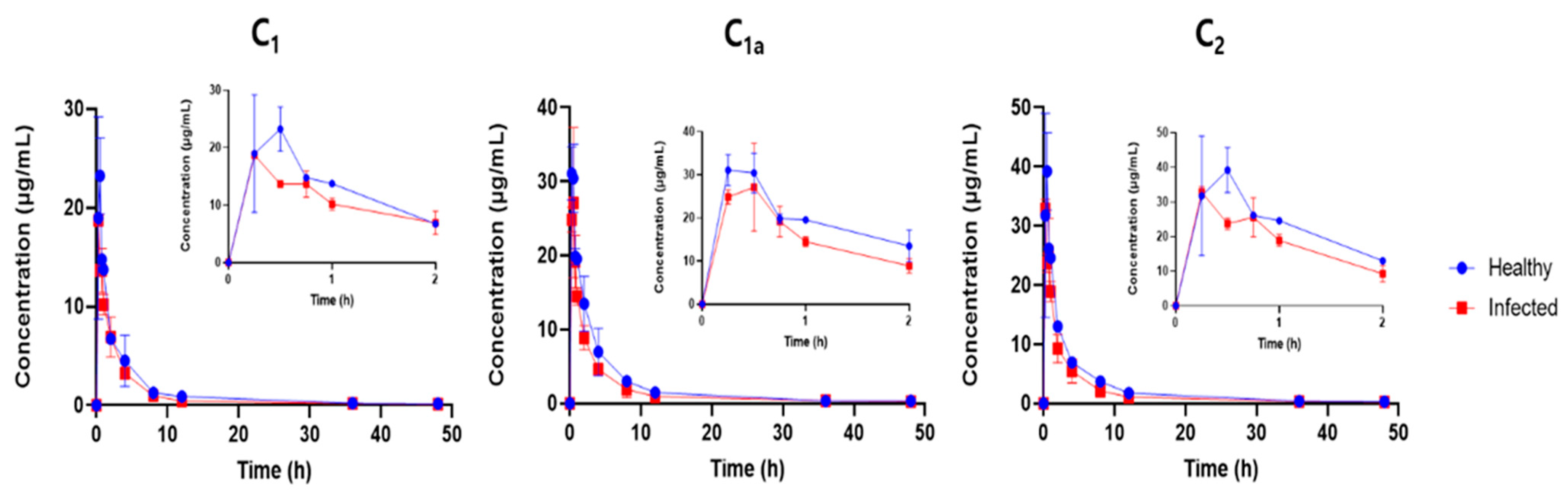
Gentamicin PK in piglets was characterized by rapid absorption from the IM site and a long elimination half-life. We found that it was inappropriate to make extensive comparisons of the PK findings of this study with previous reports because of scarcity of PK studies in piglets and several additional layers that could confound our comparisons such as the different routes of administration (intravenous versus IM), bioanalytical methods (immunoassay versus LC/MS), dose, age of animals and species of animals used. For example, in a 1995 study, gentamicin was administered intravenously (IV) to newborn male piglets aged from 4 to 12 h at the time of administration and 42-day-old castrated male piglets [
14]. In that study, the mean terminal half-life, mean residence time (MRT), volume of distribution and clearance were 5.19 h, 6.62 h, 785 mL/kg, and 121 L/h/kg in the newborn piglets and 3.50 h, 2.82 h, 474 mL/kg and 166 L/h/kg in the older group, respectively, indicating age-dependent gentamicin PK differences even after the same (IV) route of administration. The mean IM terminal half-life (17–24 h), MRT (6.7–8.0 h) and apparent volume of distribution (1756–2928 mL/kg) in the healthy piglets (
Table 3) were all greater than the IV PK values reported in the study cited above, whereas the mean apparent clearance of 74–189 mL/kg in the healthy piglets in our study was comparable with the IV clearance in the above study. The terminal half-life of IM gentamicin in the piglets in our study was also much longer than those reported for other species, including the IV half-life of 1 h in beagle dogs [
1], IV half-life of 2.4–4.3 h in horses [
2] and IM half-life of 8.5 h in laying hens [
12]. Such species-dependent and route-of-administration-dependent variability in drug clearance, as we suggested earlier, need to be identified by conducting PK studies of gentamicin in the species of interest so that tailored treatment strategies that ensure efficacy while minimizing side effects or the development of antibacterial drug resistance can be devised.
The observed differences in the gentamicin elimination rates between the healthy and diseased piglets could be attributed to the physiological changes that occur during infection [
35]. Infections can significantly affect the body’s normal physiological processes, leading to alterations in blood flow, tissue perfusion, and permeability. These changes are pivotal in understanding the pharmacokinetics of medications in infected versus healthy states. For instance, an infection often triggers an inflammatory response, which can alter blood flow and the distribution of blood to various organs. This, in turn, affects how drugs are transported to sites of action or elimination. Enhanced blood flow to certain organs might accelerate drug clearance, while reduced perfusion in others could delay it. Moreover, infections can compromise the integrity of biological membranes, affecting tissue permeability [
36]. This could lead to an increased or decreased distribution of drugs across tissues, influencing their bioavailability and elimination rates. Such physiological alterations could explain why gentamicin shows faster elimination in infected piglets. The drug’s distribution and clearance are likely affected by the systemic changes induced by the infection, leading to a shorter half-life in these animals. Understanding these changes is crucial for tailoring antibiotic therapies in veterinary medicine, ensuring effective dosing that accounts for the altered pharmacokinetic profile in diseased animals.
In contrast to a sheep study [
15] where gentamicin’s gamma half-life extended significantly to 82.1 h after multiple IM doses, this study in piglets found that gentamicin was eliminated faster in the tinfected animals compared to the healthy ones. This was evidenced by significantly greater terminal half-lives (T
1/2) in the healthy piglets across all the gentamicin components (C
1, C
1a and C
2), with the most pronounced difference observed in the C
1a component (24.94 ± 7.32 h in the healthy piglets versus 11.94 ± 0.08 h in the infected piglets,
p < 0.05). This highlights how the infection status can markedly influence the gentamicin pharmacokinetics, in contrast to the sheep study’s focus on dosage regimen differences.
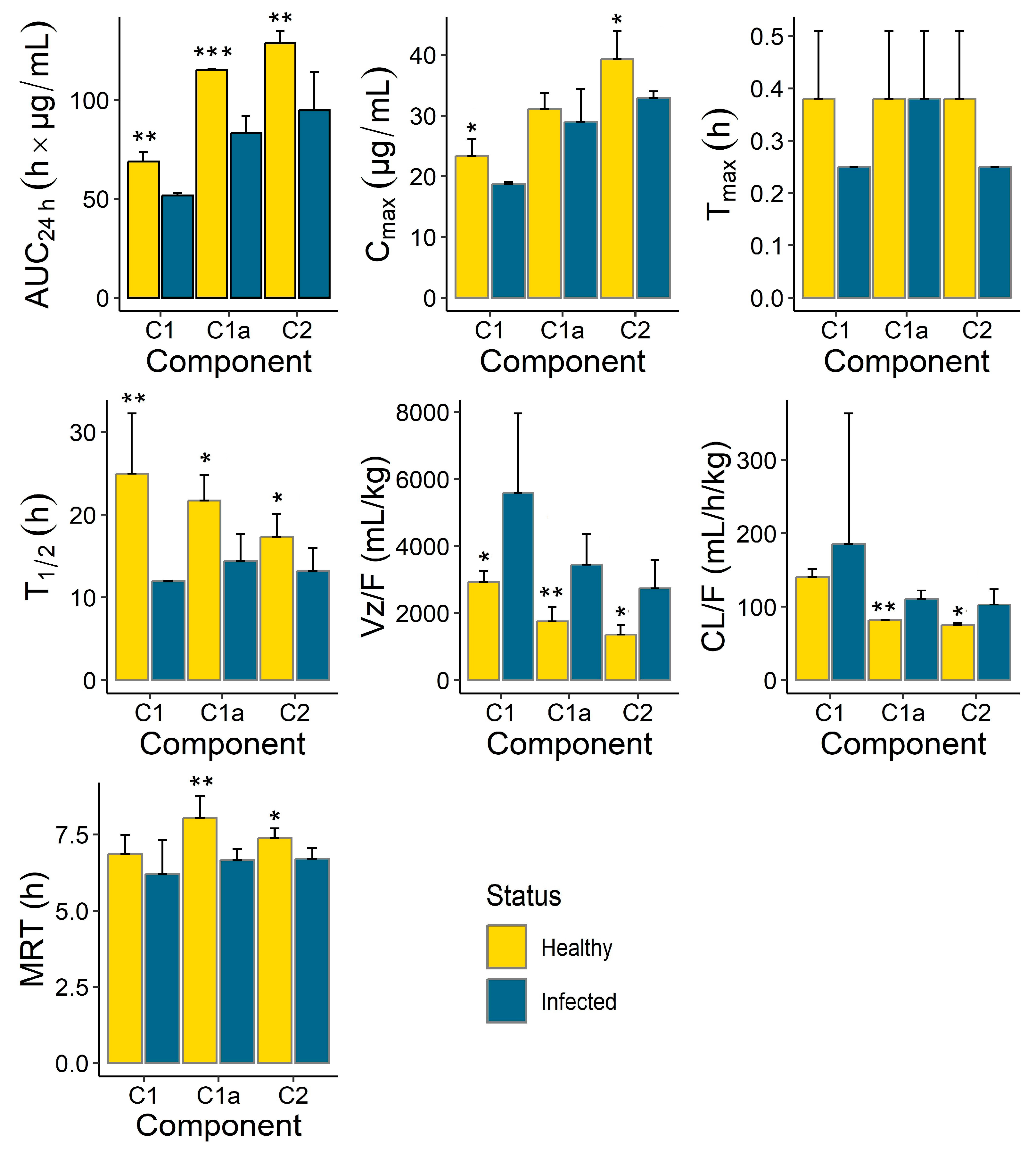
Differences were also observed in the PK parameters of the different gentamicin components (
Figure 4,
Table 3). While C
1a and C
2 seemed to clear at a similar rate, the apparent clearance of C
1 was different and was the highest. This held true for both the healthy and infected groups. Similarly, the apparent volumes of distribution of C
1a and C
2 were comparable and were smaller than that of C
1. These differences were more apparent in the healthy group but were overshadowed by the large within-group variability for C
1 observed in the infected animals for both CL/F and Vz/F (
Figure 3). No apparent differences were observed among the C
1, C
1a and C
2 components for the other PK parameters, including T
max, T
1/2 and MRT.
Our comparative PK analysis indicated that the rate and extent of gentamicin exposure were greater (greater C
max and AUC
24h) in the healthy animals compared to the infected piglets. This was also reflected in the slower elimination (longer terminal half-life and smaller apparent clearance) observed in the healthy animals compared to the infected piglets. Combining the individual exposures of the three components (C
1, C
1a, C
2), the total gentamicin C
max was 16.3% lower and the total gentamicin AUC
24h was 36.1% lower in the infected animals compared to the healthy piglets. These findings confirmed our initial hypothesis that the PK of gentamicin could be affected by the health status of the animals. Because our earlier comparative studies with tylosin, a macrolide antibiotic, also demonstrated significantly lower exposure (C
max and AUC
24h) and shorter T
1/2 values in co-infected pigs compared to healthy pigs [
37], we assume this phenomenon to be common to several classes of antimicrobial agents. The clinical significance of these differences is, however, dependent on several factors. These may include the magnitude of the PK differences with respect to the susceptibility or MIC of the target pathogen, whether the antibacterial drug has time-dependent or concentration-dependent activity, and the type of PK parameter affected by the health status of the animals. For instance, in our study, gentamicin absorption after administration via the IM route was rapid, and T
max was not affected by the health status of the piglets. And, just like other aminoglycosides, maintaining an early gentamicin C
max of 10 times or higher than the MIC could be effective against susceptible pathogens like
P. multocida with a MIC
90 of 4 μg/mL, and this was achievable for both the healthy and infected piglets. Aminoglycosides exhibit concentration-dependent killing along with prolonged persistent effects that protect against regrowth when the active drug concentration falls below the MIC [
21]. Therefore, the disease effects in later PK processes, such as a shorter elimination half-life in the infected piglets, would have minimal clinical implications. This is because the C
max was sufficiently higher than the target MIC, and the observed shorter half-life in the infected piglets was still long enough (greater than 11 h,
Table 3) to support the once-per-day IM regimen of gentamicin.
A notable limitation of our study is the absence of a direct investigation into the antibiotic residues resulting from the use of gentamicin in the piglets. Despite the comprehensive analysis of gentamicin pharmacokinetics and its disposition in healthy versus infected piglets, we did not extend our research to examine the persistence of gentamicin residues in the animal tissues post-treatment. Recognizing the critical importance of such studies is essential, as they provide key insights into the potential for antibiotic residues to enter the food chain, posing risks to consumer health and contributing to the broader challenge of antimicrobial resistance.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}