
Enhanced Efficacy of Ciprofloxacin and Tobramycin against Staphylococcus aureus When Combined with Corydalis Tuber and Berberine through Efflux Pump Inhibition


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Corydalis Tuber Extract and Berberine Synergistically Enhanced the Bactericidal Activity of Ciprofloxacin and Tobramycin
2.2. Time–Kill Curve Analyses Demonstrate Strong Synergistic Antibacterial Activity
2.3. Corydalis Tuber Extract and Berberine Are Efflux Pump Inhibitors
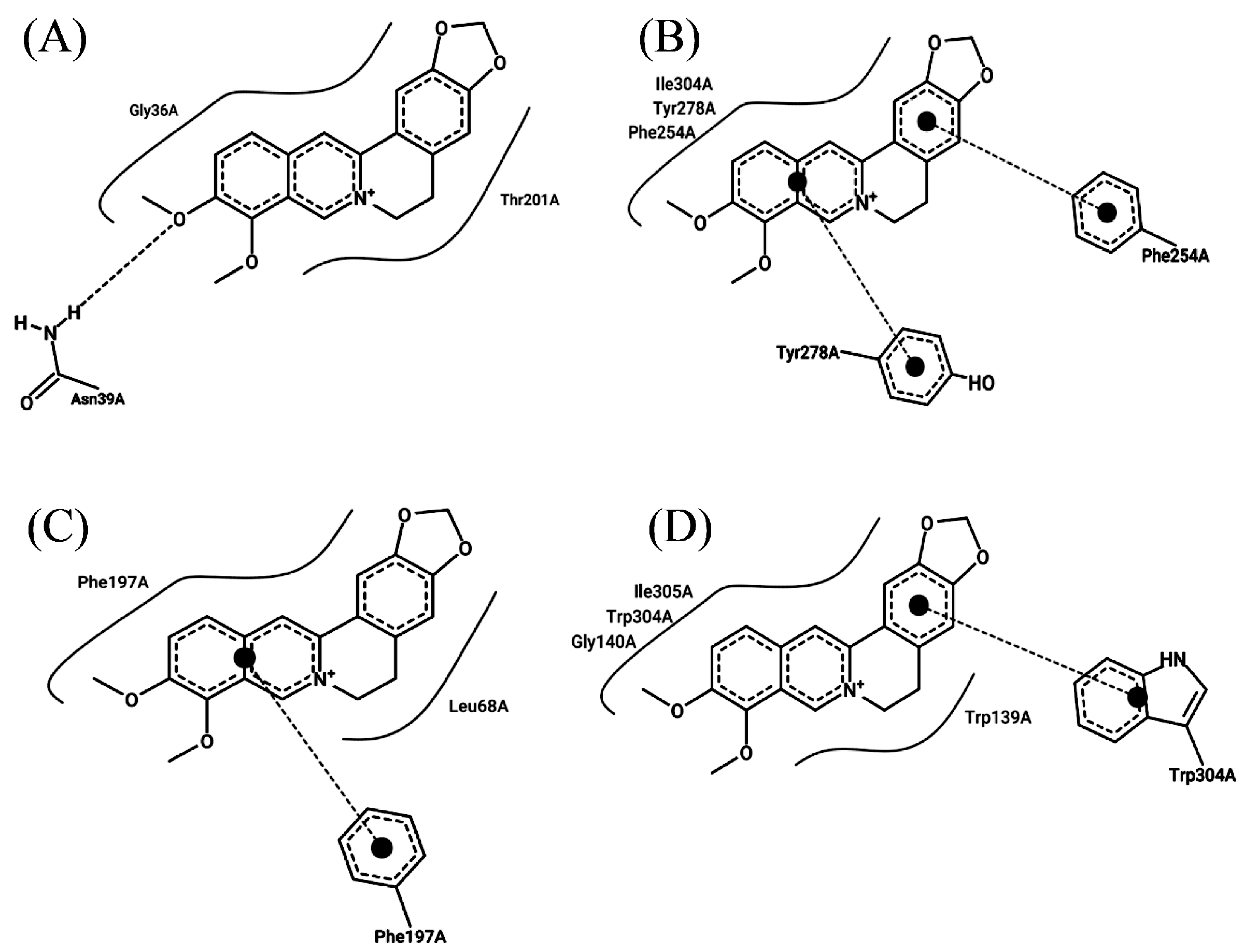
2.4. Berberine Interacts with Efflux Pump Proteins and the Protein PBP2a
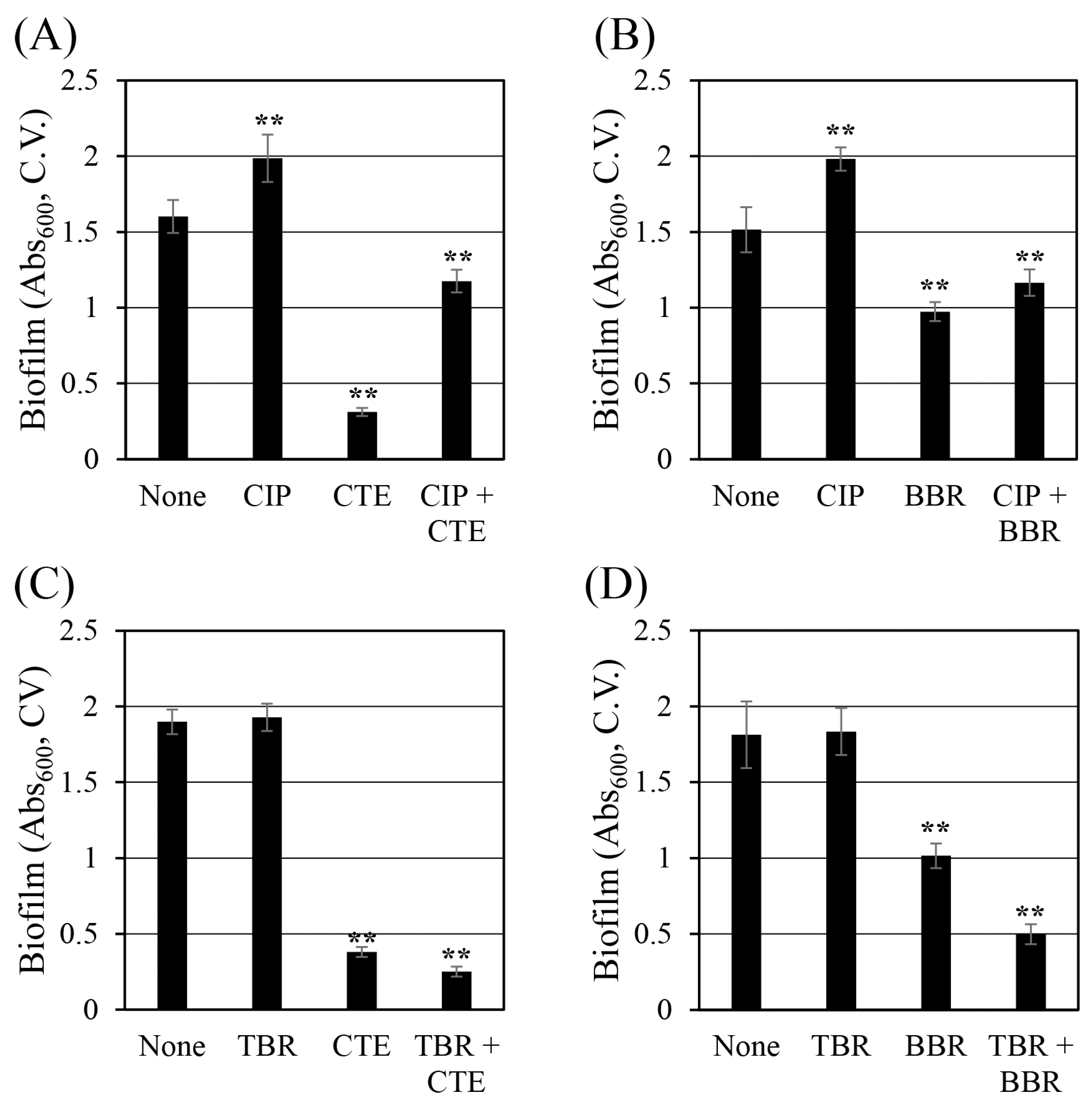
2.5. Corydalis Tuber Extract and Berberine Reduce Biofilm Formation
3. Materials and Methods
3.1. Staphylococcus aureus Strains and Culture Conditions
3.2. Chemicals
3.3. Preparing the Corydalis Tuber Extract
3.4. Analyzing the Berberine Concentration of the Corydalis Tuber Extract Using High-Performance Liquid Chromatography
3.5. Evaluating the Synergistic Antibacterial Activity of Antibiotics and Corydalis Tuber Extract or Berberine
3.6. Measuring the Time–Kill Curves for S. aureus ATCC 33593 Treated with Antibiotics and Corydalis Tuber Extract or Berberine
3.7. Evaluating the Efflux Pump Inhibition of Corydalis Tuber Extract and Berberine Using Ethidium Bromide
3.8. Analyzing the Interaction of Efflux Pump Proteins with Berberine Using Molecular Dynamic Simulation
3.9. Evaluating Biofilm Formation
3.10. Statistical Analysis
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tang, K.W.K.; Millar, B.C.; Moore, J.E. Antimicrobial Resistance (AMR). Br. J. Biomed. Sci. 2023, 80, 11387. [Google Scholar] [CrossRef] [PubMed]
- Durão, P.; Balbontín, R.; Gordo, I. Evolutionary mechanisms shaping the maintenance of antibiotic resistance. Trends Microbiol. 2018, 26, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug efflux pumps in Staphylococcus aureus: An Update. Open Microbiol. J. 2013, 7, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Pantosti, A.; Sanchini, A.; Monaco, M. Mechanisms of antibiotic resistance in Staphylococcus aureus. Future Microbiol. 2007, 2, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Köhler, T.; Pechère, J.C.; Plésiat, P. Bacterial antibiotic efflux systems of medical importance. Cell. Mol. Life Sci. 1999, 56, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Handzlik, J.; Matys, A.; Kieć-Kononowicz, K. Recent advances in multi-drug resistance (MDR) efflux pump inhibitors of Gram-positive bacteria S. aureus. Antibiotics 2013, 2, 28–45. [Google Scholar] [CrossRef] [PubMed]
- Kosmidis, C.; Schindler, B.D.; Jacinto, P.L.; Patel, D.; Bains, K.; Seo, S.M.; Kaatz, G.W. Expression of multidrug resistance efflux pump genes in clinical and environmental isolates of Staphylococcus aureus. Int. J. Antimicrob. Agents 2012, 40, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Diekema, D.J.; Hsueh, P.-R.; Mendes, R.E.; Pfaller, M.A.; Rolston, K.V.; Sader, H.S.; Jones, R.N. The microbiology of bloodstream infection: 20-year trends from the SENTRY antimicrobial surveillance program. Antimicrob. Agents Chemother. 2019, 63, e00355-19. [Google Scholar] [CrossRef]
- Lekshmi, M.; Ammini, P.; Adjei, J.; Sanford, L.M.; Shrestha, U.; Kumar, S.; Varela, M.F. Modulation of antimicrobial efflux pumps of the major facilitator superfamily in Staphylococcus aureus. AIMS Microbiol. 2018, 4, 1–18. [Google Scholar] [CrossRef]
- Sapula, S.A.; Brown, M.H. Antimicrobial drug efflux pumps in Staphylococcus aureus. In Efflux-Mediated Antimicrobial Resistance in Bacteria: Mechanisms, Regulation and Clinical Implications; Li, X.-Z., Elkins, C.A., Zgurskaya, H.I., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 165–195. [Google Scholar]
- Schindler, B.D.; Patel, D.; Seo, S.M.; Kaatz, G.W. Mutagenesis and modeling to predict structural and functional characteristics of the Staphylococcus aureus MepA multidrug efflux pump. J. Bacteriol. 2013, 195, 523–533. [Google Scholar] [CrossRef]
- Kim, M.; Seo, Y.; Kim, S.-G.; Choi, Y.; Kim, H.J.; Kim, T.-J. Synergistic antibiotic activity of Ricini Semen extract with oxacillin against methicillin-resistant Staphylococcus aureus. Antibiotics 2023, 12, 340. [Google Scholar] [CrossRef] [PubMed]
- Hemaiswarya, S.; Kruthiventi, A.K.; Doble, M. Synergism between natural products and antibiotics against infectious diseases. Phytomedicine 2008, 15, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Kim, T.-J. Synergistic antifungal activity of Magnoliae Cortex and Syzyii Flos against Candida albicans. J. Korean Wood Sci. Technol. 2021, 49, 142–153. [Google Scholar] [CrossRef]
- Yoon, J.; Kim, T.-J. Synergistic growth inhibition of herbal plant extract combinations against Candida albicans. J. Korean Wood Sci. Technol. 2023, 51, 145–156. [Google Scholar] [CrossRef]
- Zhao, W.-H.; Hu, Z.-Q.; Okubo, S.; Hara, Y.; Shimamura, T. Mechanism of synergy between epigallocatechin gallate and β-Lactams against methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2001, 45, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Mikulášová, M.; Chovanová, R.; Vaverková, Š. Synergism between antibiotics and plant extracts or essential oils with efflux pump inhibitory activity in coping with multidrug-resistant staphylococci. Phytochem. Rev. 2016, 15, 651–662. [Google Scholar] [CrossRef]
- Kalia, N.P.; Mahajan, P.; Mehra, R.; Nargotra, A.; Sharma, J.P.; Koul, S.; Khan, I.A. Capsaicin, a novel inhibitor of the NorA efflux pump, reduces the intracellular invasion of Staphylococcus aureus. J. Antimicrob. Chemother. 2012, 67, 2401–2408. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Barbosa, C.R.; Scherf, J.R.; de Freitas, T.S.; de Menezes, I.R.A.; Pereira, R.L.S.; dos Santos, J.F.S.; de Jesus, S.S.P.; Lopes, T.P.; de Sousa Silveira, Z.; de Morais Oliveira-Tintino, C.D.; et al. Effect of carvacrol and thymol on NorA efflux pump inhibition in multidrug-resistant (MDR) Staphylococcus aureus strains. J. Bioenerg. Biomembr. 2021, 53, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; He, S.; Wang, J.; Wang, C.; Wu, J.; Wang, W.; Li, F.; Li, S.; Zhao, C.; Li, F. A review of the traditional uses, botany, phytochemistry, pharmacology, pharmacokinetics, and toxicology of Corydalis yanhusuo. Nat. Prod. Commun. 2020, 15, 1934578X20957752. [Google Scholar]
- Lee, J.-R.; Park, S.-J.; Kim, Y.-W.; Cho, I.-J.; Byun, S.-H.; Kim, S.-C. Comparison of antimicrobial effects of Corydalis Tuber and processed Corydalis Tuber against propionibacterium acnes. J. Korean Med. Ophthalmol. Otolaryngol. Dermatol. 2011, 24, 17–26. [Google Scholar]
- Zhao, Y.; Gao, J.L.; Ji, J.W.; Gao, M.; Yin, Q.S.; Qiu, Q.L.; Wang, C.; Chen, S.Z.; Xu, J.; Liang, R.S.; et al. Cytotoxicity enhancement in MDA-MB-231 cells by the combination treatment of tetrahydropalmatine and berberine derived from Corydalis yanhusuo W. T. Wang. J. Intercult. Ethnopharmacol. 2014, 3, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Jin, L.; Li, L.; Wang, W.; Zeng, S.; Jiang, H.; Zhou, H. Development and validation of a HPLC-ESI-MS/MS method for simultaneous quantification of fourteen alkaloids in mouse plasma after oral administration of the extract of Corydalis yanhusuo tuber: Application to pharmacokinetic study. Molecules 2018, 23, 714. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Sun, J.; Li, W.; Zhang, S.; Yang, L.; Teng, Y.; Lv, K.; Liu, Y.; Su, Y.; Zhang, J.; et al. Analgesic effect of the main components of Corydalis yanhusuo (yanhusuo in Chinese) is caused by inhibition of voltage gated sodium channels. J. Ethnopharmacol. 2021, 280, 114457. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.-J.; Xing, X.-Y.; Xiao, X.-H.; Zhao, Y.-L.; Wei, J.-H.; Wang, J.-B.; Yang, R.-C.; Yang, M.-H. Effect of berberine on Escherichia coli, Bacillus subtilis, and their mixtures as determined by isothermal microcalorimetry. Appl. Microbiol. Biotechnol. 2012, 96, 503–510. [Google Scholar] [CrossRef]
- Xia, S.; Ma, L.; Wang, G.; Yang, J.; Zhang, M.; Wang, X.; Su, J.; Xie, M. In vitro antimicrobial activity and the mechanism of berberine against methicillin-resistant Staphylococcus aureus isolated from bloodstream infection patients. Infect. Drug Resist. 2022, 15, 1933–1944. [Google Scholar] [CrossRef]
- Domadia, P.N.; Bhunia, A.; Sivaraman, J.; Swarup, S.; Dasgupta, D. Berberine targets assembly of Escherichia coli cell division protein FtsZ. Biochemistry 2008, 47, 3225–3234. [Google Scholar] [CrossRef]
- Zhou, M.; Deng, Y.; Liu, M.; Liao, L.; Dai, X.; Guo, C.; Zhao, X.; He, L.; Peng, C.; Li, Y. The pharmacological activity of berberine, a review for liver protection. Eur. J. Pharmacol. 2021, 890, 173655. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, G.; Saigal, S.; Elongavan, A. Action and resistance mechanisms of antibiotics: A guide for clinicians. J. Anaesthesiol. Clin. Pharmacol. 2017, 33, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Jaktaji, R.P.; Mohammadi, P. Effect of total alkaloid extract of local Sophora alopecuroides on minimum inhibitory concentration and intracellular accumulation of ciprofloxacin, and acrA expression in highly resistant Escherichia coli clones. J. Glob. Antimicrob. Resist. 2018, 12, 55–60. [Google Scholar] [CrossRef]
- Schmitz, F.J.; Fluit, A.C.; Lückefahr, M.; Engler, B.; Hofmann, B.; Verhoef, J.; Heinz, H.P.; Hadding, U.; Jones, M.E. The effect of reserpine, an inhibitor of multidrug efflux pumps, on the in-vitro activities of ciprofloxacin, sparfloxacin and moxifloxacin against clinical isolates of Staphylococcus aureus. J. Antimicrob. Chemother. 1998, 42, 807–810. [Google Scholar] [CrossRef]
- Huang, W.; Pan, Y.; Jiang, H.; Chen, Y.; Hu, L.; Zhang, H.; Yan, J. A comprehensive quality evaluation method of Corydalis yanhusuo by HPLC fingerprints, chemometrics, and correlation analysis. J. Sep. Sci. 2021, 44, 2054–2064. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, S.; Fan, G.; Zou, H. Screening of antinociceptive components in Corydalis yanhusuo W.T. Wang by comprehensive two-dimensional liquid chromatography/tandem mass spectrometry. Anal. Bioanal. Chem. 2010, 396, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.-L.; Shi, J.-M.; He, K.; Zhang, Q.-W.; Li, S.-P.; Lee, S.M.-Y.; Wang, Y.-T. Yanhusuo extract inhibits metastasis of breast cancer cells by modulating mitogen-activated protein kinase signaling pathways. Oncol. Rep. 2008, 20, 819–824. [Google Scholar] [PubMed]
- Gao, J.L.; Shi, J.M.; Lee, S.M.; Zhang, Q.W.; Wang, Y.T. Angiogenic pathway inhibition of Corydalis yanhusuo and berberine in human umbilical vein endothelial cells. Oncol. Res. 2009, 17, 519–526. [Google Scholar] [CrossRef]
- Costa, S.S.; Falcão, C.; Viveiros, M.; Machado, D.; Martins, M.; Melo-Cristino, J.; Amaral, L.; Couto, I. Exploring the contribution of efflux on the resistance to fluoroquinolones in clinical isolates of Staphylococcus aureus. BMC Microbiol. 2011, 11, 241. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.Y.; Ma, D.; Nikaido, H. AcrD of Escherichia coli is an aminoglycoside efflux pump. J. Bacteriol. 2000, 182, 1754–1756. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.S.; Junqueira, E.; Palma, C.; Viveiros, M.; Melo-Cristino, J.; Amaral, L.; Couto, I. Resistance to antimicrobials mediated by efflux pumps in Staphylococcus aureus. Antibiotics 2013, 2, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Viveiros, M.; Martins, M.; Couto, I.; Rodrigues, L.; Spengler, G.; Martins, A.; Kristiansen, J.E.; Molnar, J.; Amaral, L. New methods for the identification of efflux mediated MDR bacteria, genetic assessment of regulators and efflux pump constituents, characterization of efflux systems and screening for inhibitors of efflux pumps. Curr. Drug Targets 2008, 9, 760–778. [Google Scholar] [CrossRef] [PubMed]
- Anoushiravani, M.; Falsafi, T.; Niknam, V. Proton motive force-dependent efflux of tetracycline in clinical isolates of Helicobacter pylori. J. Med. Microbiol. 2009, 58, 1309–1313. [Google Scholar] [CrossRef]
- Li, Y.; Ge, X. Role of berberine as a potential efflux pump inhibitor against MdfA from Escherichia coli: In vitro and in silico studies. Microbiol. Spectr. 2023, 11, e03324-22. [Google Scholar] [CrossRef]
- Aghayan, S.S.; Kalalian Mogadam, H.; Fazli, M.; Darban-Sarokhalil, D.; Khoramrooz, S.S.; Jabalameli, F.; Yaslianifard, S.; Mirzaii, M. The effects of berberine and palmatine on efflux pumps inhibition with different gene patterns in Pseudomonas aeruginosa isolated from burn infections. Avicenna J. Med. Biotechnol. 2017, 9, 2–7. [Google Scholar] [PubMed]
- Huang, H.; Wan, P.; Luo, X.; Lu, Y.; Li, X.; Xiong, W.; Zeng, Z. Tigecycline resistance-associated mutations in the MepA efflux pump in Staphylococcus aureus. Microbiol. Spectr. 2023, 11, e00634-23. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Song, Y.; Wang, L.; Kang, G.; Wang, P.; Yin, H.; Huang, H. A potential combination therapy of berberine hydrochloride with antibiotics against multidrug-resistant Acinetobacter baumannii. Front. Cell. Infect. Microbiol. 2021, 11, 660431. [Google Scholar] [CrossRef] [PubMed]
- Laudadio, E.; Cedraro, N.; Mangiaterra, G.; Citterio, B.; Mobbili, G.; Minnelli, C.; Bizzaro, D.; Biavasco, F.; Galeazzi, R. Natural alkaloid berberine activity against Pseudomonas aeruginosa MexXY-Mediated aminoglycoside resistance: In silico and in vitro studies. J. Nat. Prod. 2019, 82, 1935–1944. [Google Scholar] [CrossRef] [PubMed]
- Basri, D.F.; Zin, N.; Bakar, N.; Rahmat, F.; Mohtar, M. Synergistic effects of phytochemicals and oxacillin on laboratory passage-derived vancomycin-intermediate Staphylococcus aureus strain. J. Med. Sci. 2008, 8, 131–136. [Google Scholar]
- Dorcheh, F.A.; Balmeh, N.; Sanjari, S. In-silico investigation of antibacterial herbal compounds in order to find new antibiotic against Staphylococcus aureus and its resistant subtypes. Inform. Med. Unlocked 2022, 28, 100843. [Google Scholar] [CrossRef]
- Bouley, R.; Kumarasiri, M.; Peng, Z.; Otero, L.H.; Song, W.; Suckow, M.A.; Schroeder, V.A.; Wolter, W.R.; Lastochkin, E.; Antunes, N.T.; et al. Discovery of antibiotic (E)-3-(3-carboxyphenyl)-2-(4-cyanostyryl)quinazolin-4(3H)-one. J. Am. Chem. Soc. 2015, 137, 1738–1741. [Google Scholar] [CrossRef]
- Park, H.-S.; Ham, Y.; Shin, K.; Kim, Y.-S.; Kim, T.-J. Sanitizing effect of ethanol against biofilms formed by three Gram-negative pathogenic bacteria. Curr. Microbiol. 2015, 71, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.; Chakraborty, P.; Sarker, R.K.; Chatterjee, A.; Maiti, D.; Das, A.; Mandal, S.; Bhattacharjee, S.; Dastidar, D.G.; Tribedi, P. Tryptophan interferes with the quorum sensing and cell surface hydrophobicity of Staphylococcus aureus: A promising approach to inhibit the biofilm development. 3 Biotech 2021, 11, 376. [Google Scholar] [CrossRef]
- Luo, Z.-x.; Li, Y.; Liu, M.-f.; Zhao, R. Ciprofloxacin enhances the biofilm formation of Staphylococcus aureus via an agrC-dependent mechanism. Front. Microbiol. 2023, 14, 1328947. [Google Scholar] [CrossRef]
- Chu, M.; Zhang, M.-B.; Liu, Y.-C.; Kang, J.-R.; Chu, Z.-Y.; Yin, K.-L.; Ding, L.-Y.; Ding, R.; Xiao, R.-X.; Yin, Y.-N.; et al. Role of berberine in the treatment of methicillin-resistant Staphylococcus aureus infections. Sci. Rep. 2016, 6, 24748. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Zhao, X.; Li, W.; Shi, C.; Meng, R.; Liu, Z.; Yu, L. The synergy of berberine chloride and totarol against Staphylococcus aureus grown in planktonic and biofilm cultures. J. Med. Microbiol. 2015, 64, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Na, H.; Kim, T.-J. Synergistic antifungal activity of Phellodendri Cortex and Magnoliae Cortex against Candida albicans. J. Korean Wood Sci. Technol. 2022, 50, 12–30. [Google Scholar] [CrossRef]
- Soudeiha, M.A.H.; Dahdouh, E.A.; Azar, E.; Sarkis, D.K.; Daoud, Z. In vitro evaluation of the colistin-carbapenem combination in clinical isolates of A. baumannii using the checkerboard, Etest, and time-kill curve techniques. Front. Cell. Infect. Microbiol. 2017, 7, 209. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Wang, L.; Liu, S.; Hu, W.; Wang, J.; Deng, X.; Gao, J. Synergistic effect of lithocholic acid with gentamicin against Gram-positive bacteria but bot against Gram-negative bacteria. Molecules 2022, 27, 2318. [Google Scholar] [CrossRef] [PubMed]
- Longhi, C.; Marazzato, M.; Conte, M.P.; Iebba, V.; Schippa, S.; Seganti, L.; Comanducci, A. Effect of lactoferricin on fluoroquinolone susceptibility of uropathogenic Escherichia coli. J. Antibiot. 2009, 62, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Vorland, L.H.; Osbakk, S.A.; PerstØlen, T.; Ulvatne, H.; Rekdal, Ø.; Svendsen, J.S.; Gutteberg, T.J. Interference of the antimicrobial peptide lactoferricin B with the action of various antibiotics against Escherichia coli and Staphylococcus aureus. Scand. J. Infect. Dis. 1999, 31, 173–177. [Google Scholar] [PubMed]
- Zmantar, T.; Miladi, H.; Kouidhi, B.; Chaabouni, Y.; Ben Slama, R.; Bakhrouf, A.; Mahdouani, K.; Chaieb, K. Use of juglone as antibacterial and potential efflux pump inhibitors in Staphylococcus aureus isolated from the oral cavity. Microb. Pathog. 2016, 101, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Cramer, P. AlphaFold2 and the future of structural biology. Nat. Struct. Mol. Biol. 2021, 28, 704–705. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Schöning-Stierand, K.; Diedrich, K.; Fährrolfes, R.; Flachsenberg, F.; Meyder, A.; Nittinger, E.; Steinegger, R.; Rarey, M. ProteinsPlus: Interactive analysis of protein–ligand binding interfaces. Nucleic Acids Res. 2020, 48, W48–W53. [Google Scholar] [CrossRef] [PubMed]
- Ham, Y.; Park, H.-S.; Kim, M.; Kim, T.-J. Effect of sodium hypochlorite on the biofilms of Aeromonas hydrophila, Streptococcus mutans, and Yersinia enterocolitica. Microbiol. Biotechnol. Lett 2023, 51, 32–36. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Kim, M.; Kim, T.-J. Regulation of σB-dependent biofilm formation in Staphylococcus aureus through strain-specific signaling induced by diosgenin. Microorganisms 2023, 11, 2376. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | MIC (mg/L) | Test Concentration for Synergy (mg/L) | FIC | FIC Index | Decision |
|---|---|---|---|---|---|
| Ciprofloxacin | 2 | 0.25 | 0.125 | 0.25 | Synergistic |
| Corydalis Tuber | 512 | 64 | 0.125 | ||
| Ciprofloxacin | 2 | 0.25 | 0.125 | 0.375 | Synergistic |
| Berberine | 256 | 64 | 0.25 | ||
| Oxacillin | 128 | 8 | 0.0625 | 0.5625 | Partial synergistic |
| Corydalis Tuber | 512 | 256 | 0.5 | ||
| Oxacillin | 128 | 8 | 0.0625 | 0.5625 | Partial synergistic |
| Berberine | 256 | 128 | 0.5 | ||
| Tobramycin | 1024 | 128 | 0.125 | 0.375 | Synergistic |
| Corydalis Tuber | 512 | 128 | 0.25 | ||
| Tobramycin | 1024 | 256 | 0.25 | 0.5 | Synergistic |
| Berberine | 256 | 64 | 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, Y.; Kim, M.; Kim, T.-J. Enhanced Efficacy of Ciprofloxacin and Tobramycin against Staphylococcus aureus When Combined with Corydalis Tuber and Berberine through Efflux Pump Inhibition. Antibiotics 2024, 13, 469. https://doi.org/10.3390/antibiotics13050469
Seo Y, Kim M, Kim T-J. Enhanced Efficacy of Ciprofloxacin and Tobramycin against Staphylococcus aureus When Combined with Corydalis Tuber and Berberine through Efflux Pump Inhibition. Antibiotics. 2024; 13(5):469. https://doi.org/10.3390/antibiotics13050469
Chicago/Turabian StyleSeo, Yena, Minjun Kim, and Tae-Jong Kim. 2024. "Enhanced Efficacy of Ciprofloxacin and Tobramycin against Staphylococcus aureus When Combined with Corydalis Tuber and Berberine through Efflux Pump Inhibition" Antibiotics 13, no. 5: 469. https://doi.org/10.3390/antibiotics13050469