
Unveiling the Antimicrobial, Anti-Biofilm, and Anti-Quorum-Sensing Potential of Paederia foetida Linn. Leaf Extract against Staphylococcus aureus: An Integrated In Vitro–In Silico Investigation
 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Qualitative Profiling and Identification of Bioactive Anti-Quorum-Sensing Compounds from the Ethanolic Crude Extract by LC-MS/MS
2.2. Antimicrobial Activity of Paederia Foetida Leaf Crude Extract against S. aureus
2.3. Impact of Crude Extract on S. aureus Morphology Assessed by SEM
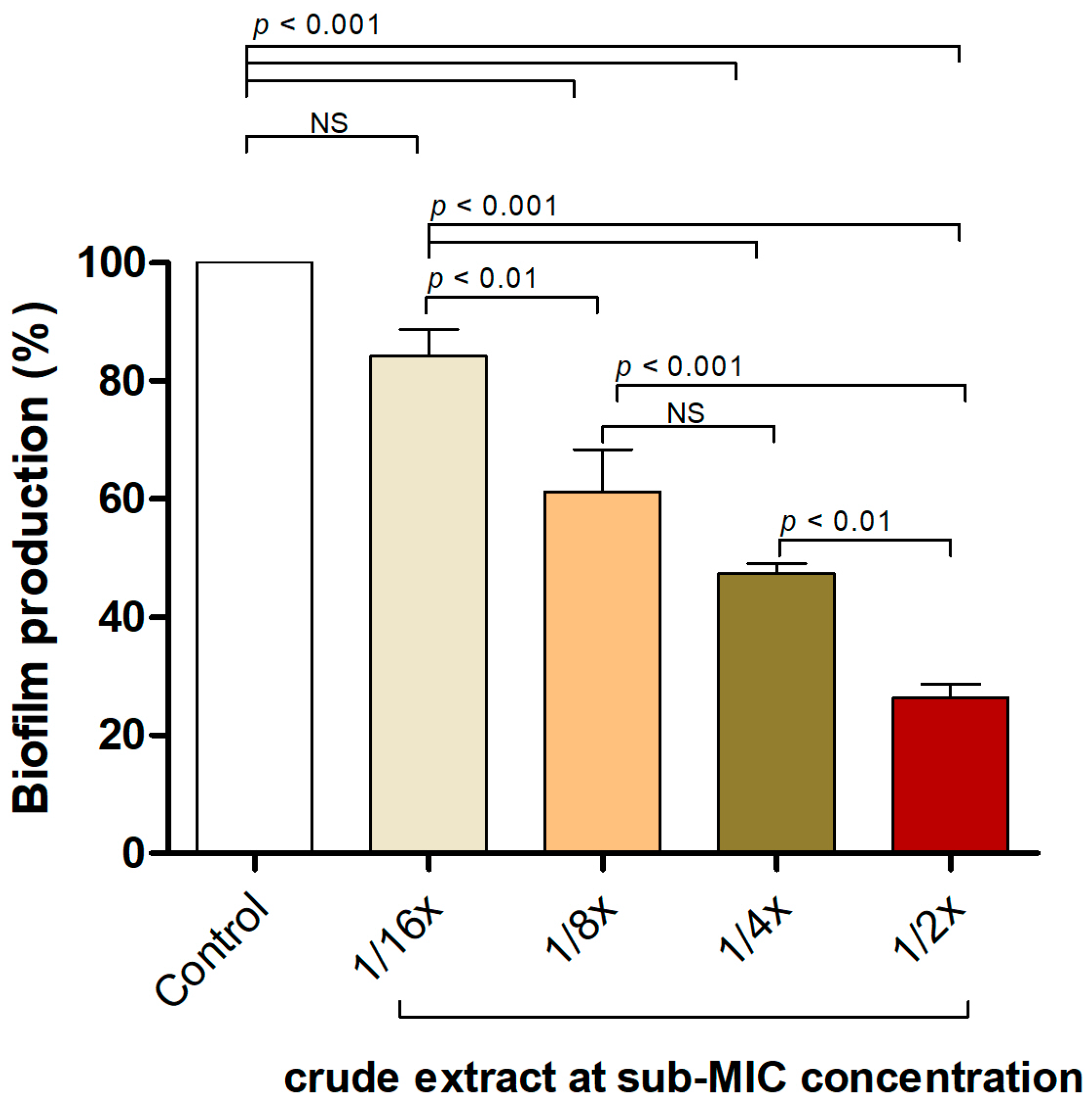
2.4. P. foetida Extract Interfered with S. aureus Biofilm Formation at Sub-Inhibitory Concentrations
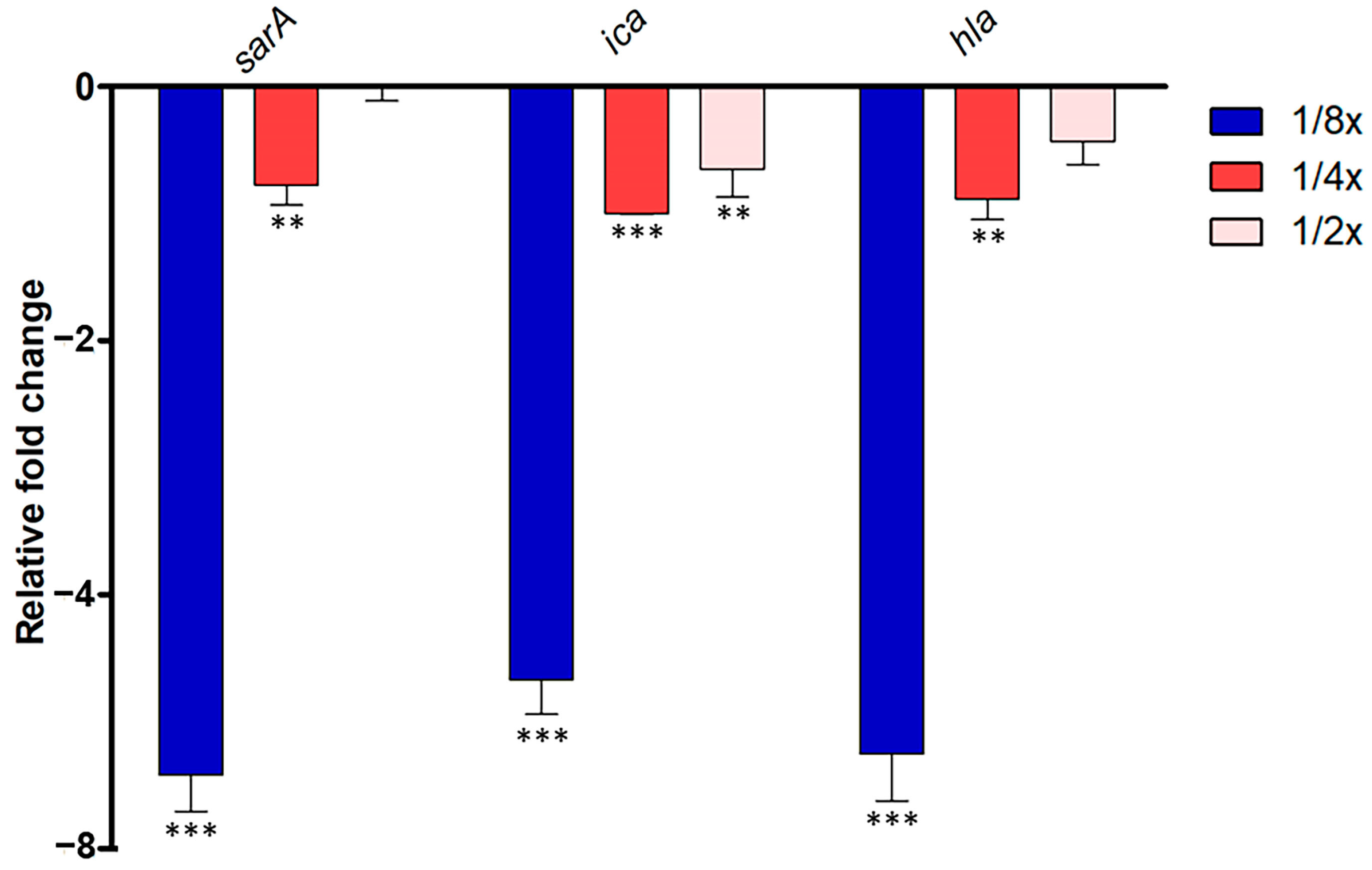
2.5. Interference of P. foetida Linn. in Quorum-Sensing Signaling of S. aurues
2.6. Molecular Docking of Candidate Anti-QS Ligand with S. aureus QS Transcriptional Regulators and In Silico ADMET Evaluation
3. Discussion
4. Materials and Methods
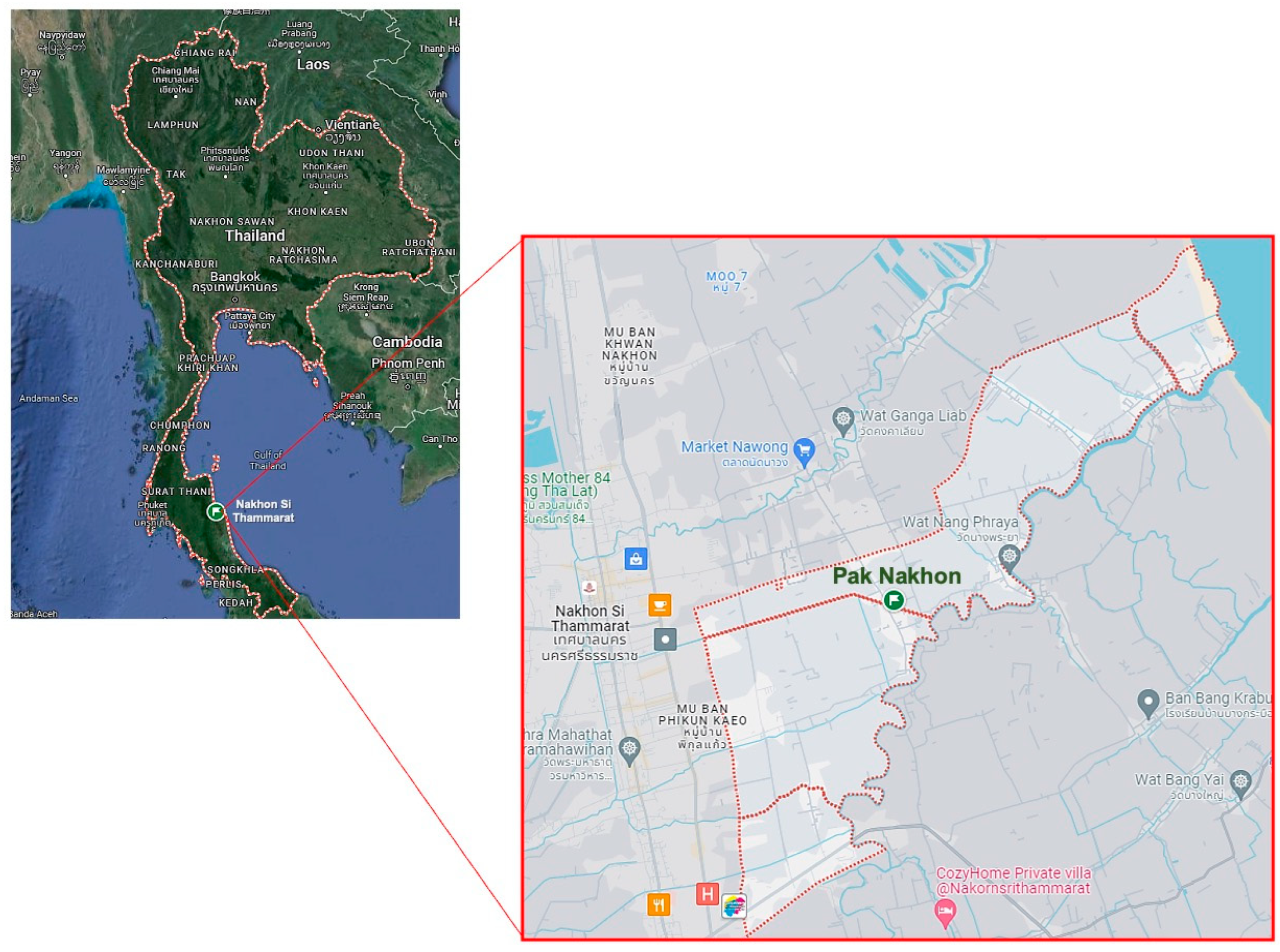
4.1. Location of Plant Collection and Bacterial Strains
4.2. Preparation of P. foetida Linn. Crude Extracts and LC-MS/MS Analysis
4.3. Minimal Inhibitory Concentration (MIC) and Minimal Bactericidal Concentration (MBC) of Ethanolic P. foetida Linn. Crude Extract against S. aureus
4.4. Scanning Electron Microscopy Analysis and Imaging
4.5. Biofilm Formation Inhibition Using Microtiter Plates Assay
4.6. QS Gene Expression Using Quantitative Reverse Transcription PCR (qRT-PCR)
4.7. In Silico Analysis of Anti-QS Activity of Candidate Bioactive Compounds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) | PCR Product (bp) | References |
|---|---|---|---|---|
| 16S rRNA | TGATCCTGGCTCAGGATGA | TTCGCTCGACTTGCATGTA | 57 | [45] |
| sarA | GCTGTATTGACATACATCAGCGAA | CGTTGTTTGCTTCAGTGATTCGT | 250 | [46] |
| hla | ACAATTTTAGAGAGCCCAACTGAT | TCCCCAATTTTGATTCACCAT | 77 | [45] |
| ica | TCGCACTCTTTATTGATAGTCGCTACGAG | TGCGACAAGAACTACTGCTGCGTTAAT | 98 | [46] |
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef]
- Bouhrour, N.; Nibbering, P.H.; Bendali, F. Medical device-associated biofilm infections and multidrug-resistant pathogens. Pathogens 2024, 13, 393. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Idrees, M.; Sawant, S.; Karodia, N.; Rahman, A. Staphylococcus aureus biofilm: Morphology, genetics, pathogenesis and treatment strategies. Int. J. Environ. Res. Public Health 2021, 18, 7602. [Google Scholar] [CrossRef]
- Ricciardi, B.F.; Muthukrishnan, G.; Masters, E.; Ninomiya, M.; Lee, C.C.; Schwarz, E.M. Staphylococcus aureus evasion of host immunity in prosthetic joint infection: Biofilm and beyond. Curr. Rev. Musculoskelet. Med. 2018, 11, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Bano, S.; Hassan, N.; Rafiq, M.; Hassan, F.; Rehman, M.; Iqbal, N.; Ahmed, S.; Kang, Y.Q. Biofilms as battlefield armor for bacteria against antibiotics: Challenges and combating strategies. Microorganisms 2023, 11, 2595. [Google Scholar] [CrossRef]
- Archer, N.K.; Mazaitis, M.J.; Costerton, J.W.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms: Properties, regulation, and roles in human disease. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef]
- Venkateswaran, P.; Vasudevan, S.; David, H.; Shaktivel, A.; Shanmugam, K.; Neelakantan, P.; Solomon, A.P. Revisiting ESKAPE pathogens: Virulence, resistance, and combating strategies focusing on quorum sensing. Front. Cell. Infect. Microbiol. 2023, 13, 1128214. [Google Scholar] [CrossRef]
- Chung, P.Y.; Toh, Y.S. Anti-biofilm agents: Recent breakthrough against multi-drug resistant Staphylococcus aureus. Pathog. Dis. 2014, 70, 231–239. [Google Scholar] [CrossRef]
- Yadav, M.K.; Chae, S.W.; Go, Y.Y.; Im, G.J.; Song, J.J. In vitro multi-species biofilms of methicillin-resistant Staphylococcus aureus and Pseudomonas aeruginosa and host interaction during in vivo colonization of an otitis media rat model. Front. Cell. Infect. Microbiol. 2017, 7, 125. [Google Scholar] [CrossRef]
- Paharik, A.E.; Horswill, A.R. The staphylococcal biofilm: Adhesins, regulation, and host response. In Virulence Mechanisms of Bacterial Pathogens, 5th ed.; Fodor, A.A., Adler, B., Eds.; ASM Press: Washington, DC, USA, 2016; pp. 529–566. [Google Scholar]
- Morshed, H.; Islam, M.S.; Parvin, S.; Ahmed, M.U.; Islam, M.S.; Mostofa, A.G.M.; Sayeed, M.S.B. Antimicrobial and cytotoxic activity of the methanol extract of Paederia foetida Linn. (Rubiaceae). J. Appl. Pharm. Sci. 2012, 2, 77–80. [Google Scholar]
- Karmakar, U.; Akter, S.; Sultana, S. Investigation of antioxidant, analgesic, antimicrobial, and anthelmintic activity of the aerial parts of Paederia foetida (Family: Rubiaceaea). Jordan J. Pharm. Sci. 2020, 13, 2. [Google Scholar]
- Soni, R.K.; Irchhaiya, R.; Dixit, V.; Alok, S. Paederia foetida Linn: Phytochemistry, pharmacological and traditional uses. Int. J. Pharm. Sci. Res. 2013, 4, 4525. [Google Scholar]
- Priyanto, J.A.; Prastya, M.E.; Sinarawadi, G.S.; Datu’salamah, W.; Avelina, T.Y.; Yanuar, A.I.A.; Budianto, I.T.; Mozef, T. The antibacterial and antibiofilm potential of Paederia foetida Linn. leaves extract. J. Appl. Pharm. Sci. 2022, 12, 117–124. [Google Scholar] [CrossRef]
- Dutta, P.P.; Marbaniang, K.; Sen, S.; Dey, B.K.; Talukdar, N.C. A review on phytochemistry of Paederia foetida Linn. Phytomedicine Plus 2023, 3, 100411. [Google Scholar] [CrossRef]
- Asfour, H.Z. Anti-quorum sensing natural compounds. J. Microsc. Ultrastruct. 2018, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; Cheng, Q.; Zhu, J.; Li, M. Developing natural products as potential anti-biofilm agents. Chin. Med. 2019, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Asma, S.T.; Imre, K.; Morar, A.; Herman, V.; Acaroz, U.; Mukhtar, H.; Kiran, A.; Ilk, S.; Rodrigo, C.P.; Boari, R.M.; et al. An overview of biofilm formation-combating strategies and mechanisms of action of antibiofilm agents. Life 2022, 12, 1110. [Google Scholar] [CrossRef]
- Kumar, S.; Balaya, R.D.A.; Kanekar, S.; Raju, R.; Prasad, T.S.K.; Kandasamy, R.K. Computational tools for exploring peptide-membrane interactions in gram-positive bacteria. Comput. Struct. Biotechnol. J. 2023, 21, 1995–2008. [Google Scholar] [CrossRef]
- Wang, X.; Ni, D.; Liu, Y.; Lu, S. Rational design of peptide-based inhibitors disrupting protein-protein interactions. Front. Chem. 2021, 9, 682675. [Google Scholar] [CrossRef] [PubMed]
- Fleitas Martínez, O.; Cardoso, M.H.; Ribeiro, S.M.; Franco, O.L. Recent advances in anti-virulence therapeutic strategies with a focus on dismantling bacterial membrane microdomains, toxin neutralization, quorum-sensing interference and biofilm inhibition. Front. Cell. Infect. Microbiol. 2019, 9, 74. [Google Scholar] [CrossRef] [PubMed]
- Cella, E.; Giovanetti, M.; Benedetti, F.; Scarpa, F.; Johnston, C.; Borsetti, A.; Galuppo, L.; Pompili, M.; D’Agostino, C.; Lanni, L.; et al. Joining forces against antibiotic resistance: The one health solution. Pathogens 2023, 12, 1074. [Google Scholar] [CrossRef] [PubMed]
- Namsena, P.; Maneetap, J.; Phonpheng, D.; Ngamsane, W. Comparative study of antibacterial activity and phytochemical analysis of stem, root and leaf extracts of Paederia foetida L. against phytopathogenic bacteria. Agric. Nat. Resour. 2019, 53, 395–401. [Google Scholar]
- Uddin, B.; Nahar, T.; Khalil, M.I.; Hossain, S. In vitro antibacterial activity of the ethanol extract of Paederia foetida L. (Rubiaceae) leaves. Bangladesh J. life Sci. 2007, 19, 141–143. [Google Scholar]
- Defres, S.; Marwick, C.; Nathwani, D. MRSA as a cause of lung infection including airway infection, community-acquired pneumonia and hospital-acquired pneumonia. Eur. Respir. J. 2009, 34, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.; Cirioni, O.; Giacometti, A.; Chiselli, R.; Braunstein, J.B.; Silvestri, C.; Mocchegiani, F.; Saba, V.; Scalise, G. Treatment of Staphylococcus aureus Biofilm Infection by the Quorum-Sensing Inhibitor RIP. Antimicrob. Agents. Chemother. 2007, 51, 2226–2229. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, E.; Kollef, M.H.; Nathwani, D. Pneumonia attributable to methicillin-resistant Staphylococcus aureus. Clin. Infect. Dis. 2008, 46, S378–S385. [Google Scholar] [CrossRef]
- Mostafa, A.A.; Al-Askar, A.A.; Almaary, K.S.; Dawoud, T.M.; Sholkamy, E.N.; Bakri, M.M. Evaluation of plant extract antimicrobial efficacy against bacterial strains associated with foodborne illnesses. Saudi J. Biol. Sci. 2018, 25, 361–366. [Google Scholar] [CrossRef]
- Cheung, A.L.; Nishina, K.A.; Trotonda, M.P.; Tamber, S. The SarA protein family of Staphylococcus aureus. Int. J. Biochem. Cell Biol. 2008, 40, 355–361. [Google Scholar] [CrossRef]
- Schlag, S.; Biswas, R.; Krismer, B.; Kohler, T.; Zoll, S.; Yu, W.; Gotz, F. The significance of staphylococcal wall teichoic acid in modulating the activity of the major autolysin Atl. Mol. Microbiol. 2010, 75, 864–873. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Septama, A.W.; Tasfiyati, A.N.; Kristiana, R.; Jaisi, A. Chemical profiles of essential oil from Javanese turmeric (Curcuma xanthorrhiza Roxb.), evaluation of its antibacterial and antibiofilm activities against selected clinical isolates. S. Afr. J. Bot. 2022, 146, 728–734. [Google Scholar] [CrossRef]
- Jaber, H.; Oubihi, A.; Ouryemchi, I.; Boulamtat, R.; Oubayoucef, A.; Bourkhiss, B.; Ouhssine, M. Chemical composition and antibacterial activities of eight plant essential oils from Morocco against Escherichia coli strains isolated from different turkey organs. Biochem. Res. Int. 2021, 2021, 5525956. [Google Scholar] [CrossRef]
- Herman, T.S.; da Silva Goersch, C.; Bocca, A.L.; Fernandes, L. Resazurin to determine the minimum inhibitory concentration on antifungal susceptibility assays for Fonsecaea sp. using a modified EUCAST protocol. Braz. J. Microbiol. 2024; in press. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Richardson, A.; Sofian-Azirun, M. Antibacterial activity of propolis and honey against Staphylococcus aureus and Escherichia coli. Afr. J. Microbiol. Res. 2010, 4, 1872–1878. [Google Scholar]
- Jiamboonsri, P.; Eurtivong, C.; Wanwong, S. Assessing the potential of gallic acid and methyl gallate to enhance the efficacy of β-lactam antibiotics against methicillin-resistant Staphylococcus aureus by targeting β-lactamase: In silico and in vitro studies. Antibiotics 2023, 12, 1622. [Google Scholar] [CrossRef]
- Gajewska, J.; Chajęcka-Wierzchowska, W. Biofilm formation ability and presence of adhesion genes among coagulase-negative and coagulase-positive staphylococci isolates from raw cow’s milk. Pathogens 2020, 9, 654. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Madhavi Sastry, G.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Halgren, T. New method for fast and accurate binding-site identification and analysis. Chem. Biol. Drug Des. 2007, 69, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Forli, S.; Huey, R.; Pique, M.E.; Sanner, M.F.; Goodsell, D.S.; Olson, A.J. Computational protein–ligand docking and virtual drug screening with the AutoDock suite. Nature Protocols 2016, 11, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Wu, Z.; Yi, J.; Fu, L.; Yang, Z.; Hsieh, C.; Tomczuk, M.; Sander, T.; Cao, D. ADMETlab 2.0: An integrated online platform for accurate and comprehensive predictions of ADMET properties. Nucleic Acids Res. 2021, 49, W5–W14. [Google Scholar] [CrossRef] [PubMed]
- Sully, E.K.; Malachowa, N.; Elmore, B.O.; Alexander, S.M.; Femling, J.K.; Gray, B.M.; DeLeo, F.R.; Otto, M.; Cheung, A.L.; Edwards, B.S.; et al. Selective chemical inhibition of agr quorum sensing in Staphylococcus aureus promotes host defense with minimal impact on resistance. PLoS Pathog. 2014, 10, e1004174. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, T.; Wang, K.E.; Hou, C.; Cai, S.; Huang, Y.; Du, Z.; Huang, H.; Kong, J.; Chen, Y. Baicalein inhibits Staphylococcus aureus biofilm formation and the quorum sensing system in vitro. PLoS ONE 2016, 11, e0153468. [Google Scholar] [CrossRef]


| No. | Name of Bioactive Compounds and PubChem CID | Structure of the Bioactive Compounds | Molecular Formula | XLogP | Hydrogen Bond Donor | Hydrogen Bond Acceptor | Retention Time (min) | Mass-to-Charge Ratio (m/z) |
|---|---|---|---|---|---|---|---|---|
| 1. | Isoquercetin (CID 5280804) |  | C21H20O12 | 0.4 | 8 | 12 | 9.22 | 303.0497 |
| 2. | Quercetin 3-galactoside (CID 5281643) | 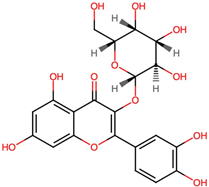 | C21H20O12 | 0.4 | 8 | 12 | 9.22 | 303.0493 |
| 3. | Quercetin 3-beta-D-glucoside (CID 44259136) | 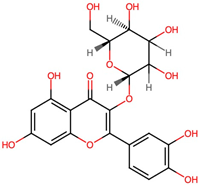 | C21H20O12 | 0.4 | 8 | 12 | 9.22 | 303.0503 |
| 4. | Kaempferol 3-alpha-D-galactoside (CID 44258736) |  | C21H20O11 | 0.7 | 7 | 11 | 9.60 | 287.0536 |
| 5. | Kaempferol 3-alpha-D-glucoside (CID 44258798) | 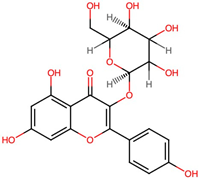 | C21H20O11 | 0.7 | 7 | 11 | 9.63 | 287.0532 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santajit, S.; Tunyong, W.; Horpet, D.; Binmut, A.; Kong-Ngoen, T.; Wisessaowapak, C.; Thavorasak, T.; Pumirat, P.; Indrawattana, N. Unveiling the Antimicrobial, Anti-Biofilm, and Anti-Quorum-Sensing Potential of Paederia foetida Linn. Leaf Extract against Staphylococcus aureus: An Integrated In Vitro–In Silico Investigation. Antibiotics 2024, 13, 613. https://doi.org/10.3390/antibiotics13070613
Santajit S, Tunyong W, Horpet D, Binmut A, Kong-Ngoen T, Wisessaowapak C, Thavorasak T, Pumirat P, Indrawattana N. Unveiling the Antimicrobial, Anti-Biofilm, and Anti-Quorum-Sensing Potential of Paederia foetida Linn. Leaf Extract against Staphylococcus aureus: An Integrated In Vitro–In Silico Investigation. Antibiotics. 2024; 13(7):613. https://doi.org/10.3390/antibiotics13070613
Chicago/Turabian StyleSantajit, Sirijan, Witawat Tunyong, Dararat Horpet, Asma Binmut, Thida Kong-Ngoen, Churaibhon Wisessaowapak, Techit Thavorasak, Pornpan Pumirat, and Nitaya Indrawattana. 2024. "Unveiling the Antimicrobial, Anti-Biofilm, and Anti-Quorum-Sensing Potential of Paederia foetida Linn. Leaf Extract against Staphylococcus aureus: An Integrated In Vitro–In Silico Investigation" Antibiotics 13, no. 7: 613. https://doi.org/10.3390/antibiotics13070613