
First Data on WGS-Based Typing and Antimicrobial Resistance of Human Salmonella Enteritidis Isolates in Greece


Abstract
:1. Introduction
2. Results
2.1. Plasmid Detection
2.2. Antimicrobial Resistance
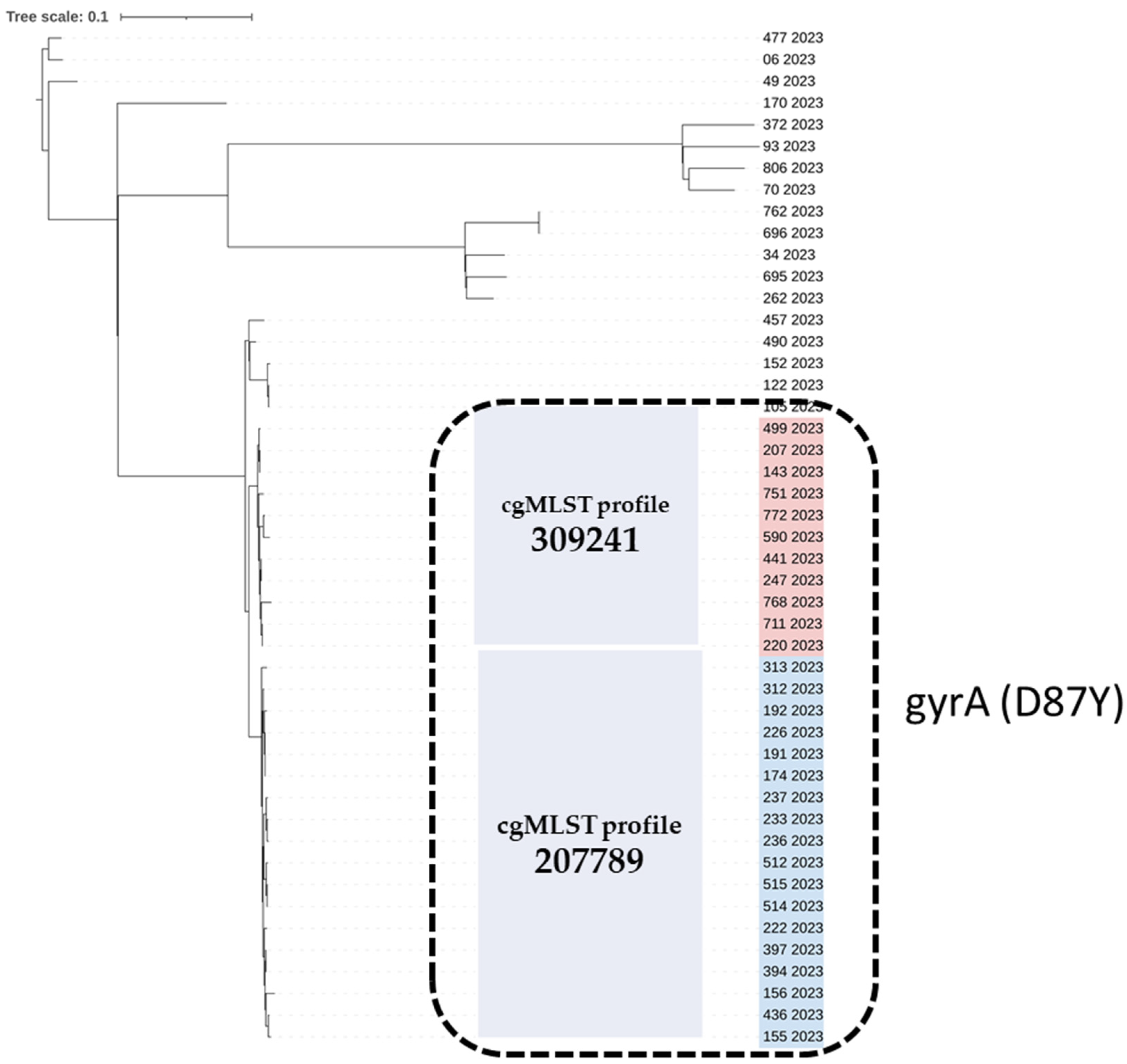
2.3. Genetic Diversity-cgMLST and SNP Analysis
2.4. Salmonella Pathogenicity Island (SPI) Identification
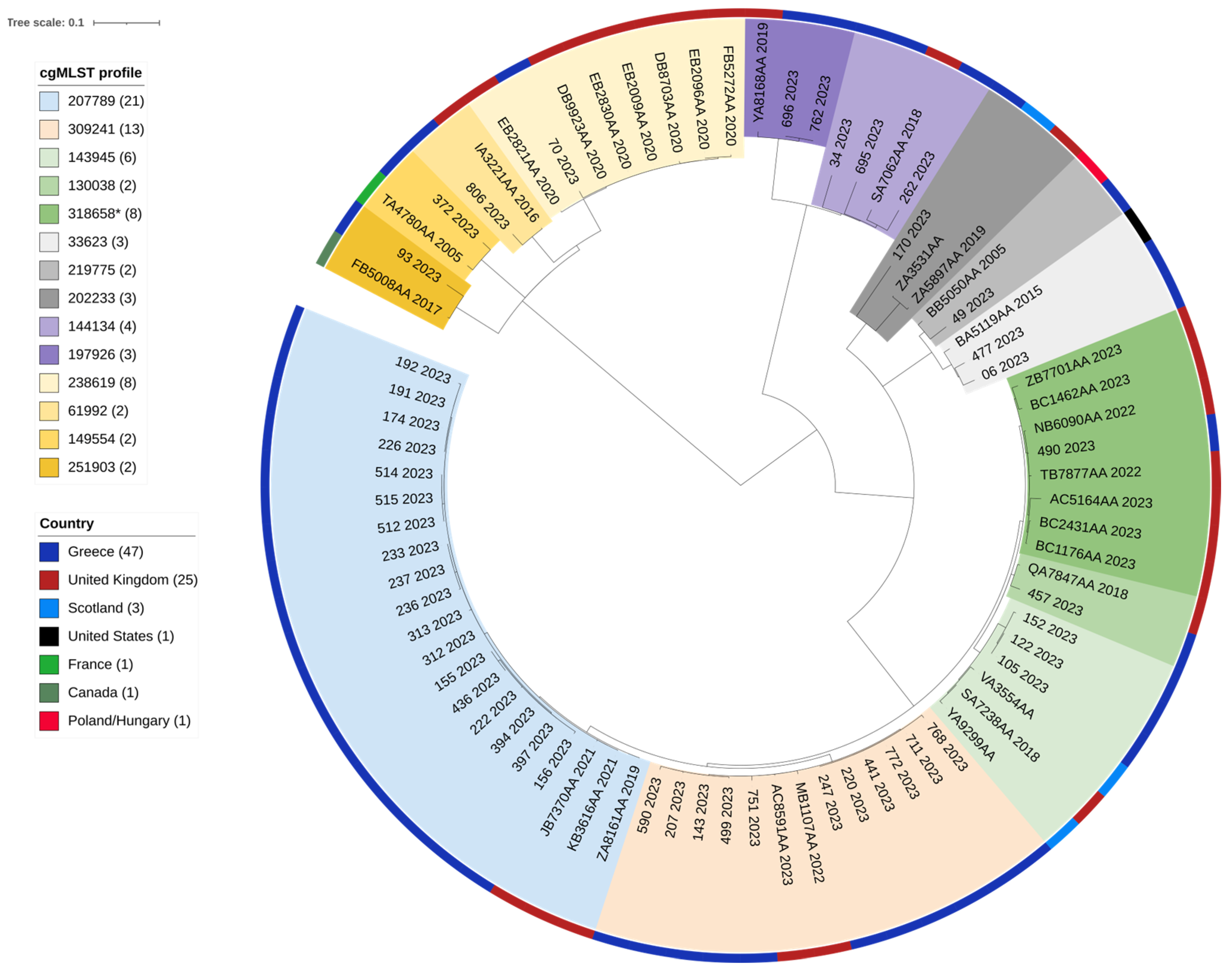
2.5. Assessment of the Genetic Relatedness of Greek Isolates in the Context of EnteroBase
3. Discussion
4. Materials and Methods
4.1. Sample Collection, Serotyping, and Antimicrobial Susceptibility Testing (AST)
4.2. Whole-Genome Sequencing (WGS)
4.3. Bioinformatic Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiology Reference Group 2007–2015; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Kadhum, W.R.; Sviridova, L.; Snegirev, D. The analysis of Salmonella’s ability to survive in different external environments. Biostat. Epidemiol. 2023, 7, e2265277. [Google Scholar] [CrossRef]
- Finn, S.; Condell, O.; McClure, P.; Amézquita, A.; Fanning, S. Mechanisms of survival, responses and sources of Salmonella in low-moisture environments. Front. Microbiol. 2013, 4, 331. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention Control. The European Union One Health 2022 Zoonoses Report. EFSA J. 2023, 21, e8442. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Salmonellosis. Available online: https://www.ecdc.europa.eu/en/salmonellosis (accessed on 10 June 2024).
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M.; International Collaboration on Enteric Disease ‘Burden of Illness’ Studies. The Global Burden of Nontyphoidal Salmonella Gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention Control. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, e07666. [Google Scholar] [CrossRef]
- Almansour, A.M.; Alhadlaq, M.A.; Alzahrani, K.O.; Mukhtar, L.E.; Alharbi, A.L.; Alajel, S.M. The Silent Threat: Antimicrobial-Resistant Pathogens in Food-Producing Animals and Their Impact on Public Health. Microorganisms 2023, 11, 2127. [Google Scholar] [CrossRef]
- Fastl, C.; De Carvalho Ferreira, H.C.; Babo Martins, S.; Sucena Afonso, J.; di Bari, C.; Venkateswaran, N.; Pires, S.M.; Mughini-Gras, L.; Huntington, B.; Rushton, J.; et al. Animal sources of antimicrobial-resistant bacterial infections in humans: A systematic review. Epidemiol. Infect. 2023, 151, e143. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, Y.; Mulchandani, R.; Van Boeckel, T.P. Global surveillance of antimicrobial resistance in food animals using priority drugs maps. Nat. Commun. 2024, 15, 763. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2021–2022. EFSA J. 2024, 22, e8583. [Google Scholar] [CrossRef]
- Teklemariam, A.D.; Al-Hindi, R.R.; Albiheyri, R.S.; Alharbi, M.G.; Alghamdi, M.A.; Filimban, A.A.R.; Al Mutiri, A.S.; Al-Alyani, A.M.; Alseghayer, M.S.; Almaneea, A.M.; et al. Human Salmonellosis: A Continuous Global Threat in the Farm-to-Fork Food Safety Continuum. Foods 2023, 12, 1756. [Google Scholar] [CrossRef]
- World Health Organization. Salmonella (Non-Typhoidal). Available online: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) (accessed on 10 June 2024).
- Rabsch, W.; Hargis, B.M.; Tsolis, R.M.; Kingsley, R.A.; Hinz, K.H.; Tschäpe, H.; Bäumler, A.J. Competitive exclusion of Salmonella enteritidis by Salmonella gallinarum in poultry. Emerg Infect Dis. 2000, 6, 443–448. [Google Scholar] [CrossRef]
- Hendriksen, R.S.; Vieira, A.R.; Karlsmose, S.; Lo Fo Wong, D.M.A.; Jensen, A.B.; Wegener, H.C.; Aarestrup, F.M. Global Monitoring of Salmonella Serovar Distribution from the World Health Organization Global Foodborne Infections Network Country Data Bank: Results of Quality Assured Laboratories from 2001 to 2007. Foodborne Pathog. Dis. 2011, 8, 887–900. [Google Scholar] [CrossRef]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef]
- Gantois, I.; Ducatelle, R.; Pasmans, F.; Haesebrouck, F.; Gast, R.; Humphrey, T.J.; Van Immerseel, F. Mechanisms of egg contamination by Salmonella Enteritidis. FEMS Microbiol. Rev. 2009, 33, 718–738. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, X.; Ding, X.; Bin, P.; Zhu, G. The vertical transmission of Salmonella Enteritidis in a One-Health context. One Health 2023, 16, 100469. [Google Scholar] [CrossRef]
- De Reu, K.; Grijspeerdt, K.; Messens, W.; Heyndrickx, M.; Uyttendaele, M.; Debevere, J.; Herman, L. Eggshell factors influencing eggshell penetration and whole egg contamination by different bacteria, including Salmonella enteritidis. Int. J. Food Microbiol. 2006, 112, 253–260. [Google Scholar] [CrossRef]
- Hofer, U. Salmonella Enteritidis: Chicken or egg? Nat. Rev. Microbiol. 2021, 19, 682. [Google Scholar] [CrossRef]
- Li, S.; He, Y.; Mann, D.A.; Deng, X. Global spread of Salmonella Enteritidis via centralized sourcing and international trade of poultry breeding stocks. Nat. Commun. 2021, 12, 5109. [Google Scholar] [CrossRef]
- Miyamoto, T.; Baba, E.; Tanaka, T.; Sasai, K.; Fukata, T.; Arakawa, A. Salmonella enteritidis contamination of eggs from hens inoculated by vaginal, cloacal, and intravenous routes. Avian Dis. 1997, 41, 296–303. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention Control. Multi-country outbreak of Salmonella Enteritidis infections linked to Polish eggs. EFSA Support. Publ. 2017, 14, 1353E. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention Control and European Food Safety Authority. Multi-country outbreak of Salmonella Enteritidis sequence type (ST)11 infections linked to eggs and egg products—8 February 2022. EFSA Support. Publ. 2022, 19, 7180E. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention Control and European Food Safety Authority. Three clusters of Salmonella Enteritidis ST11 infections linked to chicken meat and chicken meat products. EFSA Support. Publ. 2023, 20, 8388E. [Google Scholar] [CrossRef]
- World Health Organization. Whole Genome Sequencing for Foodborne Disease Surveillance: Landscape Paper; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Nadon, C.; Van Walle, I.; Gerner-Smidt, P.; Campos, J.; Chinen, I.; Concepcion-Acevedo, J.; Gilpin, B.; Smith, A.M.; Man Kam, K.; Perez, E.; et al. PulseNet International: Vision for the implementation of whole genome sequencing (WGS) for global food-borne disease surveillance. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. Eur. Commun. Dis. Bull. 2017, 22. [Google Scholar] [CrossRef]
- National Public Health Organization; Department of Food-Borne and Water-Borne Diseases. Epidemiological Data for Salmonellosis (Non-Typhoid/Paratyphoid) in Greece, 2004–2022, Mandatory Notification System. Available online: https://eody.gov.gr/wp-content/uploads/2023/06/epidemiological-data-for-salmonellosis_2004-2022.pdf (accessed on 10 June 2024).
- Mellou, K.; Gkova, M.; Panagiotidou, E.; Tzani, M.; Sideroglou, T.; Mandilara, G. Diversity and Resistance Profiles of Human Non-typhoidal Salmonella spp. in Greece, 2003–2020. Antibiotics 2021, 10, 983. [Google Scholar] [CrossRef]
- Silva, M.; Machado, M.P.; Silva, D.N.; Rossi, M.; Moran-Gilad, J.; Santos, S.; Ramirez, M.; Carriço, J.A. chewBBACA: A complete suite for gene-by-gene schema creation and strain identification. Microb. Genom. 2018, 4, e000166. [Google Scholar] [CrossRef]
- Interactive Tree of Life (ITOL) v3: An Online Tool for the Display and Annotation of Phylogenetic and Other Trees. Available online: https://itol.embl.de/ (accessed on 10 June 2024).
- Tseemann/Snippy: Rapid Haploid Variant Calling and Core Genome Alignment. Available online: https://github.com/tseemann/snippy (accessed on 10 June 2024).
- RAxML (Randomized Axelerated Maximum Likelihood), Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogeni. Available online: https://github.com/stamatak/standard-RAxML (accessed on 10 June 2024).
- Luo, L.; Payne, M.; Kaur, S.; Hu, D.; Cheney, L.; Octavia, S.; Wang, Q.; Tanaka, M.M.; Sintchenko, V.; Lan, R. Elucidation of global and national genomic epidemiology of Salmonella enterica serovar Enteritidis through multilevel genome typing. Microb. Genom. 2021, 7, e000605. [Google Scholar] [CrossRef]
- Liu, J.; Hou, Y.; Zhao, L.; Chen, G.; Chen, J.; Zhao, Q.; Ye, L.; Cui, S.; Wang, C. Antimicrobial Resistance and the Genomic Epidemiology of Multidrug-Resistant Salmonella enterica serovar Enteritidis ST11 in China. Front. Biosci. 2024, 29, 112. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, Z.; Zhao, H.; Li, S.; Zhuang, H.; Wang, J.; Li, R.; Zheng, W.; Zhu, H.; Xie, P.; et al. Antimicrobial Resistance and Genomic Characterization of Salmonella Isolated from Pigeons in China. Transbound. Emerg. Dis. 2024, 2024, 3315678. [Google Scholar] [CrossRef]
- Leão, C.; Silveira, L.; Usié, A.; Gião, J.; Clemente, L.; Themudo, P.; Amaro, A.; Pista, A. Genetic Diversity of Salmonella enterica subsp. enterica Serovar Enteritidis from Human and Non-Human Sources in Portugal. Pathogens 2024, 13, 112. [Google Scholar] [CrossRef]
- Foley, S.L.; Kaldhone, P.R.; Ricke, S.C.; Han, J. Incompatibility Group I1 (IncI1) Plasmids: Their Genetics, Biology, and Public Health Relevance. Microbiol. Mol. Biol. Rev. MMBR 2021, 85, e00031-20. [Google Scholar] [CrossRef] [PubMed]
- Nohejl, T.; Valcek, A.; Papousek, I.; Palkovicova, J.; Wailan, A.M.; Pratova, H.; Minoia, M.; Dolejska, M. Genomic analysis of qnr-harbouring IncX plasmids and their transferability within different hosts under induced stress. BMC Microbiol. 2022, 22, 136. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.D.M.; Ziora, Z.M.; Blaskovich, M.A.T. Quinolone antibiotics. MedChemComm 2019, 10, 1719–1739. [Google Scholar] [CrossRef] [PubMed]
- Gouvêa, R.; Santos, F.F.d.; Aquino, M.H.C.d.; Pereira, V.d.A. Fluoroquinolones in industrial poultry production, bacterial resistance and food residues: A review. Braz. J. Poult. Sci. 2015, 17, 619–627. [Google Scholar] [CrossRef]
- Fei, Z.; Song, S.; Yang, X.; Jiang, D.; Gao, J.; Yang, D. Occurrence and Risk Assessment of Fluoroquinolone Residues in Chicken and Pork in China. Antibiotics 2022, 11, 1292. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C. Positive Association between the Use of Quinolones in Food Animals and the Prevalence of Fluoroquinolone Resistance in E. coli and K. pneumoniae, A. baumannii and P. aeruginosa: A Global Ecological Analysis. Antibiotics 2021, 10, 1193. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention Control, European Food Safety Authority; European Medicines Agency. Antimicrobial consumption and resistance in bacteria from humans and food-producing animals. EFSA J. 2024, 22, e8589. [Google Scholar] [CrossRef]
- Jacoby, G.A. Mechanisms of resistance to quinolones. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2005, 41 (Suppl. S2), S120–S126. [Google Scholar] [CrossRef] [PubMed]
- Vidovic, S.; An, R.; Rendahl, A. Molecular and Physiological Characterization of Fluoroquinolone-Highly Resistant Salmonella Enteritidis Strains. Front. Microbiol. 2019, 10, 729. [Google Scholar] [CrossRef]
- Baker, D.J.; Robbins, A.; Newman, J.; Anand, M.; Wolfgang, W.J.; Mendez-Vallellanes, D.V.; Wirth, S.E.; Mingle, L.A. Challenges Associated with Investigating Salmonella Enteritidis with Low Genomic Diversity in New York State: The Impact of Adjusting Analytical Methods and Correlation with Epidemiological Data. Foodborne Pathog. Dis. 2023, 20, 230–236. [Google Scholar] [CrossRef]
- Xian, Z.; Li, S.; Mann, D.A.; Huang, Y.; Xu, F.; Wu, X.; Tang, S.; Zhang, G.; Stevenson, A.; Ge, C.; et al. Subtyping Evaluation of Salmonella Enteritidis Using Single Nucleotide Polymorphism and Core Genome Multilocus Sequence Typing with Nanopore Reads. Appl. Environ. Microbiol. 2022, 88, e00785-00722. [Google Scholar] [CrossRef] [PubMed]
- Schwan, C.L.; Lomonaco, S.; Bastos, L.M.; Cook, P.W.; Maher, J.; Trinetta, V.; Bhullar, M.; Phebus, R.K.; Gragg, S.; Kastner, J.; et al. Genotypic and Phenotypic Characterization of Antimicrobial Resistance Profiles in Non-typhoidal Salmonella enterica Strains Isolated from Cambodian Informal Markets. Front. Microbiol. 2021, 12, 711472. [Google Scholar] [CrossRef] [PubMed]
- Grimont, P.A.; Weill, F.X. Antigenic Formulae of the Salmonella Serovars, 9th ed.; WHO Collaborating Centre for Reference and Research on Salmonella and Institut Pasteur: Paris, France, 2007. [Google Scholar]
- EUCAST—European Committee on Antimicrobial Susceptibility Testing. Available online: https://www.eucast.org/ (accessed on 10 June 2024).
- AMR Workflow Documentation. Available online: https://github.com/phac-nml/staramr (accessed on 10 June 2024).
- Enterobase. cgMLST Scheme Documentation. Available online: https://enterobase.warwick.ac.uk (accessed on 10 June 2024).
- Roer, L.; Hendriksen, R.S.; Leekitcharoenphon, P.; Lukjancenko, O.; Kaas, R.S.; Hasman, H.; Aarestrup, F.M. Is the Evolution of Salmonella enterica subsp. enterica Linked to Restriction-Modification Systems? mSystems 2016, 1, e00009-16. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Alikhan, N.F.; Mohamed, K.; Fan, Y.; Achtman, M. The EnteroBase user’s guide, with case studies on Salmonella transmissions, Yersinia pestis phylogeny, and Escherichia core genomic diversity. Genome Res. 2020, 30, 138–152. [Google Scholar] [CrossRef]
- Achtman, M.; Zhou, Z.; Alikhan, N.F.; Tyne, W.; Parkhill, J.; Cormican, M.; Chiou, C.S.; Torpdahl, M.; Litrup, E.; Prendergast, D.M.; et al. Genomic diversity of Salmonella enterica-The UoWUCC 10K genomes project. Wellcome Open Res. 2020, 5, 223. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| NRCS ID | Specimen | Age Group of Cases | AΜR Genotype | Sequence Type (ST) | cgMST Profile | Predicted AMR Phenotype | Plasmids | SPIs * |
|---|---|---|---|---|---|---|---|---|
| 155 | Stool | >65 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 156 | Stool | 6–14 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | None | 1,2,3,4,5,10,13,14 and C63PI |
| 174 | Blood | 15–64 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 191 | Stool | 0–11 months | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 192 | Stool | 0–11 months | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 222 | Stool | 6–14 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 226 | Stool | 1–5 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 233 ** A | Stool | 1–5 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 236 ** A | Stool | 15–64 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 237 ** A | Stool | 15–64 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 312 | Stool | unknown | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 313 | Stool | unknown | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 394 | Stool | 6–14 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 397 | Stool | 15–64 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 436 | Stool | 6–14 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 512 ** B | Stool | unknown | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 514 ** B | Stool | 15–64 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 515 ** B | Stool | 15–64 years | gyrA (D87Y) | 11 | 207789 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 143 | Stool | 1–5 years | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 207 | Stool | 6–14 years | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 220 | Stool | 6–14 years | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 247 | Stool | 1–5 years | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 441 | Blood | >65 years | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 499 | Stool | 15–64 years | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 590 | unknown | unknown | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 711 | Blood | unknown | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 751 | Blood | 0–11 months | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 768 | Stool | 1–5 years | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | None | 1,2,3,4,5,10,13,14 and C63PI |
| 772 | Stool | 6–14 years | gyrA (D87Y) | 11 | 309241 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 457 | Stool | unknown | None | 11 | 130038 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 105 | Stool | 1–5 years | None | 11 | 143945 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 122 | Stool | 6–14 years | None | 11 | 143945 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 152 | Stool | 15–64 years | None | 11 | 143945 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 490 | Stool | 6–14 years | None | 11 | 318658 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 170 | Stool | 1–5 years | None | 11 | 202233 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 6 | Blood | >65 years | None | 11 | 33623 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 477 | Blood | unknown | None | 11 | 33623 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 49 | Blood | >65 years | None | 11 | 219775 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 34 | Blood | >65 years | None | 11 | 144134 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 262 | Stool | unknown | None | 11 | 144134 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 695 | Blood | >65 years | None | 11 | 144134 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 696 | Stool | 6–14 years | None | 11 | 197926 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 762 | Stool | 6–14 years | None | 11 | 197926 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 372 | Stool | unknown | None | 11 | 149554 | Susceptible | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 70 | Stool | 15–64 years | gyrA (D87N) | 11 | 238619 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,5,10,13,14 and C63PI |
| 806 | Blood | >65 years | gyrA (S83Y) | 11 | 61992 | ciprofloxacin, nalidixic acid | IncFIB(S), IncFII(S) | 1,2,3,4,5,10,13,14 and C63PI |
| 93 | Stool | 1–5 years | aph(3′)-Ia, qnrS13, tet(A) | 11 | 251903 | kanamycin, ciprofloxacin, tetracycline | IncFIB(S), IncFII(S), IncI1-I(Alpha), IncX1 | 1,2,3,4,5,10,13,14 and C63PI |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polemis, M.; Sideroglou, T.; Chrysostomou, A.; Mandilara, G.D. First Data on WGS-Based Typing and Antimicrobial Resistance of Human Salmonella Enteritidis Isolates in Greece. Antibiotics 2024, 13, 708. https://doi.org/10.3390/antibiotics13080708
Polemis M, Sideroglou T, Chrysostomou A, Mandilara GD. First Data on WGS-Based Typing and Antimicrobial Resistance of Human Salmonella Enteritidis Isolates in Greece. Antibiotics. 2024; 13(8):708. https://doi.org/10.3390/antibiotics13080708
Chicago/Turabian StylePolemis, Michalis, Theologia Sideroglou, Anthi Chrysostomou, and Georgia D. Mandilara. 2024. "First Data on WGS-Based Typing and Antimicrobial Resistance of Human Salmonella Enteritidis Isolates in Greece" Antibiotics 13, no. 8: 708. https://doi.org/10.3390/antibiotics13080708