
Microplastic-Mediated Transfer of Tetracycline Resistance: Unveiling the Role of Mussels in Marine Ecosystems

 ,
,  , ,
, ,  and
and {kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
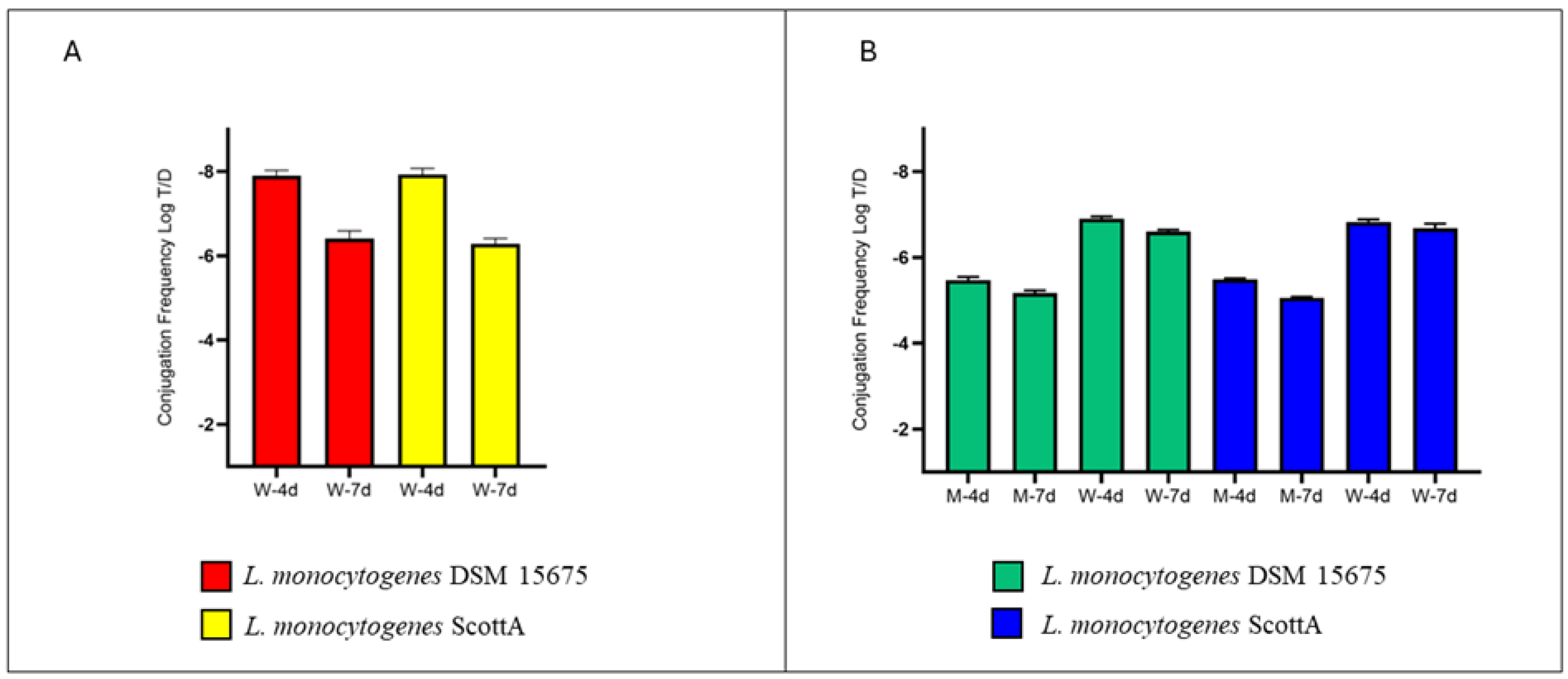
2.1. Involvement of MPs in ARG Exchange inside Mussels
2.2. The Role of MGE of L. monocytogenes in HGT
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. L. monocytogenes and E. faecium Biofilm Formation: Microtiter Assay and Microplastic Colonization
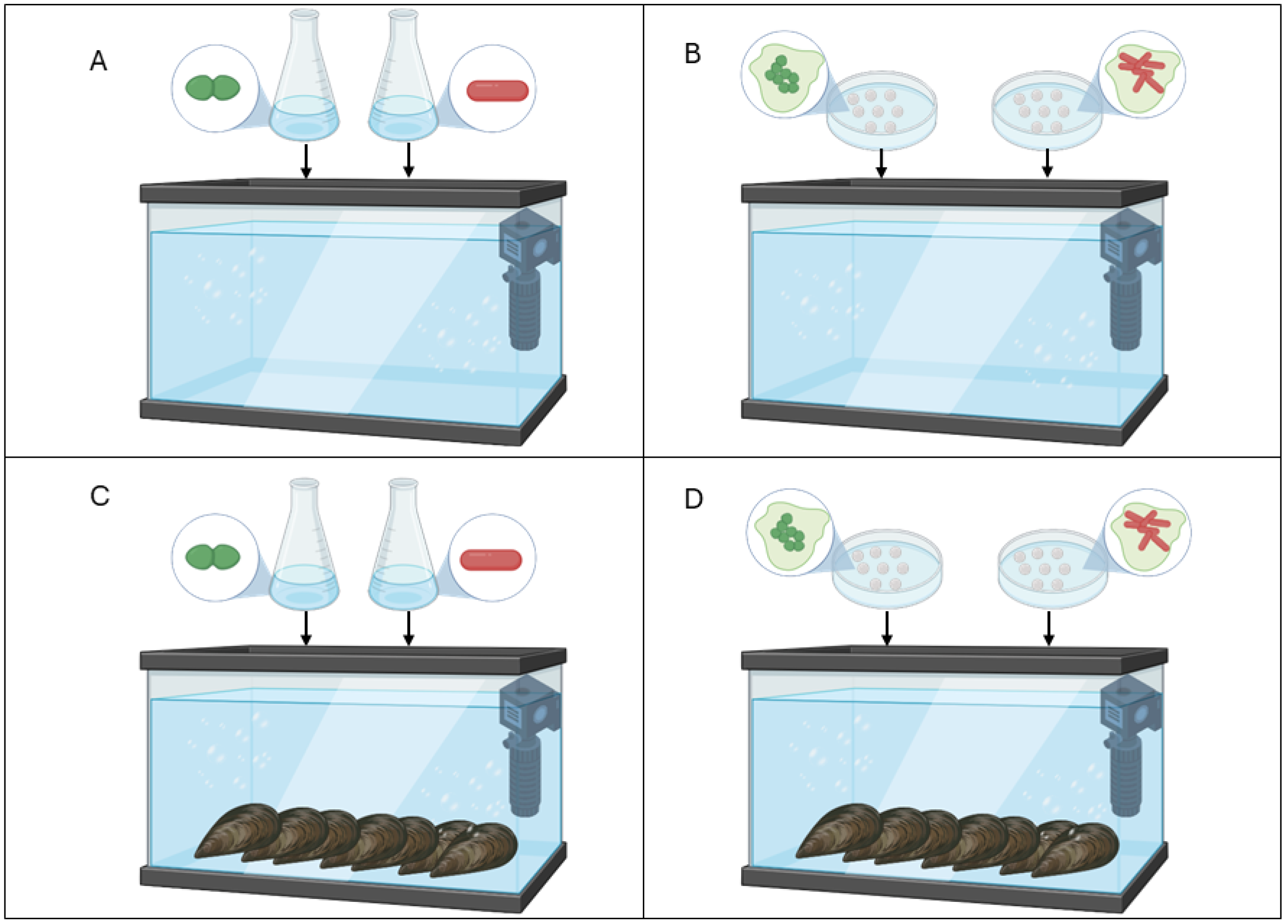
4.3. Experimental Design
4.4. Microbiological and Molecular Analysis for Transconjugants Confirmation
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aslam, B.; Khurshid, M.; Arshad, M.I.; Muzammil, S.; Rasool, M.; Yasmeen, N.; Shah, T.; Chaudhry, T.H.; Rasool, M.H.; Shahid, A.; et al. Antibiotic resistance: One health one world outlook. Front. Cell. Infect. Microbiol. 2021, 11, 771510. [Google Scholar] [CrossRef] [PubMed]
- Coque, T.M.; Cantón, R.; Pérez-Cobas, A.E.; Fernández-De-Bobadilla, M.D.; Baquero, F. Antimicrobial resistance in the global health network: Known unknowns and challenges for efficient responses in the 21st century. Microorganisms 2023, 11, 1050. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; Flach, C.-F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2021, 20, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [PubMed]
- Bean, E.L.; McLellan, L.K.; Grossman, A.D. Activation of the integrative and conjugative element Tn916 causes growth arrest and death of host bacteria. PLoS Genet. 2022, 18, e1010467. [Google Scholar] [CrossRef] [PubMed]
- Vinayamohan, P.G.; Pellissery, A.J.; Venkitanarayanan, K. Role of horizontal gene transfer in the dissemination of antimicrobial resistance in food animal production. Curr. Opin. Food Sci. 2022, 47, 100882. [Google Scholar] [CrossRef]
- Aminov, R.I. Horizontal gene exchange in environmental microbiota. Front. Microbiol. 2011, 2, 158. [Google Scholar] [CrossRef] [PubMed]
- Hatosy, S.M.; Martiny, A.C. The ocean as a global reservoir of antibiotic resistance genes. Appl. Environ. Microbiol. 2015, 81, 7593–7599. [Google Scholar] [CrossRef]
- Li, C.; Li, X.; Bank, M.S.; Dong, T.; Fang, J.K.-H.; Leusch, F.D.L.; Rillig, M.C.; Wang, J.; Wang, L.; Xia, Y.; et al. The “Microplastome”—A holistic perspective to capture the real-world ecology of microplastics. Environ. Sci. Technol. 2024, 58, 4060–4069. [Google Scholar] [CrossRef]
- Moore, C.; Moore, S.; Weisberg, S.; Lattin, G.; Zellers, A. A comparison of neustonic plastic and zooplankton abundance in southern California’s coastal waters. Mar. Pollut. Bull. 2002, 44, 1035–1038. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, X.; Qiu, P.; Chen, B.; Xu, C.; Dong, W.; Liu, T. Microplastics can selectively enrich intracellular and extracellular antibiotic resistant genes and shape different microbial communities in aquatic systems. Sci. Total. Environ. 2022, 822, 153488. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, L.C.M.; van der Zwet, J.; Damsteeg, J.-W.; Slat, B.; Andrady, A.; Reisser, J. River plastic emissions to the world’s oceans. Nat. Commun. 2017, 8, 15611. [Google Scholar] [CrossRef] [PubMed]
- Barboza, L.G.A.; Lopes, C.; Oliveira, P.; Bessa, F.; Otero, V.; Henriques, B.; Raimundo, J.; Caetano, M.; Vale, C.; Guilhermino, L. Microplastics in wild fish from North East Atlantic Ocean and its potential for causing neurotoxic effects, lipid oxidative damage, and human health risks associated with ingestion exposure. Sci. Total. Environ. 2019, 717, 134625. [Google Scholar] [CrossRef] [PubMed]
- Avio, C.G.; Pittura, L.; D’errico, G.; Abel, S.; Amorello, S.; Marino, G.; Gorbi, S.; Regoli, F. Distribution and characterization of microplastic particles and textile microfibers in Adriatic food webs: General insights for biomonitoring strategies. Environ. Pollut. 2019, 258, 113766. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Green, C.; Reynolds, A.; Shi, H.; Rotchell, J.M. Microplastics in mussels sampled from coastal waters and supermarkets in the United Kingdom. Environ. Pollut. 2018, 241, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Bueris, V.; Sellera, F.P.; Fuga, B.; Sano, E.; Carvalho, M.P.N.; Couto, S.C.F.; Moura, Q.; Lincopan, N. Convergence of virulence and resistance in international clones of WHO critical priority enterobacterales isolated from marine bivalves. Sci. Rep. 2022, 12, 5707. [Google Scholar] [CrossRef]
- Hammerum, A.M. Enterococci of Animal Origin and Their Significance for Public Health. Clin. Microbiol. Infect. 2012, 18, 619–625. [Google Scholar] [CrossRef]
- Klare, I.; Konstabel, C.; Badstübner, D.; Werner, G.; Witte, W. Occurrence and Spread of Antibiotic Resistances in Enterococcus Faecium. Int. J. Food Microbiol. 2003, 88, 269–290. [Google Scholar] [CrossRef] [PubMed]
- Hegstad, K.; Mikalsen, T.; Coque, T.M.; Werner, G.; Sundsfjord, A. Mobile Genetic Elements and Their Contribution to the Emergence of Antimicrobial Resistant Enterococcus Faecalis and Enterococcus Faecium. Clin. Microbiol. Infect. 2010, 16, 541–554. [Google Scholar] [CrossRef]
- Daza, M.V.B.; Milani, G.; Cortimiglia, C.; Pietta, E.; Bassi, D.; Cocconcelli, P.S. Genomic insights of Enterococcus faecium UC7251, a multi-drug resistant strain from ready-to-eat food, highlight the risk of antimicrobial resistance in the food chain. Front. Microbiol. 2022, 13, 894241. [Google Scholar] [CrossRef]
- Haubert, L.; da Cunha, C.E.P.; Lopes, G.V.; da Silva, W.P. Food isolate Listeria monocytogenes harboring tetM gene plasmid-mediated exchangeable to Enterococcus faecalis on the surface of processed cheese. Food Res. Int. 2018, 107, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, A.J.; Gajewska, J.; Chajęcka-Wierzchowska, W.; Załuski, D.; Zadernowska, A. Prevalence of Listeria monocytogenes and other Listeria species in fish, fish products and fish processing environment: A systematic review and meta-analysis. Sci. Total. Environ. 2024, 907, 167912. [Google Scholar] [CrossRef] [PubMed]
- Marinho, A.; Martins, P.; Ditmer, E.; D’Azevedo, P.; Frazzon, J.; Der Sand, S.; Frazzon, A. Biofilm formation on polystyrene under different temperatures by antibiotic resistant Enterococcus faecalis and Enterococcus faecium isolated from food. Braz. J. Microbiol. 2013, 44, 423–426. [Google Scholar] [CrossRef]
- Di Ciccio, P.; Rubiola, S.; Panebianco, F.; Lomonaco, S.; Allard, M.; Bianchi, D.M.; Civera, T.; Chiesa, F. Biofilm formation and genomic features of Listeria monocytogenes strains isolated from meat and dairy industries located in Piedmont (Italy). Int. J. Food Microbiol. 2022, 378, 109784. [Google Scholar] [CrossRef] [PubMed]
- Șchiopu, P.; Toc, D.A.; Colosi, I.A.; Costache, C.; Ruospo, G.; Berar, G.; Gălbău, Ș.-G.; Ghilea, A.C.; Botan, A.; Pană, A.-G.; et al. An overview of the factors involved in biofilm production by the Enterococcus genus. Int. J. Mol. Sci. 2023, 24, 11577. [Google Scholar] [CrossRef]
- Trokter, M.; Waksman, G. Translocation through the conjugative type iv secretion system requires unfolding of its protein substrate. J. Bacteriol. 2018, 200, e00615-17. [Google Scholar] [CrossRef] [PubMed]
- Radisic, V.; Grevskott, D.H.; Junghardt, N.; Øvreås, L.; Marathe, N.P. Multidrug-resistant Enterococcus faecium strains enter the Norwegian marine environment through treated sewage. Microbiologyopen 2024, 13, e1397. [Google Scholar] [CrossRef] [PubMed]
- Heim, A.v.B.T.; Janice, J.; Bjørnholt, J.V.; Lunestad, B.T.; Hegstad, K.; Svanevik, C.S. Genomic insights into Enterococcus faecium isolates from marine bivalves highlight One Health concerns and healthcare linkages. Microb. Genom. 2023, 9, 001154. [Google Scholar] [CrossRef]
- Peratikos, P.; Tsitsos, A.; Damianos, A.; Kyritsi, M.A.; Hadjichristodoulou, C.; Soultos, N.; Economou, V. Listeria monocytogenes from marine fish and the seafood market environment in northern greece: Prevalence, molecular characterization, and antibiotic resistance. Appl. Sci. 2024, 14, 2725. [Google Scholar] [CrossRef]
- Mao, P.; Wang, Y.; Li, L.; Ji, S.; Li, P.; Liu, L.; Chen, J.; Sun, H.; Luo, X.; Ye, C. The isolation, genetic analysis and biofilm characteristics of Listeria spp. from the marine environment in China. Microorganisms 2023, 11, 2166. [Google Scholar] [CrossRef]
- Wang, H.; Xu, K.; Wang, J.; Feng, C.; Chen, Y.; Shi, J.; Ding, Y.; Deng, C.; Liu, X. Microplastic biofilm: An important microniche that may accelerate the spread of antibiotic resistance genes via natural transformation. J. Hazard. Mater. 2023, 459, 132085. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Pan, J.; Li, M.; Li, Y.; Bartlam, M.; Wang, Y. Selective enrichment of bacterial pathogens by microplastic biofilm. Water Res. 2019, 165, 114979. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, C.C.C.R. Marine biofilms: A successful microbial strategy with economic implications. Front. Mar. Sci. 2018, 5, 126. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, C.; Grohmann, E. Horizontal gene transfer of antibiotic resistance genes in biofilms. Antibiotics 2023, 12, 328. [Google Scholar] [CrossRef]
- Arias-Andres, M.; Klümper, U.; Rojas-Jimenez, K.; Grossart, H.-P. Microplastic pollution increases gene exchange in aquatic ecosystems. Environ. Pollut. 2018, 237, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Watkinson, A.; Murby, E.; Kolpin, D.; Costanzo, S. The occurrence of antibiotics in an urban watershed: From wastewater to drinking water. Sci. Total. Environ. 2009, 407, 2711–2723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Pei, J.; Zhang, R.; Wang, S.; Zeng, W.; Huang, D.; Wang, Y.; Zhang, Y.; Wang, Y.; Yu, K. Occurrence and distribution of antibiotics in mariculture farms, estuaries and the coast of the Beibu Gulf, China: Bioconcentration and diet safety of seafood. Ecotoxicol. Environ. Saf. 2018, 154, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Junaid, M.; Chen, G.; Wang, J. Interactions and associated resistance development mechanisms between microplastics, antibiotics and heavy metals in the aquaculture environment. Rev. Aquac. 2021, 14, 1028–1045. [Google Scholar] [CrossRef]
- Qu, X.; Su, L.; Li, H.; Liang, M.; Shi, H. Assessing the relationship between the abundance and properties of microplastics in water and in mussels. Sci. Total. Environ. 2017, 621, 679–686. [Google Scholar] [CrossRef]
- Digka, N.; Tsangaris, C.; Torre, M.; Anastasopoulou, A.; Zeri, C. Microplastics in mussels and fish from the Northern Ionian Sea. Mar. Pollut. Bull. 2018, 135, 30–40. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Jia, M.; Xiang, Y.; Song, B.; Xiong, W.; Cao, J.; Peng, H.; Yang, Y.; Wang, W.; Yang, Z.; et al. Biofilm on microplastics in aqueous environment: Physicochemical properties and environmental implications. J. Hazard. Mater. 2021, 424, 127286. [Google Scholar] [CrossRef] [PubMed]
- Sooriyakumar, P.; Bolan, N.; Kumar, M.; Singh, L.; Yu, Y.; Li, Y.; Weralupitiya, C.; Vithanage, M.; Ramanayaka, S.; Sarkar, B.; et al. Biofilm formation and its implications on the properties and fate of microplastics in aquatic environments: A review. J. Hazard. Mater. Adv. 2022, 6, 100077. [Google Scholar] [CrossRef]
- Wang, R.; Wu, J.; Jiang, N.; Lin, H.; An, F.; Wu, C.; Yue, X.; Shi, H.; Wu, R. Recent developments in horizontal gene transfer with the adaptive innovation of fermented foods. Crit. Rev. Food Sci. Nutr. 2022, 63, 569–584. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, X.; Xie, Y.; Bi, D.; Sun, J.; Li, J.; Tai, C.; Deng, Z.; Ou, H.-Y. ICEberg 2.0: An updated database of bacterial integrative and conjugative elements. Nucleic Acids Res. 2018, 47, D660–D665. [Google Scholar] [CrossRef]
- Hchaichi, I.; Bandini, F.; Spini, G.; Banni, M.; Cocconcelli, P.S.; Puglisi, E. Enterococcus faecalis and Vibrio harveyi colonize low-density polyethylene and biodegradable plastics under marine conditions. FEMS Microbiol. Lett. 2020, 367, fnaa125. [Google Scholar] [CrossRef]
- Palm, M.; Fransson, A.; Hultén, J.; Stenman, K.B.; Allouche, A.; Chiang, O.E.; Constandse, M.L.; van Dijk, K.J.; Icli, S.; Klimesova, B.; et al. The Effect of Heavy Metals on Conjugation Efficiency of an F-Plasmid in Escherichia coli. Antibiotics 2022, 11, 1123. [Google Scholar] [CrossRef]
- Zhou, X.; Jiao, X. Polymerase chain reaction detection of Listeria monocytogenes using oligonucleotide primers targeting actA gene. Food Control. 2005, 16, 125–130. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- World Health Organization. Laboratory Biosafety Manual, 3rd ed.; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milani, G.; Cortimiglia, C.; Belloso Daza, M.V.; Greco, E.; Bassi, D.; Cocconcelli, P.S. Microplastic-Mediated Transfer of Tetracycline Resistance: Unveiling the Role of Mussels in Marine Ecosystems. Antibiotics 2024, 13, 727. https://doi.org/10.3390/antibiotics13080727
Milani G, Cortimiglia C, Belloso Daza MV, Greco E, Bassi D, Cocconcelli PS. Microplastic-Mediated Transfer of Tetracycline Resistance: Unveiling the Role of Mussels in Marine Ecosystems. Antibiotics. 2024; 13(8):727. https://doi.org/10.3390/antibiotics13080727
Chicago/Turabian StyleMilani, Giovanni, Claudia Cortimiglia, Mireya Viviana Belloso Daza, Emanuele Greco, Daniela Bassi, and Pier Sandro Cocconcelli. 2024. "Microplastic-Mediated Transfer of Tetracycline Resistance: Unveiling the Role of Mussels in Marine Ecosystems" Antibiotics 13, no. 8: 727. https://doi.org/10.3390/antibiotics13080727
APA StyleMilani, G., Cortimiglia, C., Belloso Daza, M. V., Greco, E., Bassi, D., & Cocconcelli, P. S. (2024). Microplastic-Mediated Transfer of Tetracycline Resistance: Unveiling the Role of Mussels in Marine Ecosystems. Antibiotics, 13(8), 727. https://doi.org/10.3390/antibiotics13080727