
Bacteriophage Therapy on an In Vitro Wound Model and Synergistic Effects in Combination with Beta-Lactam Antibiotics

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
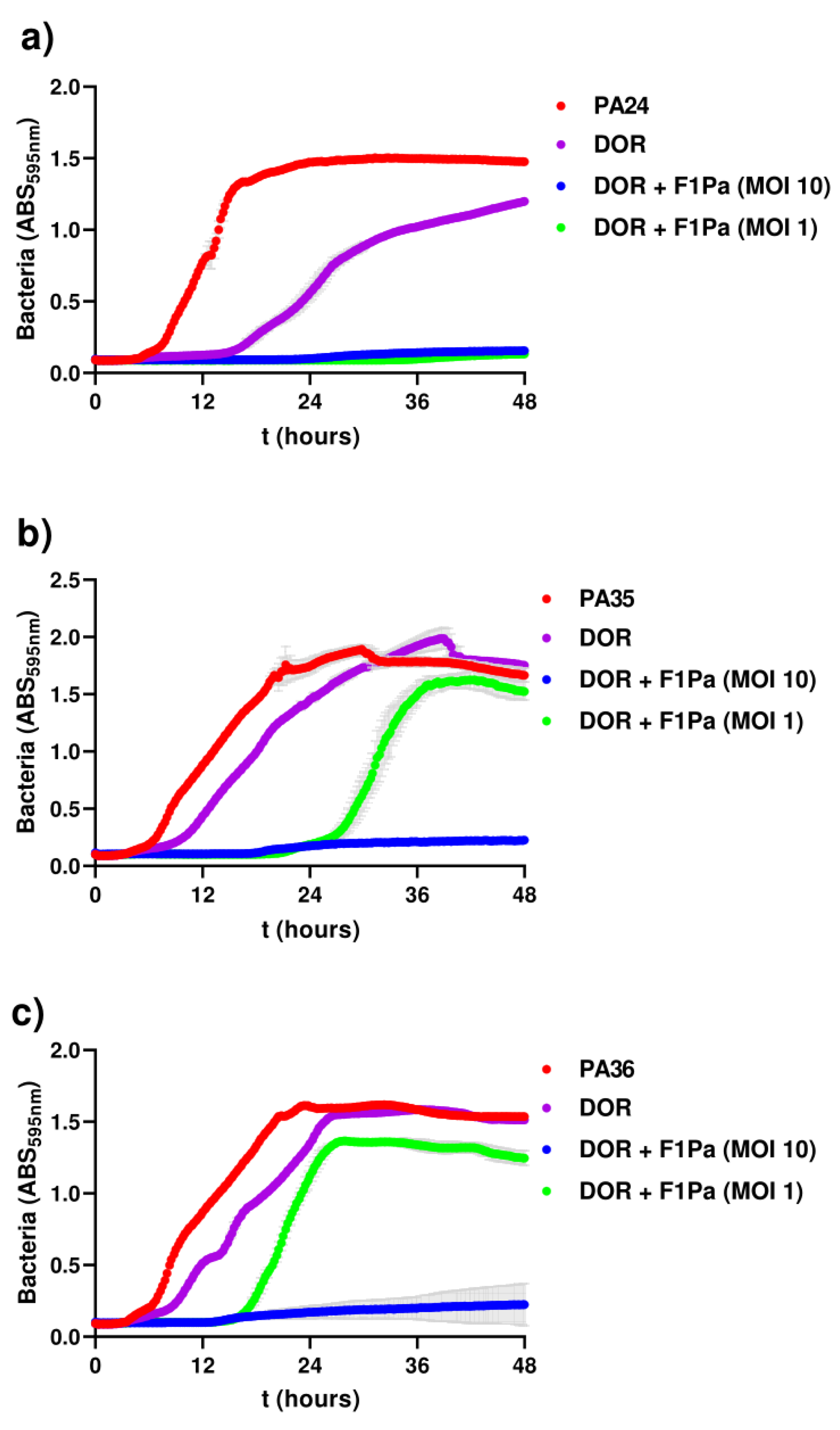
2.1. In Vitro Wound-like Model
2.2. Bacteriophage-Antibiotic Synergy
2.3. Phage–Antibiotic Inhibition Assays
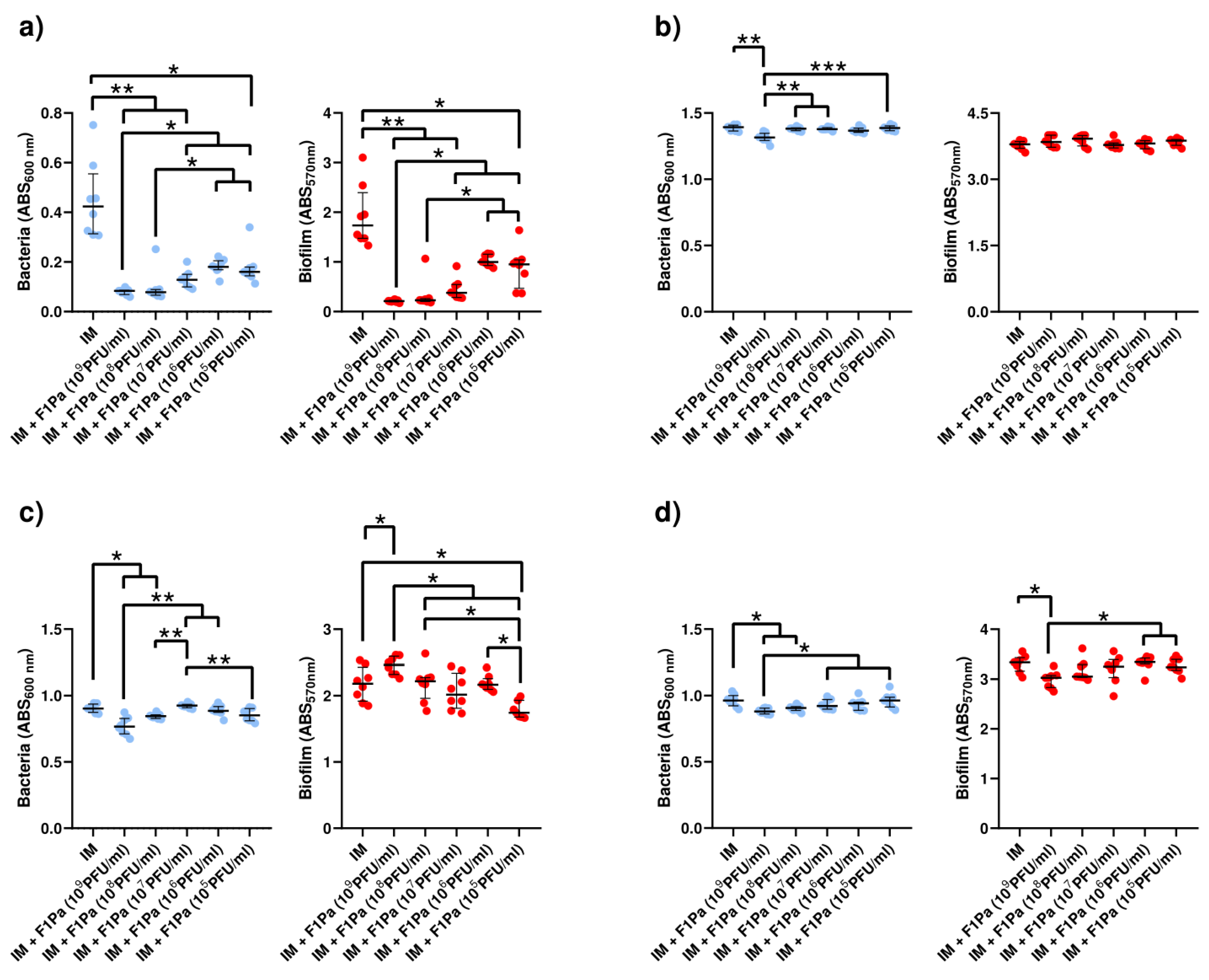
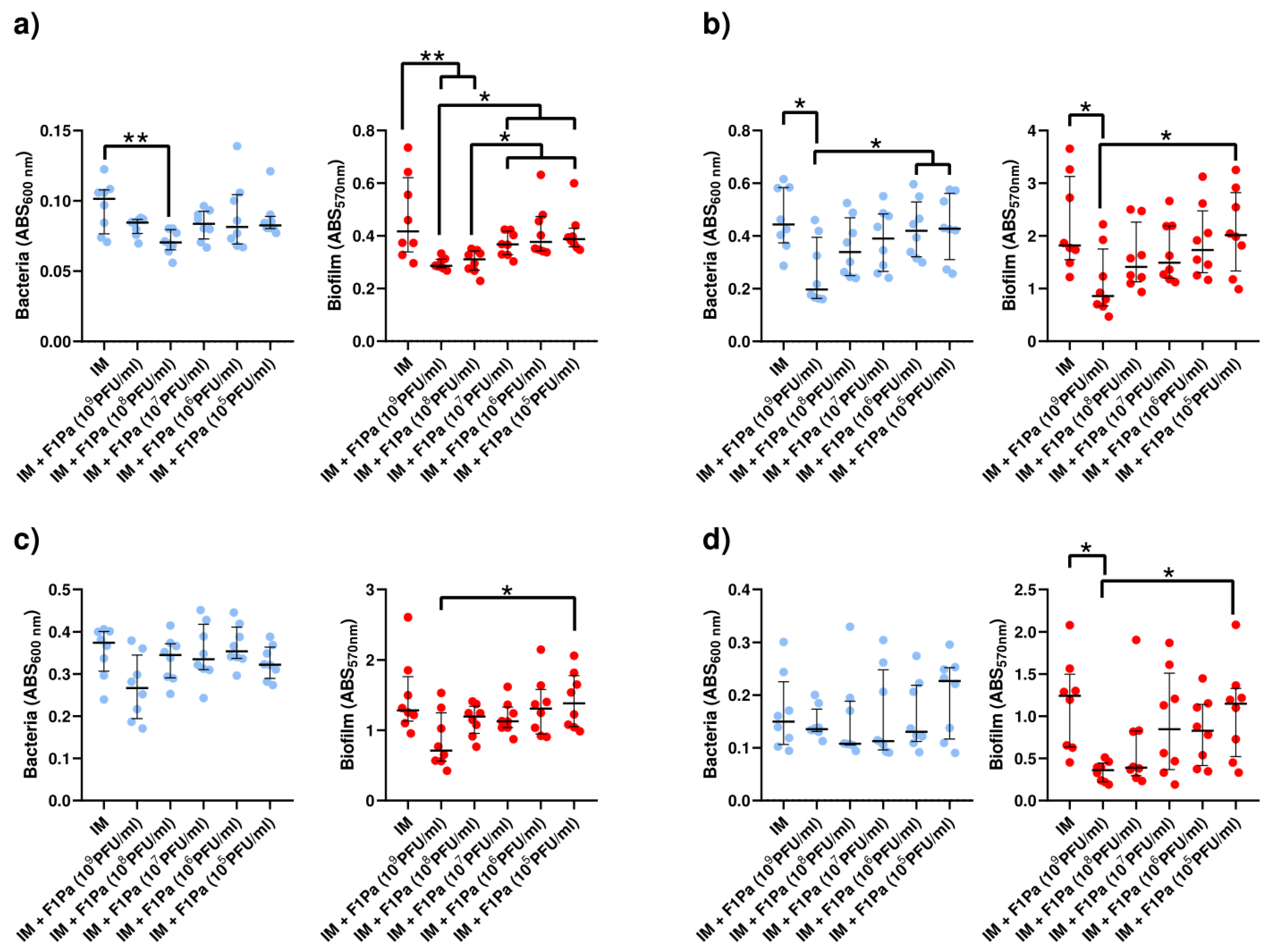
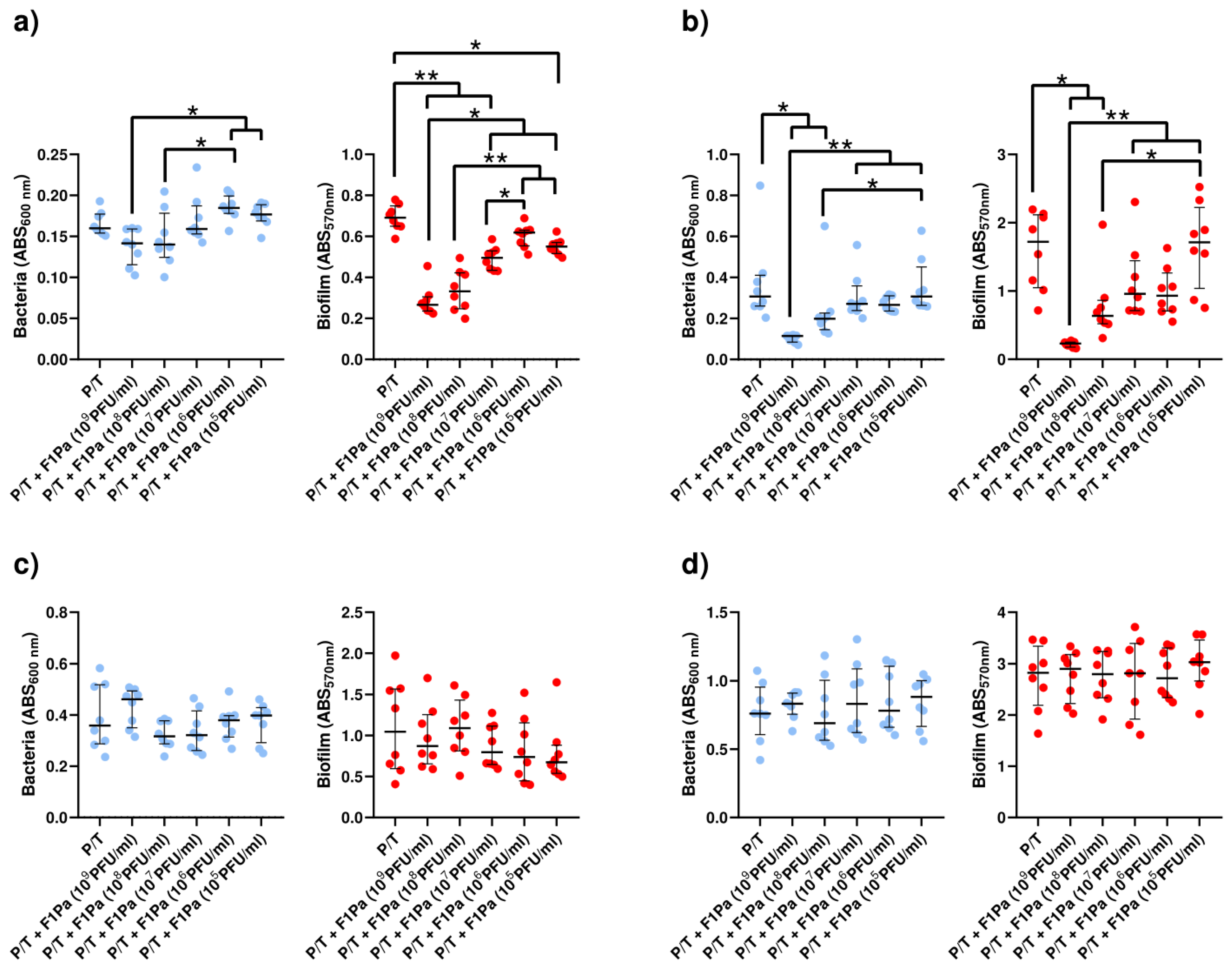
2.4. Phage–Antibiotic Effect on Pseudomonal Biofilms
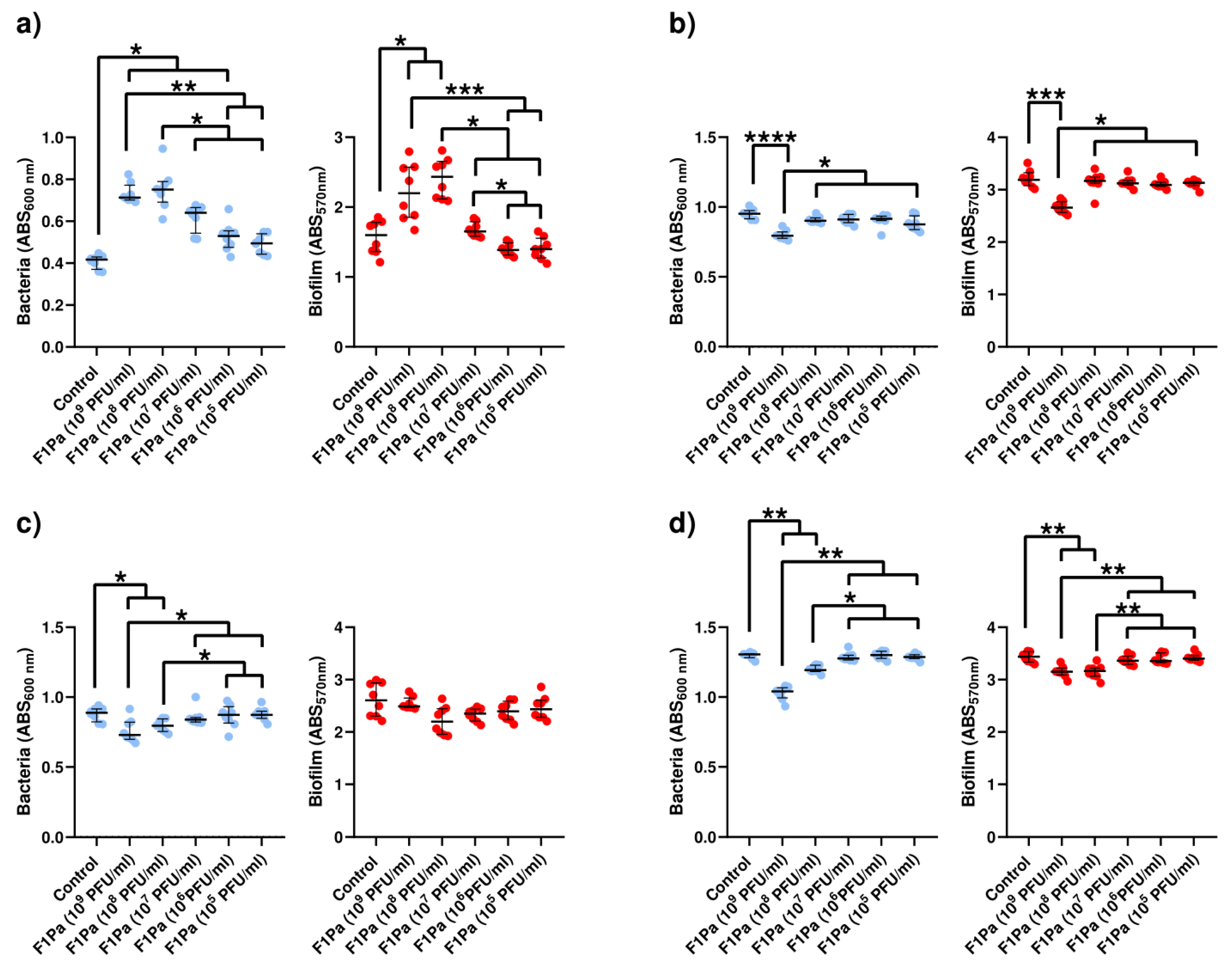
2.4.1. F1Pa
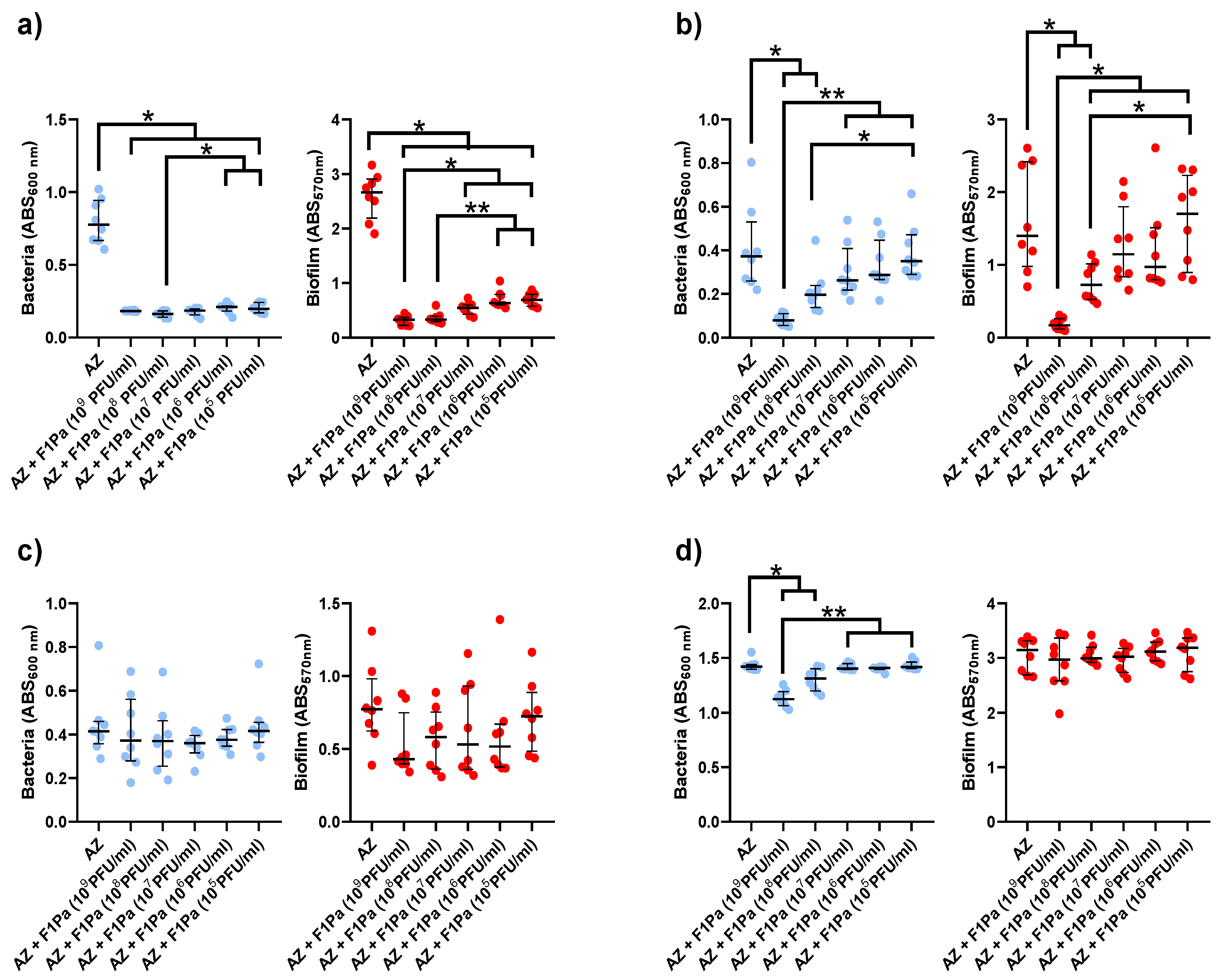
2.4.2. Aztreonam-F1Pa
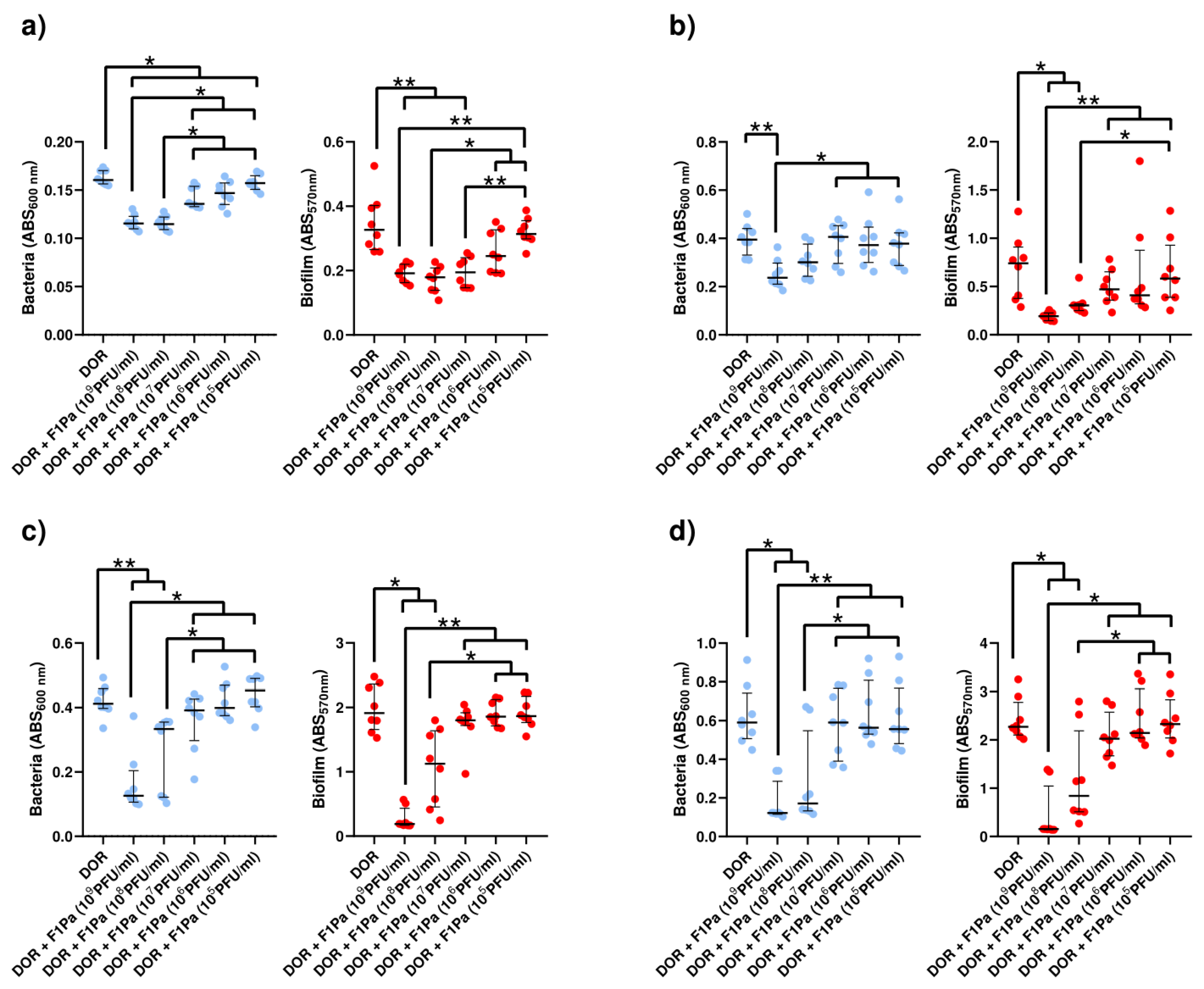
2.4.3. Doripenem–F1Pa
Doripenem (8.79 µg/mL)–F1Pa
Doripenem (23 µg/mL)-F1Pa
2.4.4. Imipenem-F1Pa
Imipenem (21 µg/mL)-F1Pa
Imipenem (39 µg/mL)–F1Pa
2.4.5. Meropenem–F1Pa
2.4.6. Piperacillin/Tazobactam–F1Pa
3. Discussion
4. Material and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Bacteriophage Isolation
4.3. Bacteriophage Propagation
4.4. Bacteriophage Titration
4.5. In Vitro Wound-like Medium
4.6. Bacteriophage–Antibiotic Interactions
4.7. Phage–Antibiotic Inhibition Assays
4.8. Phage–Antibiotic Effect on Pseudomonal Biofilm
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sen, C.K.; Gordillo, G.M.; Roy, S.; Kirsner, R.; Lambert, L.; Hunt, T.K.; Gottrup, F.; Gurtner, G.C.; Longaker, M.T. Human skin wounds: A major and snowballing threat to public health and the economy. Wound Repair. Regen. 2009, 17, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Aurora, A.; Le, T.D.; Akers, K.S.; Blyth, D.M.; Graybill, J.C.; Clemens, M.S.; Chung, K.K.; Rizzo, J.A. Recurrent Bacteremia: A 10-year retrospective study in combat-related burn casualties. Burns 2019, 45, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Azzopardi, E.A.; Azzopardi, E.; Camilleri, L.; Villapalos, J.; Boyce, D.E.; Dziewulski, P.; Dickson, W.A.; Whitaker, I.S. Gram negative wound infection in hospitalised adult burn patients—Systematic review and metanalysis-. PLoS ONE 2014, 9, e95042. [Google Scholar] [CrossRef] [PubMed]
- Suetens, C.; Kärki, T.; Diamantis, P. Point prevalence survey of healthcare-associated infections and antimicrobial use in european acute care hospitals 2022–2023; Publications Office of the European Union: Luxembourg, 2022. [Google Scholar] [CrossRef]
- Cwajda-białasik, J.; Mościcka, P.; Jawień, A.; Szewczyk, M.T. Microbiological status of venous leg ulcers and its predictors: A single-center cross-sectional study. Int. J. Environ. Res. Public Health 2021, 18, 12965. [Google Scholar] [CrossRef] [PubMed]
- Fayolle, M.; Morsli, M.; Gelis, A.; Chateauraynaud, M.; Yahiaoui-Martinez, A.; Sotto, A.; Lavigne, J.P.; Dunyach-Remy, C. The persistence of Staphylococcus aureus in pressure ulcers: A colonising role. Genes 2021, 12, 1883. [Google Scholar] [CrossRef]
- Mahnic, A.; Breznik, V.; Bombek Ihan, M.; Rupnik, M. Comparison between cultivation and sequencing based approaches for microbiota analysis in swabs and biopsies of chronic wounds. Front. Med. 2021, 8, 607255. [Google Scholar] [CrossRef]
- Zmudzińska, M.; Czarnecka-Operacz, M.; Silny, W. Bacterial flora of leg ulcers in patients admitted to Department of Dermatology, Poznań University of Medical Sciences, during the 1998–2002 period. Acta Dermatovenerol. Croat. 2005, 13, 168–172. [Google Scholar]
- Renner, R.; Sticherling, M.; Rüger, R.; Simon, J. Persistence of bacteria like Pseudomonas aeruginosa in non-healing venous ulcers. Eur. J. Dermatol. 2012, 22, 751–757. [Google Scholar] [CrossRef]
- Körber, A.; Schmid, E.; Buer, J.; Klode, J.; Schadendorf, D.; Dissemond, J. Bacterial colonization of chronic leg ulcers: Current results compared with data 5 years ago in a specialized dermatology department. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 1017–1025. [Google Scholar] [CrossRef]
- Al Ghazal, P.; Körber, A.; Klode, J.; Schmid, E.N.; Buer, J.; Dissemond, J. Evaluation of the essen rotary as a new technique for bacterial swabs: Results of a prospective controlled clinical investigation in 50 patients with chronic leg ulcers. Int. Wound J. 2014, 11, 44–49. [Google Scholar] [CrossRef]
- Jockenhöfer, F.; Gollnick, H.; Herberger, K.; Isbary, G.; Renner, R.; Stücker, M.; Valesky, E.; Wollina, U.; Weichenthal, M.; Karrer, S.; et al. Bacteriological pathogen spectrum of chronic leg ulcers: Results of a multicenter trial in dermatologic wound care centers differentiated by regions. JDDG J. Der Dtsch. Dermatol. Ges. 2013, 11, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Öien, R.F.; Åkesson, N. bacterial cultures, rapid strep test, and antibiotic treatment in infected hard-to-heal ulcers in primary care. Scand. J. Prim. Health Care 2012, 30, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.; Teixeira, S.; Fonseca, L.; Abreu, M.; Carvalho, A.; Pereira, M.T.; Amaral, C.; Freitas, C.; Ferreira, L.; Neto, H.R.; et al. Evolutionary trends in bacteria isolated from moderate and severe diabetic foot infections in a Portuguese tertiary center. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Kwiecińska-Piróg, J.; Przekwas, J.; Majkut, M.; Skowron, K.; Gospodarek-Komkowska, E. Biofilm formation reducing properties of manuka honey and propolis in Proteus mirabilis rods isolated from chronic wounds. Microorganisms 2020, 8, 1823. [Google Scholar] [CrossRef]
- Davies, C.E.; Hill, K.E.; Newcombe, R.G.; Stephens, P.; Wilson, M.J.; Harding, K.G.; Thomas, D.W. A prospective study of the microbiology of chronic venous leg ulcers to reevaluate the clinical predictive value of tissue biopsies and swabs. Wound Repair. Regen. 2007, 15, 17–22. [Google Scholar] [CrossRef]
- Nelson, A.; Wright-Hughes, A.; Backhouse, M.R.; Lipsky, B.A.; Nixon, J.; Bhogal, M.S.; Reynolds, C.; Brown, S. CODIFI (Concordance in Diabetic Foot Ulcer Infection): A cross-sectional study of wound swab versus tissue sampling in infected diabetic foot ulcers in England. BMJ Open 2018, 8, e019437. [Google Scholar] [CrossRef]
- Sloan, T.J.; Turton, J.C.; Tyson, J.; Musgrove, A.; Fleming, V.M.; Lister, M.M.; Loose, M.W.; Elizabeth Sockett, R.; Diggle, M.; Game, F.L.; et al. Examining diabetic heel ulcers through an ecological lens: Microbial community dynamics associated with healing and infection. J. Med. Microbiol. 2019, 68, 230–240. [Google Scholar] [CrossRef]
- Di Domenico, E.G.; Farulla, I.; Prignano, G.; Gallo, M.T.; Vespaziani, M.; Cavallo, I.; Sperduti, I.; Pontone, M.; Bordignon, V.; Cilli, L.; et al. Biofilm is a major virulence determinant in bacterial colonization of chronic skin ulcers independently from the multidrug resistant phenotype. Int. J. Mol. Sci. 2017, 18, 1077. [Google Scholar] [CrossRef]
- Di Domenico, E.G.; De Angelis, B.; Cavallo, I.; Sivori, F.; Orlandi, F.; D’autilio, M.F.L.M.; Di Segni, C.; Gentile, P.; Scioli, M.G.; Orlandi, A.; et al. Silver sulfadiazine eradicates antibiotic-tolerant Staphylococcus aureus and Pseudomonas aeruginosa biofilms in patients with infected diabetic foot ulcers. J. Clin. Med. 2020, 9, 3807. [Google Scholar] [CrossRef]
- Bessa, L.J.; Fazii, P.; Di Giulio, M.; Cellini, L. Bacterial isolates from infected wounds and their antibiotic susceptibility pattern: Some remarks about wound infection. Int. Wound J. 2015, 12, 47–52. [Google Scholar] [CrossRef]
- Thomsen, T.R.; Aasholm, M.S.; Rudkjøbing, V.B.; Saunders, A.M.; Bjarnsholt, T.; Givskov, M.; Kirketerp-Møller, K.; Nielsen, P.H. The bacteriology of chronic venous leg ulcer examined by culture-independent molecular methods. Wound Repair. Regen. 2010, 18, 38–49. [Google Scholar] [CrossRef]
- Gjødsbøl, K.; Skindersoe, M.E.; Christensen, J.J.; Karlsmark, T.; Jørgensen, B.; Jensen, A.M.; Klein, B.M.; Sonnested, M.K.; Krogfelt, K.A. No need for biopsies: Comparison of three sample techniques for wound microbiota determination. Int. Wound J. 2012, 9, 295–302. [Google Scholar] [CrossRef]
- Gjødsbøl, K.; Christensen, J.J.; Karlsmark, T.; Jørgensen, B.; Klein, B.M.; Krogfelt, K.A. Multiple bacterial species reside in chronic wounds: A longitudinal study. Int. Wound J. 2006, 3, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Phan, S.; Feng, C.H.; Huang, R.; Lee, Z.X.; Moua, Y.; Phung, O.J.; Lenhard, J.R. Relative abundance and detection of Pseudomonas aeruginosa from chronic wound infections globally. Microorganisms 2023, 11, 1210. [Google Scholar] [CrossRef] [PubMed]
- Saltoglu, N.; Surme, S.; Ezirmik, E.; Kadanali, A.; Kurt, A.F.; Sahin Ozdemir, M.; Ak, O.; Altay, F.A.; Acar, A.; Cakar, Z.S.; et al. The effects of antimicrobial resistance and the compatibility of initial antibiotic treatment on clinical outcomes in patients with diabetic foot infection. Int. J. Low. Extrem. Wounds 2023, 22, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Saltoglu, N.; Yemisen, M.; Ergonul, O.; Kadanali, A.; Karagoz, G.; Batirel, A.; Ak, O.; Eraksoy, H.; Cagatay, A.; Vatan, A.; et al. Predictors for limb loss among patient with diabetic foot infections: An observational retrospective multicentric study in Turkey. Clin. Microbiol. Infect. 2015, 21, 659–664. [Google Scholar] [CrossRef]
- Saltoglu, N.; Ergonul, O.; Tulek, N.; Yemisen, M.; Kadanali, A.; Karagoz, G.; Batirel, A.; Ak, O.; Sonmezer, C.; Eraksoy, H.; et al. Influence of multidrug resistant organisms on the outcome of diabetic foot infection. Int. J. Infect. Dis. 2018, 70, 10–14. [Google Scholar] [CrossRef]
- McManus, A.T.; Mason, A.D.; McManus, W.F.; Pruitt, B.A. Twenty-five year review of Pseudomonas aeruginosa bacteremia in a burn center. Eur. J. Clin. Microbiol. 1985, 4, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Gang, R.K.; Bang, R.L.; Sanyal, S.C.; Mokaddas, E.; Lari, A.R. Pseudomonas aeruginosa septicaemia in burns. Burns 1999, 25, 611–616. [Google Scholar] [CrossRef]
- Breidenstein, E.B.M.; de la Fuente-Núñez, C.; Hancock, R.E.W. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- López-Causapé, C.; Cabot, G.; del Barrio-Tofiño, E.; Oliver, A. The versatile mutational resistome of Pseudomonas aeruginosa. Front. Microbiol. 2018, 9, 685. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Mills, S.; Ross, R.P. Phages & antibiotic resistance: Are the most abundant entities on earth ready for a comeback? Future Microbiol. 2018, 13, 711–726. [Google Scholar]
- Kanj, S.S.; Kanafani, Z.A. current concepts in antimicrobial therapy against resistant Gram-negative organisms: Extended-spectrum beta-lactamase-producing Enterobacteriaceae, carbapenem-resistant Enterobacteriaceae, and multidrug-resistant Pseudomonas aeruginosa. Mayo Clin. Proc. 2011, 86, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Hraiech, S.; Brégeon, F.; Rolain, J. Bacteriophage-based therapy in cystic infections: Rationale and current status. Drug Des. Devel Ther. 2015, 9, 3653–3663. [Google Scholar]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 284, 1318–1322. [Google Scholar] [CrossRef]
- Lewis, K. Riddle of biofilm resistance. Antimicrob. Agents Chemother. 2001, 45, 999–1007. [Google Scholar] [CrossRef]
- Fong, S.A.; Drilling, A.J.; Ooi, M.L.; Paramasivan, S.; Finnie, J.W.; Morales, S.; Psaltis, A.J.; Vreugde, S.; Wormald, P.J. Safety and efficacy of a bacteriophage cocktail in an in vivo model of Pseudomonas aeruginosa sinusitis. Transl. Res. 2019, 206, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Engeman, E.; Freyberger, H.R.; Corey, B.W.; Ward, A.M.; He, Y.; Nikolich, M.P.; Filippov, A.A.; Tyner, S.D.; Jacobs, A.C. Synergistic killing and re-sensitization of Pseudomonas aeruginosa to antibiotics by phage-antibiotic combination treatment. Pharmaceuticals 2021, 14, 184. [Google Scholar] [CrossRef]
- Xuan, G.; Lin, H.; Kong, J.; Wang, J. Phage resistance evolution induces the sensitivity of specific antibiotics in Pseudomonas aeruginosa PAO1. Microbiol. Spectr. 2022, 10, e0135622. [Google Scholar] [CrossRef]
- Damir, G.; Knezevic, P. Filamentous Pseudomonas phage Pf4 in the context of therapy-inducibility, infectivity, lysogenic conversion, and potential application. Viruses 2022, 14, 1261. [Google Scholar] [CrossRef]
- Holger, D.J.; Lev, K.L.; Kebriaei, R.; Morrisette, T.; Shah, R.; Alexander, J.; Lehman, S.M.; Rybak, M.J. Bacteriophage-Antibiotic Combination therapy for multidrug-resistant Pseudomonas aeruginosa: In vitro synergy testing. J. Appl. Microbiol. 2022, 133, 1636–1649. [Google Scholar] [CrossRef] [PubMed]
- Wannasrichan, W.; Htoo, H.H.; Suwansaeng, R.; Pogliano, J.; Nonejuie, P.; Chaikeeratisak, V. Phage-resistant Pseudomonas aeruginosa against a novel lytic phage JJ01 exhibits hypersensitivity to colistin and reduces biofilm production. Front. Microbiol. 2022, 13, 1004733. [Google Scholar] [CrossRef]
- Valappil, S.K.; Shetty, P.; Deim, Z.; Terhes, G.; Urbán, E.; Váczi, S.; Patai, R.; Polgár, T.; Pertics, Z.B.; Schneider, G.; et al. Survival comes at a cost: A coevolution of phage and its host leads to phage resistance and antibiotic sensitivity of Pseudomonas aeruginosa multidrug resistant strains. Front. Microbiol. 2021, 12, 783722. [Google Scholar] [CrossRef]
- Aghaee, B.L.; Mirzaei, M.K.; Alikhani, M.Y.; Mojtahedi, A.; Maurice, C.F. Improving the inhibitory effect of phages against Pseudomonas aeruginosa isolated from a burn patient using a combination of phages and antibiotics. Viruses 2021, 13, 334. [Google Scholar] [CrossRef]
- Gurney, J.; Pradier, L.; Griffin, J.S.; Gougat-Barbera, C.; Chan, B.K.; Turner, P.E.; Kaltz, O.; Hochberg, M.E. Phage steering of antibiotic-resistance evolution in the bacterial pathogen, Pseudomonas aeruginosa. Evol. Med. Public. Health 2020, 2020, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, J.; Shigehisa, R.; Nasukawa, T.; Mizukami, K.; Takemura-Uchiyama, I.; Ujihara, T.; Murakami, H.; Imanishi, I.; Nishifuji, K.; Sakaguchi, M.; et al. Piperacillin and ceftazidime produce the strongest synergistic phage–antibiotic effect in Pseudomonas aeruginosa. Arch. Virol. 2018, 163, 1941–1948. [Google Scholar] [CrossRef]
- Van Nieuwenhuyse, B.; Van der Linden, D.; Chatzis, O.; Lood, C.; Wagemans, J.; Lavigne, R.; Schroven, K.; Paeshuyse, J.; de Magnée, C.; Sokal, E.; et al. Bacteriophage-antibiotic combination therapy against extensively drug-resistant Pseudomonas aeruginosa infection to allow liver transplantation in a toddler. Nat. Commun. 2022, 13, 5725. [Google Scholar] [CrossRef]
- Fiscarelli, E.V.; Rossitto, M.; Rosati, P.; Essa, N.; Crocetta, V.; Di Giulio, A.; Lupetti, V.; Di Bonaventura, G.; Pompilio, A. In vitro newly isolated environmental phage activity against biofilms preformed by Pseudomonas aeruginosa from patients with cystic fibrosis. Microorganisms 2021, 9, 478. [Google Scholar] [CrossRef]
- Racenis, K.; Rezevska, D.; Madelane, M.; Lavrinovics, E.; Djebara, S.; Petersons, A.; Kroica, J. Use of phage cocktail BFC 1.10 in combination with ceftazidime-avibactam in the treatment of multidrug-resistant Pseudomonas aeruginosa femur osteomyelitis—A case report. Front. Med. 2022, 9, 851310. [Google Scholar] [CrossRef]
- Broncano-Lavado, A.; Santamaría-Corral, G.; Esteban, J.; García-Quintanilla, M. Advances in bacteriophage therapy against relevant multidrug-resistant pathogens. Antibiotics 2021, 10, 672. [Google Scholar] [CrossRef]
- Al-Anany, A.M.; Fatima, R.; Hynes, A.P. Temperate phage-antibiotic synergy eradicates bacteria through depletion of lysogens. Cell Rep. 2021, 35, 109172. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.G.; Green, S.I.; Min, L.; Clark, J.R.; Salazar, K.C.; Terwilliger, A.L.; Kaplan, H.B.; Trautner, B.W.; Ramig, R.F.; Maresso, A.W. Phage-antibiotic synergy is driven by a unique combination of antibacterial mechanism of action and stoichiometry. mBio 2020, 11, e01462-20. [Google Scholar] [CrossRef] [PubMed]
- Santamaría-Corral, G.; Pagán, I.; Aguilera-Correa, J.J.; Esteban, J.; García-Quintanilla, M. A novel bacteriophage infecting multi-drug- and extended-drug-resistant Pseudomonas aeruginosa strains. Antibiotics 2024, 13, 523. [Google Scholar] [CrossRef] [PubMed]
- Akturk, E.; Oliveira, H.; Santos, S.B.; Costa, S.; Kuyumcu, S.; Melo, L.D.R.; Azeredo, J. Synergistic action of phage and antibiotics: Parameters to enhance the killing efficacy against mono and dual-species biofilms. Antibiotics 2019, 8, 103. [Google Scholar] [CrossRef]
- De Soir, S.; Parée, H.; Kamarudin, N.H.N.; Wagemans, J.; Lavigne, R.; Braem, A.; Merabishvili, M.; De Vos, D.; Pirnay, J.-P.; Van Bambeke, F. Exploiting phage-antibiotic synergies to disrupt Pseudomonas aeruginosa PAO1 biofilms in the context of orthopedic infections. Microbiol. Spectr. 2023, 12, e0321923. [Google Scholar] [CrossRef]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Salerian, A.J. Burn wound infections and Pseudomonas aeruginosa. Burns 2020, 46, 257–258. [Google Scholar] [CrossRef]
- Sivanmaliappan, T.S.; Sevanan, M. Antimicrobial susceptibility patterns of Pseudomonas aeruginosa from diabetes patients with foot ulcers. Int. J. Microbiol. 2011, 2011, 605195. [Google Scholar] [CrossRef] [PubMed]
- Rahim, K.; Saleha, S.; Zhu, X.; Huo, L.; Basit, A.; Franco, O.L. Bacterial contribution in chronicity of wounds. Microb. Ecol. 2017, 73, 710–721. [Google Scholar] [CrossRef]
- Kirketerp-Møller, K.; Jensen, P.; Fazli, M.; Madsen, K.G.; Pedersen, J.; Moser, C.; Tolker-Nielsen, T.; Høiby, N.; Givskov, M.; Bjarnsholt, T. Distribution, organization, and ecology of bacteria in chronic wounds. J. Clin. Microbiol. 2008, 46, 2717–2722. [Google Scholar] [CrossRef]
- Sun, Y.; Dowd, S.E.; Smith, E.; Rhoads, D.D.; Wolcott, R.D. In vitro multispecies lubbock chronic wound biofilm model. Wound Repair. Regen. 2008, 16, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Akturk, E.; Melo, L.D.R.; Oliveira, H.; Crabbé, A.; Coenye, T.; Azeredo, J. Combining phages and antibiotic to enhance antibiofilm efficacy against an in vitro dual species wound biofilm. Biofilm 2023, 6, 100147. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, P.; Clark, J.R.; Liu, C.G.; Terwilliger, A.; Maresso, A.W. Class-driven synergy and antagonism between a pseudomonas phage and antibiotics. Infect. Immun. 2023, 91, e0006523. [Google Scholar] [CrossRef]
- Łusiak-Szelachowska, M.; Międzybrodzki, R.; Drulis-Kawa, Z.; Cater, K.; Knežević, P.; Winogradow, C.; Amaro, K.; Jończyk-Matysiak, E.; Weber-Dąbrowska, B.; Rękas, J.; et al. Bacteriophages and antibiotic interactions in clinical practice: What we have learned so far. J. Biomed. Sci. 2022, 29, 23. [Google Scholar] [CrossRef]
- Shariati, A.; Noei, M.; Chegini, Z. Bacteriophages: The promising therapeutic approach for enhancing ciprofloxacin efficacy against bacterial infection. J. Clin. Lab. Anal. 2023, 37, e24932. [Google Scholar] [CrossRef]
- Martin, I.; Morales, S.; Alton, E.W.F.W.; Davies, J.C. Lytic bacteriophage is a promising adjunct to common antibiotics across cystic fibrosis clinical strains and culture models of Pseudomonas aeruginosa infection. Antibiotics 2023, 12, 593. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, K.; Rørbo, N.; Rybtke, M.L.; Martinet, M.G.; Tolker-Nielsen, T.; Høiby, N.; Middelboe, M.; Ciofu, O.P. aeruginosa flow-cell biofilms are enhanced by repeated phage treatments but can be eradicated by phage-ciprofloxacin combination. Pathog. Dis. 2019, 77, ftz011. [Google Scholar] [CrossRef] [PubMed]
- Yoon, R.; Chang, K.; Das, T.; Manos, J.; Kutter, E.; Morales, S.; Chan, H.-K. Bacteriophage PEV20 and ciprofloxacin combination treatment enhances removal of P. aeruginosa biofilm isolated from cystic fibrosis and wound patients. HHS Public Access. AAPS J. 2020, 21, 49. [Google Scholar] [CrossRef]
- Drilling, A.; Morales, S.; Jardeleza, C.; Vreugde, S.; Speck, P.; Wormald, P.J. Bacteriophage reduces biofilm of Staphylococcus aureus ex vivo isolates from chronic rhinosinusitis patients. Am. J. Rhinol. Allergy 2014, 28, 3–11. [Google Scholar] [CrossRef]
- Clokie, M.R.J.; Kropinski, A.M.; Lavigne, R. (Eds.) Bacteriophages: Methods and Protocols, Volume 3; Humana: New York, NY, USA, 2017; ISBN 978-1-4939-7343-9. [Google Scholar]
- Nikolic, I.; Vukovic, D.; Gavric, D.; Cvetanovic, J.; Aleksic Sabo, V.; Gostimirovic, S.; Narancic, J.; Knezevic, P. An optimized checkerboard method for phage-antibiotic synergy detection. Viruses 2022, 14, 1542. [Google Scholar] [CrossRef]
- Oesterreicher, Z.; Minichmayr, I.; Sauermann, R.; Marhofer, D.; Lackner, E.; Jäger, W.; Maier-Salamon, A.; Schwameis, R.; Kloft, C.; Zeitlinger, M. Pharmacokinetics of doripenem in plasma and epithelial lining fluid (elf): Comparison of two dosage regimens. Eur. J. Clin. Pharmacol. 2017, 73, 1609–1613. [Google Scholar] [CrossRef] [PubMed]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary biofilm device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Merck & Co., Inc. Primaxin ® I.V (Imipenem and Cilastatin for Injection). Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2009/050587s072,050630s035lbl.pdf (accessed on 26 May 2024).
- Uricová, J.; Kacířová, I.; Brozmanová, H. Meropenem serum concentrations in intensive care patients: A retrospective analysis. Ceska Slov. Farm. 2020, 69, 230–236. [Google Scholar] [PubMed]
- Alvarez-Lerma, F.; Grau, S.; Ferrández, O. Characteristics of doripenem: A new broad-spectrum antibiotic. Drug Des. Devel Ther. 2009, 3, 173. [Google Scholar] [CrossRef]
- Varghese, J.M.; Jarrett, P.; Boots, R.J.; Kirkpatrick, C.M.J.; Lipman, J.; Roberts, J.A. Pharmacokinetics of piperacillin and tazobactam in plasma and subcutaneous interstitial fluid in critically ill patients receiving continuous venovenous haemodiafiltration. Int. J. Antimicrob. Agents 2014, 43, 343–348. [Google Scholar] [CrossRef]
- Ramsey, C.; MacGowan, A.P. A Review of the pharmacokinetics and pharmacodynamics of aztreonam. J. Antimicrob. Chemother. 2016, 71, 2704–2712. [Google Scholar] [CrossRef]
- Schober, P.; Schwarte, L.A. Correlation coefficients: Appropriate use and interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAO1 | PA24 | PA35 | PA36 | |
|---|---|---|---|---|
| Aztreonam | ≤0.60 | 0.23 | 0.03 | ≤0.13 |
| Doripenem | ≤0.60 | 0.10 | 1.00 | ≤0.12 |
| Imipenem | ≤2.00 | 0.10 | 1.01 | ≤0.10 |
| Meropenem | ≤2.00 | 0.10 | 0.10 | ≤0.12 |
| Piperacillin/tazobactam | ≤0.60 | 0.13 | 1.01 | ≤0.13 |
| PA24 | PA35 | PA36 | |
|---|---|---|---|
| Source | Sputum | - | Sputum |
| MDR/XDR | MDR | XDR | MDR |
| AMK | S | S | S |
| AZ | S | R | S |
| C/T | S | R | S |
| CEF | S | R | S |
| CFT | S | R | R |
| CIP | R | R | R |
| CO | S | S | S |
| DOR | R | R | R |
| GE | R | R | R |
| IM | R | R | R |
| MP | S | R | S |
| P/T | S | R | R |
| TOB | R | R | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santamaría-Corral, G.; Aguilera-Correa, J.J.; Esteban, J.; García-Quintanilla, M. Bacteriophage Therapy on an In Vitro Wound Model and Synergistic Effects in Combination with Beta-Lactam Antibiotics. Antibiotics 2024, 13, 800. https://doi.org/10.3390/antibiotics13090800
Santamaría-Corral G, Aguilera-Correa JJ, Esteban J, García-Quintanilla M. Bacteriophage Therapy on an In Vitro Wound Model and Synergistic Effects in Combination with Beta-Lactam Antibiotics. Antibiotics. 2024; 13(9):800. https://doi.org/10.3390/antibiotics13090800
Chicago/Turabian StyleSantamaría-Corral, Guillermo, John Jairo Aguilera-Correa, Jaime Esteban, and Meritxell García-Quintanilla. 2024. "Bacteriophage Therapy on an In Vitro Wound Model and Synergistic Effects in Combination with Beta-Lactam Antibiotics" Antibiotics 13, no. 9: 800. https://doi.org/10.3390/antibiotics13090800