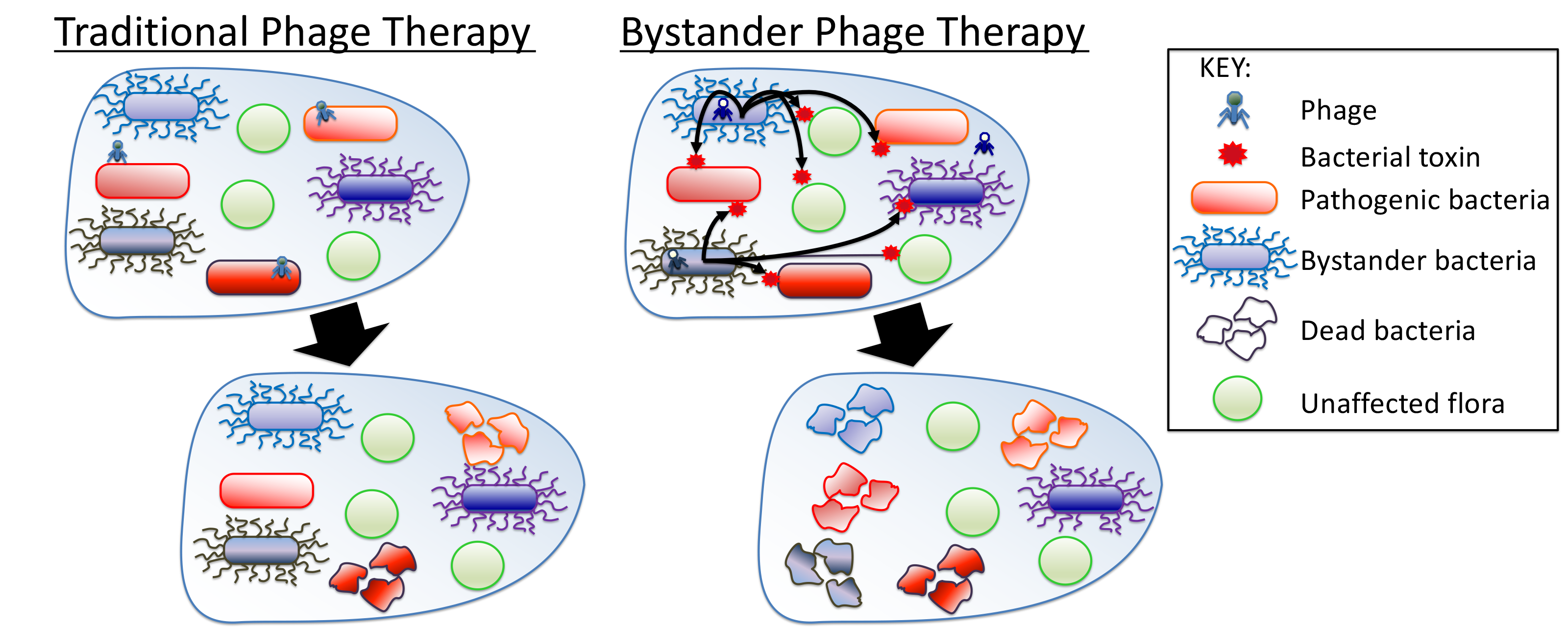
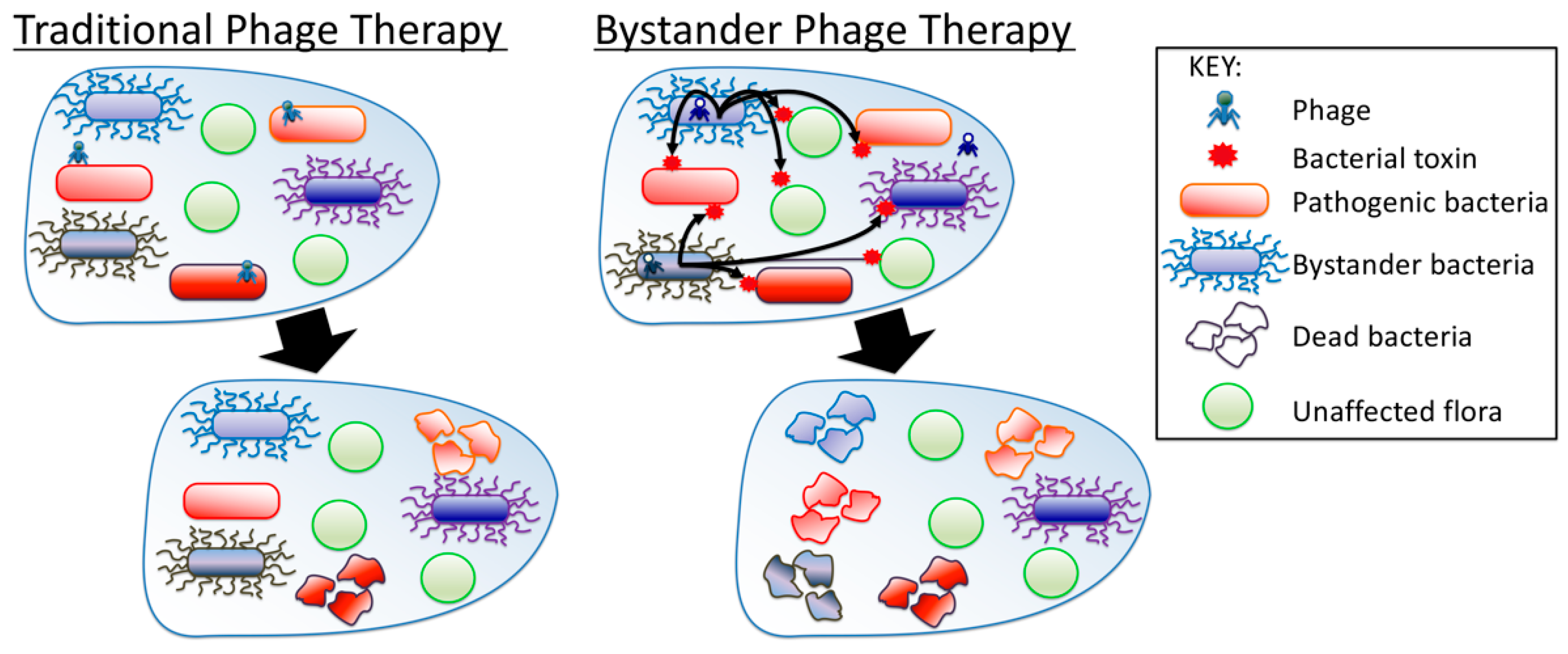
Bystander Phage Therapy: Inducing Host-Associated Bacteria to Produce Antimicrobial Toxins against the Pathogen Using Phages

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Phage Characteristics and Host Range
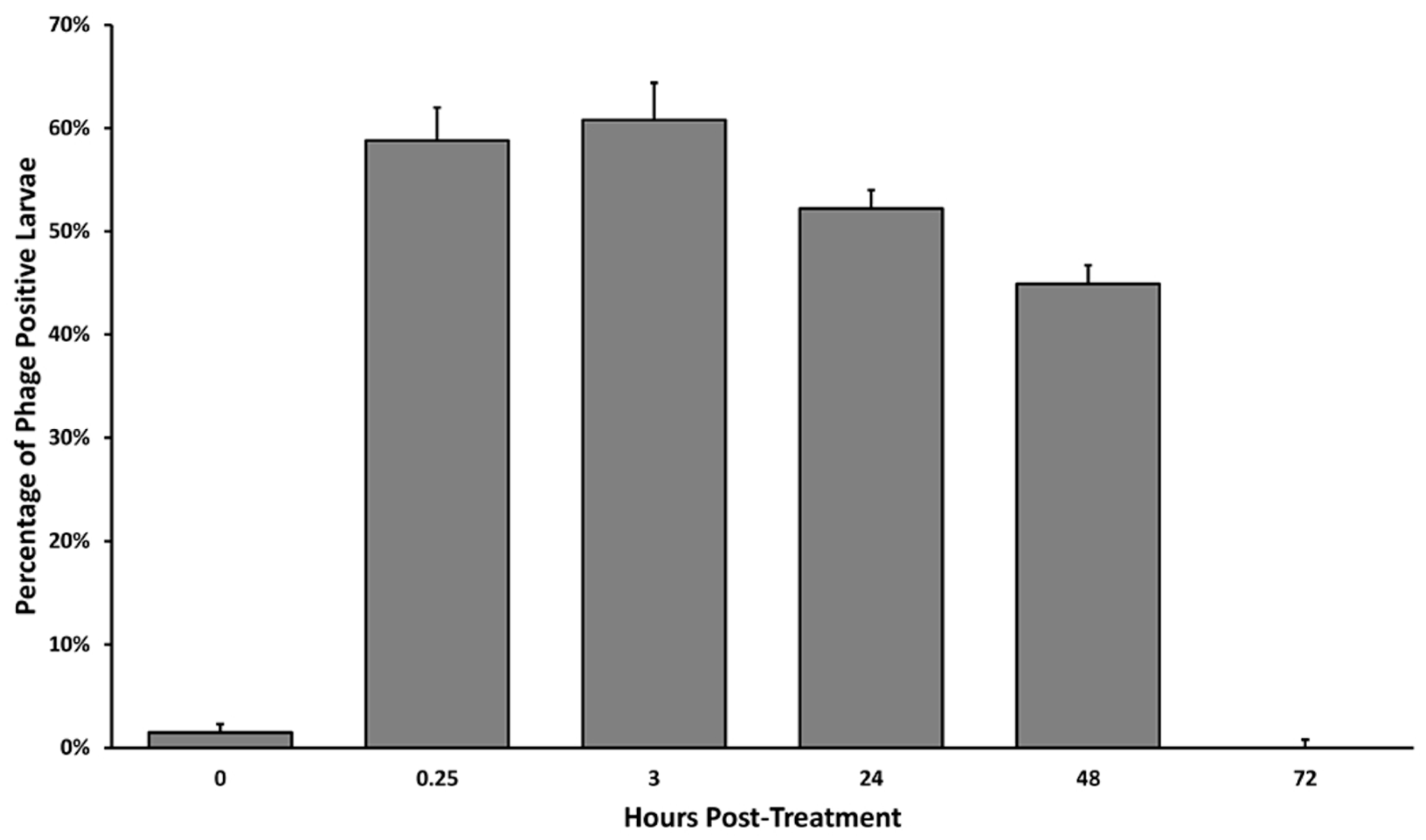
2.2. Phage Persistence in the Larval Honeybee
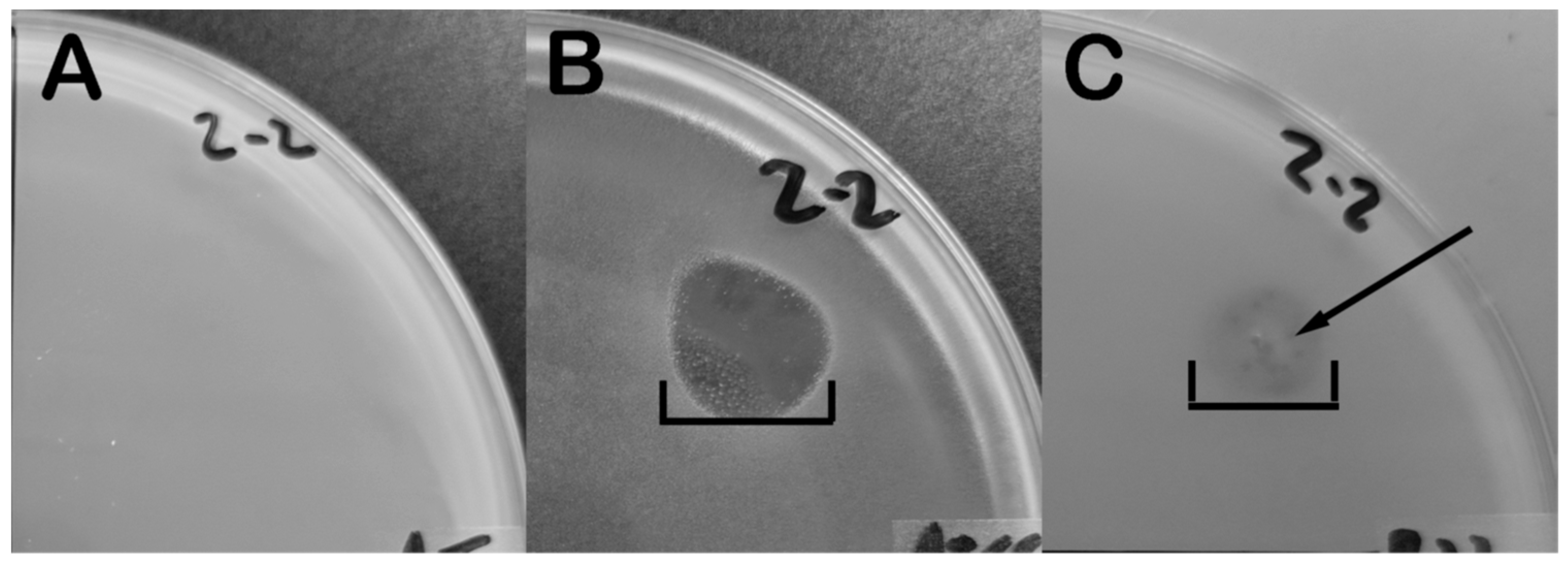
2.3. Phage Infection Induces B. laterosporus to Produce Antimicrobials
2.4. Phage-Induced B. laterosporus Antimicrobial Products Shows Inert Characteristics Against Honeybees
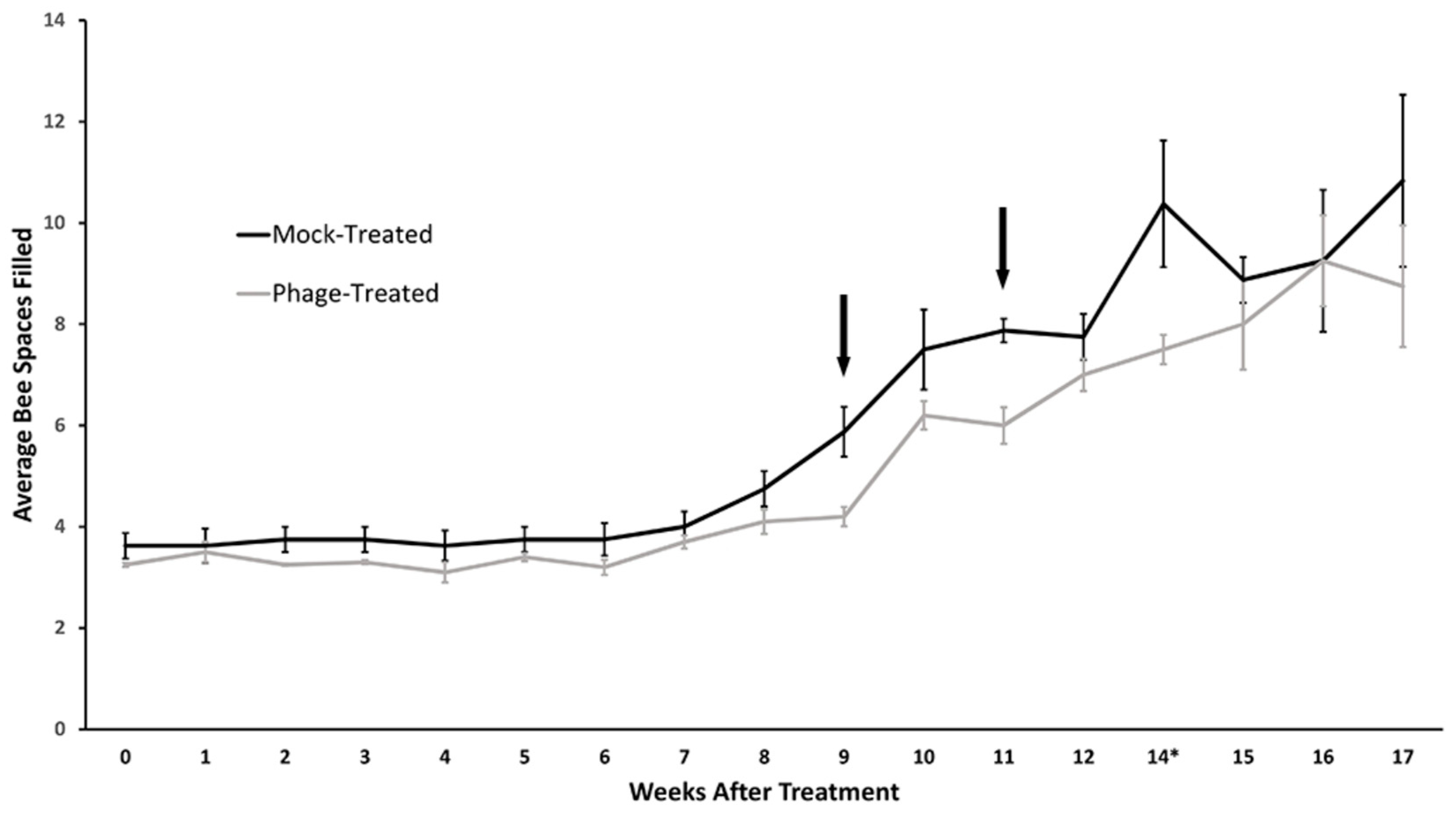
2.5. B. laterosporus Phages Can Effectively Treat an Active AFB Infection
2.6. B. Laterosporus Phages Do not Prevent Reinfection by Latent P. larvae Spores
3. Discussion
4. Materials and Methods
4.1. Gathering B. laterosporus Field Isolates
4.2. Isolating Phages Specific for B. laterosporus
4.3. Host Range and Phage Presence Testing for Isolated Phages
4.4. Electron Microscopy
4.5. Creation of Bacterial Lysate to Test for B. Laterosporus and Phage Cocktail Treatments
4.6. Phage Beehive Parameters
4.7. Statistics
5. Conclusions
6. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roman-Blanco, C.; Sanz-Gomez, J.J.; Lopez-Diaz, T.M.; Otero, A.; Garcia-Lopez, M.L. Numbers and species of Bacillus during the manufacture and ripening of Castellano cheese. Milchwissenschaft-Milk Sci. Int. 1999, 54, 385–388. [Google Scholar]
- Khan, M.R.; Saha, M.L.; Afroz, H. Microorganisms associated with gemstones. Bangladesh J. Bot. 2001, 30, 93–96. [Google Scholar]
- De Oliveira, E.J.; Rabinovitch, L.; Monnerat, R.G.; Passos, L.K.J.; Zahner, V. Molecular characterization of Brevibacillus laterosporus and its potential use in biological control. Appl. Environ. Microbiol. 2004, 70, 6657–6664. [Google Scholar] [CrossRef] [PubMed]
- Suslova, M.Y.; Lipko, I.A.; Mamaeva, E.V.; Parfenova, V.V. Diversity of cultivable bacteria isolated from the water column and bottom sediments of the Kara Sea shelf. Microbiology 2012, 81, 484–491. [Google Scholar] [CrossRef]
- Ruiu, L. Brevibacillus laterosporus, a Pathogen of Invertebrates and a Broad-Spectrum Antimicrobial Species. Insects 2013, 4, 476–492. [Google Scholar] [CrossRef]
- Alippi, A.M.; Lopez, A.C.; Aguilar, O.M. Differentiation of Paenibacillus larvae subsp larvae, the cause of American foulbrood of honeybees, by using PCR and restriction fragment analysis of genes encoding 16S rRNA. Appl. Environ. Microbiol. 2002, 68, 3655–3660. [Google Scholar] [CrossRef]
- Genersch, E. American Foulbrood in honeybees and its causative agent, Paenibacillus larvae. J. Invertebr. Pathol. 2010, 103, S10–S19. [Google Scholar] [CrossRef]
- Pohorecka, K.; Skubida, M.; Bober, A.; Zdanska, D. Screening of Paenibacillus Larvae Spores in Apiaries from Eastern Poland. Nationwide Survey. Part I. Bull. Vet. Inst. Pulawy 2012, 56, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Ebeling, J.; Knispel, H.; Hertlein, G.; Funfhaus, A.; Genersch, E. Biology of Paenibacillus larvae, a deadly pathogen of honey bee larvae. Appl. Microbiol. Biotechnol. 2016, 100, 7387–7395. [Google Scholar] [CrossRef]
- Yang, X.; Huang, E.; Yuan, C.H.; Zhang, L.W.; Yousef, A.E. Isolation and Structural Elucidation of Brevibacillin, an Antimicrobial Lipopeptide from Brevibacillus laterosporus That Combats Drug-Resistant Gram-Positive Bacteria. Appl. Environ. Microbiol. 2016, 82, 2763–2772. [Google Scholar] [CrossRef]
- Khaled, J.M.; Al-Mekhlafi, F.A.; Mothana, R.A.; Alharbi, N.S.; Alzaharni, K.E.; Sharafaddin, A.H.; Kadaikunnan, S.; Alobaidi, A.S.; Bayaqoob, N.I.; Govindarajan, M.; et al. Brevibacillus laterosporus isolated from the digestive tract of honeybees has high antimicrobial activity and promotes growth and productivity of honeybee’s colonies. Environ. Sci. Pollut. Res. Int. 2017, 25, 10447–10455. [Google Scholar] [CrossRef] [PubMed]
- Alippi, A.M.; Reynaldi, F.J. Inhibition of the growth of Paenibacillus larvae, the causal agent of American foulbrood of honeybees, by selected strains of aerobic spore-forming bacteria isolated from apiarian sources. J. Invertebr. Pathol. 2006, 91, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Saikia, R.; Gogoi, D.K.; Mazumder, S.; Yadav, A.; Sarma, R.K.; Bora, T.C.; Gogoi, B.K. Brevibacillus laterosporus strain BPM3, a potential biocontrol agent isolated from a natural hot water spring of Assam, India. Microbiol. Res. 2011, 166, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Marche, M.G.; Mura, M.E.; Ruiu, L. Brevibacillus laterosporus inside the insect body: Beneficial resident or pathogenic outsider? J. Invertebr. Pathol. 2016, 137, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.F.; Nielsen-LeRoux, C. Mosquitocidal bacterial toxins: Diversity, mode of action and resistance phenomena. Mem. Inst. Oswaldo Cruz 2000, 95, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Ruiu, L.; Satta, A.; Floris, I. Observations on house fly larvae midgut ultrastructure after Brevibacillus laterosporus ingestion. J. Invertebr. Pathol. 2012, 111, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Bashir, F.; Aslam, S.; Khan, R.A.; Shahzadi, R. Larvicidal Activity of Bacillus laterosporus Against Mosquitoes. Pak. J. Zool. 2016, 48, 281–284. [Google Scholar]
- Mura, M.E.; Ruiu, L. Brevibacillus laterosporus pathogenesis and local immune response regulation in the house fly midgut. J. Invertebr. Pathol. 2017, 145, 55–61. [Google Scholar] [CrossRef]
- Merrill, B.D.; Grose, J.H.; Breakwell, D.P.; Burnett, S.H. Characterization of Paenibacillus larvae bacteriophages and their genomic relationships to firmicute bacteriophages. BMC Genom. 2014, 15, 745. [Google Scholar] [CrossRef]
- Merrill, B.D.; Berg, J.A.; Graves, K.A.; Ward, A.T.; Hilton, J.A.; Wake, B.N.; Grose, J.H.; Breakwell, D.P.; Burnett, S.H. Genome Sequences of Five Additional Brevibacillus laterosporus Bacteriophages. Genome Announc. 2015, 3, e01146-15. [Google Scholar] [CrossRef]
- Berg, J.A.; Merrill, B.D.; Crockett, J.T.; Esplin, K.P.; Evans, M.R.; Heaton, K.E.; Hilton, J.A.; Hyde, J.R.; McBride, M.S.; Schouten, J.T.; et al. Characterization of Five Novel Brevibacillus Bacteriophages and Genomic Comparison of Brevibacillus Phages. PLoS ONE 2016, 11, e0156838. [Google Scholar] [CrossRef]
- Stamereilers, C.; Fajardo, C.P.; Walker, J.K.; Mendez, K.N.; Castro-Nallar, E.; Grose, J.H.; Hope, S.; Tsourkas, P.K. Genomic Analysis of 48 Paenibacillus larvae Bacteriophages. Viruses 2018, 10, 377. [Google Scholar] [CrossRef] [PubMed]
- Brady, T.S.; Merrill, B.D.; Hilton, J.A.; Payne, A.M.; Stephenson, M.B.; Hope, S. Bacteriophages as an alternative to conventional antibiotic use for the prevention or treatment of Paenibacillus larvae in honeybee hives. J. Invertebr. Pathol. 2017, 150, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Pereira, C.; Santos, L.; Klumpp, J.; Almeida, A. Potential of phage cocktails in the inactivation of Enterobacter cloacae–An in vitro study in a buffer solution and in urine samples. Virus Res. 2016, 211, 199–208. [Google Scholar] [CrossRef]
- Mateus, L.; Costa, L.; Silva, Y.J.; Pereira, C.; Cunha, A.; Almeida, A. Efficiency of phage cocktails in the inactivation of Vibrio in aquaculture. Aquaculture 2014, 424, 167–173. [Google Scholar] [CrossRef]
- Yost, D.G.; Tsourkas, P.; Amy, P.S. Experimental bacteriophage treatment of honeybees (Apis mellifera) infected with Paenibacillus larvae, the causative agent of American Foulbrood Disease. Bacteriophage 2016, 6, e1122698. [Google Scholar] [CrossRef] [PubMed]
- Bruttin, A.; Brussow, H. Human volunteers receiving Escherichia coli phage T4 orally: A safety test of phage therapy. Antimicrob. Agents Chemother. 2005, 49, 2874–2878. [Google Scholar] [CrossRef]
- Endersen, L.; Buttimer, C.; Nevin, E.; Coffey, A.; Neve, H.; Oliveira, H.; Lavigne, R.; O’Mahony, J. Investigating the biocontrol and anti-biofilm potential of a three phage cocktail against Cronobacter sakazakii in different brands of infant formula. Int. J. Food Microbiol. 2017, 253, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, M.K.; Nilsson, A.S. Isolation of Phages for Phage Therapy: A Comparison of Spot Tests and Efficiency of Plating Analyses for Determination of Host Range and Efficacy. PLoS ONE 2015, 10, 13. [Google Scholar] [CrossRef]
- Regeimbal, J.M.; Jacobs, A.C.; Corey, B.W.; Henry, M.S.; Thompson, M.G.; Pavlicek, R.L.; Quinones, J.; Hannah, R.M.; Ghebremedhin, M.; Crane, N.J.; et al. Personalized Therapeutic Cocktail of Wild Environmental Phages Rescues Mice from Acinetobacter baumannii Wound Infections. Antimicrob. Agents Chemother. 2016, 60, 5806–5816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutter, E.; De Vos, D.; Gvasalia, G.; Alavidze, Z.; Gogokhia, L.; Kuhl, S.; Abedon, S.T. Phage Therapy in Clinical Practice: Treatment of Human Infections. Curr. Pharm. Biotechnol. 2010, 11, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Ruiu, L.; Satta, A.; Floris, I. Emerging entomopathogenic bacteria for insect pest management. Bull. Insectol. 2013, 66, 181–186. [Google Scholar]
- Furfaro, L.L.; Payne, M.S.; Chang, B.J. Bacteriophage Therapy: Clinical Trials and Regulatory Hurdles. Front. Cell. Infect. Microbiol. 2018, 8, 376. [Google Scholar] [CrossRef] [PubMed]
- Forsgren, E.; Stevanovic, J.; Fries, I. Variability in germination and in temperature and storage resistance among Paenibacillus larvae genotypes. Vet. Microbiol. 2008, 129, 342–349. [Google Scholar] [CrossRef]
- De Graaf, D.C.; Alippi, A.M.; Antunez, K.; Aronstein, K.A.; Budge, G.; De Koker, D.; De Smet, L.; Dingman, D.W.; Evans, J.D.; Foster, L.J.; et al. Standard methods for American Foulbrood research. J. Apic. Res. 2013, 52, 1–28. [Google Scholar] [CrossRef]
- Genersch, E.; Forsgren, E.; Pentikainen, J.; Ashiralieva, A.; Rauch, S.; Kilwinski, J.; Fries, I. Reclassification of Paenibacillus larvae subsp. pulvifaciens and Paenibacillus larvae subsp. larvae as Paenibacillus larvae without subspecies differentiation. Int. J. Syst. Evol. Microbiol. 2006, 56, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.A.; Merrill, B.D.; Breakwell, D.P.; Hope, S.; Grose, J.H. A PCR-Based Method for Distinguishing between Two Common Beehive Bacteria, Paenibacillus larvae and Brevibacillus laterosporus. J. Appl. Environ. Microbiol. 2018, 84, e01886-18. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Alippi, A.M.; López, A.C.; Aguilar, O.M. A PCR-based method that permits specific detection of Paenibacillus larvae subsp. larvae, the cause of American Foulbrood of honey bees, at the subspecies level. Lett. Appl. Microbiol. 2004, 39, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, B.J.; Helgason, T.; Poppinga, L.; Funfhaus, A.; Genersch, E.; Budge, G.E. Biogeography of Paenibacillus larvae, the causative agent of American foulbrood, using a new multilocus sequence typing scheme. Environ. Microbiol. 2015, 17, 1414–1424. [Google Scholar] [CrossRef] [PubMed]
- Versalovic, J.; Schneider, M.; De Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage | BL2 | BL6 | BL14 | 40A1 | 40A2 | 40A3 | 40A4 | 40A5 | 40A6 | 40A8 | 40A9 | 40A10 | PL ATCC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jimmer1 | ++++ | − | ++++ | + | − | − | − | − | − | − | − | − | − |
| Jimmer2 | ++++ | − | ++++ | + | − | − | − | − | − | − | − | − | − |
| Osiris | ++++ | − | ++ | ++ | + | + | − | + | ++ | − | ++ | + | − |
| Fawkes | ++++ | − | ++ | +++ | + | − | − | − | + | − | ++++ | ++ | − |
| Lauren | ++++ | − | ++++ | + | − | − | − | − | + | − | + | + | − |
| Powder/ Sundance | +++ | − | +++ | +++ | − | + | − | − | − | − | +++ | − | − |
| SecTim467 | +++ | ++ | +++ | ++ | + | − | − | − | − | − | +++ | − | − |
| Jenst | − | ++++ | − | + | − | − | − | − | + | − | +++ | − | − |
| Davies | − | ++++ | − | ++++ | ++ | + | − | +++ | +++ | +++ | − | − | − |
| Emery/ Abouo | − | ++++ | − | ++++ | ++++ | + | − | +++ | +++ | ++ | +++ | − | − |
| Source Tested | B. Laterosporus (BL-2) | B. laterosporus (BL-6) | P. larvae | E. coli | A. tumefaciens | S. Meliloti |
|---|---|---|---|---|---|---|
| Emery/Abouo Phage lysate (BL-6) | +++ | ++ * | ++++ | + | − | − |
| Fawkes Phage lysate (BL-2) | ++ * | +++ | ++++ | + | − | − |
| Supernatant of live B. Laterosporus | − | − | − | − | − | − |
| Supernatant of UV killed B. Laterosporus | − | − | − | − | − | − |
| Supernatant of mechanically lysed B. Laterosporus | − | − | − | − | − | − |
| Hive Status | Total Hives | AFB-Free Post-Treatment | Hive Survival Over Winter |
|---|---|---|---|
| Uninfected hives | 28 | 92.85% * | 78.1% † |
| AFB infected hives | 12 | 75% | 62.5% |
| Hive Status | Week 16 | Week 18 * | Week 20 | Week 22 * | Week 24 | Week 26 * | Week 28 |
|---|---|---|---|---|---|---|---|
| Healthy | 5 | 4 | 5 | 3 | 5 | 0 | 5 |
| AFB+ | 0 | 1 | 0 | 2 | 0 | 5 | 0 |
| Primer | Sequence | Direction | Purpose | Reference |
|---|---|---|---|---|
| 27F | 5′-AGAGTTTGATCMTGGCTCAG-3′ | Forward | 16S rRNA universal primer | [38] |
| 907R | 5′-CCGTCAATTCMTTTRAGTTT-3′ | Reverse | ||
| BLrpoB-F | 5′-GCAGGTAAACTGGTCCAGAGCG-3′ | Forward | B. laterosporus rpoB | - |
| BLrpoB-R | 5′-CACCTGTTGATTTATCAATCAGCG-3′ | Reverse | ||
| KAT1 | 5′-ACAAACACTGGACCCGATCTAC-3′ | Forward | P. larvae ERIC-1 or ERIC-2 | [39] |
| KAT2 | 5′-CCGCCTTCTTCATATCTCCC-3′ | Reverse | ||
| PLrpoB-F | 5′-ATAACGCGAGACATTCCTAA-3′ | Forward | Amplifies P. larvae rpoB | [40] |
| PLrpoB-R | 5′-GAACGGCATATCTTCTTCAG-3′ | Reverse | ||
| PLftsA-F | 5′-AAATCGGTGAGGAAGACATT-3′ | Forward | Amplifies P. larvae ftsA | [40] |
| PLftsA-R | 5′-TGCCAATACGGTTTACTTTA-3′ | Reverse | ||
| ERIC1R | 5′-ATGTAAGCTCCTGGGGATTCAC-3′ | Forward | Generates multiple amplicons to fingerprint the bacteria tested | [41] |
| ERIC2 | 5′-AAGTAAGTGACTGGGGTGAGCG-3′ | Reverse |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brady, T.S.; Fajardo, C.P.; Merrill, B.D.; Hilton, J.A.; Graves, K.A.; Eggett, D.L.; Hope, S. Bystander Phage Therapy: Inducing Host-Associated Bacteria to Produce Antimicrobial Toxins against the Pathogen Using Phages. Antibiotics 2018, 7, 105. https://doi.org/10.3390/antibiotics7040105
Brady TS, Fajardo CP, Merrill BD, Hilton JA, Graves KA, Eggett DL, Hope S. Bystander Phage Therapy: Inducing Host-Associated Bacteria to Produce Antimicrobial Toxins against the Pathogen Using Phages. Antibiotics. 2018; 7(4):105. https://doi.org/10.3390/antibiotics7040105
Chicago/Turabian StyleBrady, T. Scott, Christopher P. Fajardo, Bryan D. Merrill, Jared A. Hilton, Kiel A. Graves, Dennis L. Eggett, and Sandra Hope. 2018. "Bystander Phage Therapy: Inducing Host-Associated Bacteria to Produce Antimicrobial Toxins against the Pathogen Using Phages" Antibiotics 7, no. 4: 105. https://doi.org/10.3390/antibiotics7040105
APA StyleBrady, T. S., Fajardo, C. P., Merrill, B. D., Hilton, J. A., Graves, K. A., Eggett, D. L., & Hope, S. (2018). Bystander Phage Therapy: Inducing Host-Associated Bacteria to Produce Antimicrobial Toxins against the Pathogen Using Phages. Antibiotics, 7(4), 105. https://doi.org/10.3390/antibiotics7040105