The Contribution of Efflux Pumps in Mycobacterium abscessus Complex Resistance to Clarithromycin

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Phenotypic and Genotypic Characterization of the M. abscessus Strains
2.2. Evaluation of the Synergistic Effect between Efflux Inhibitors and Clarithromycin
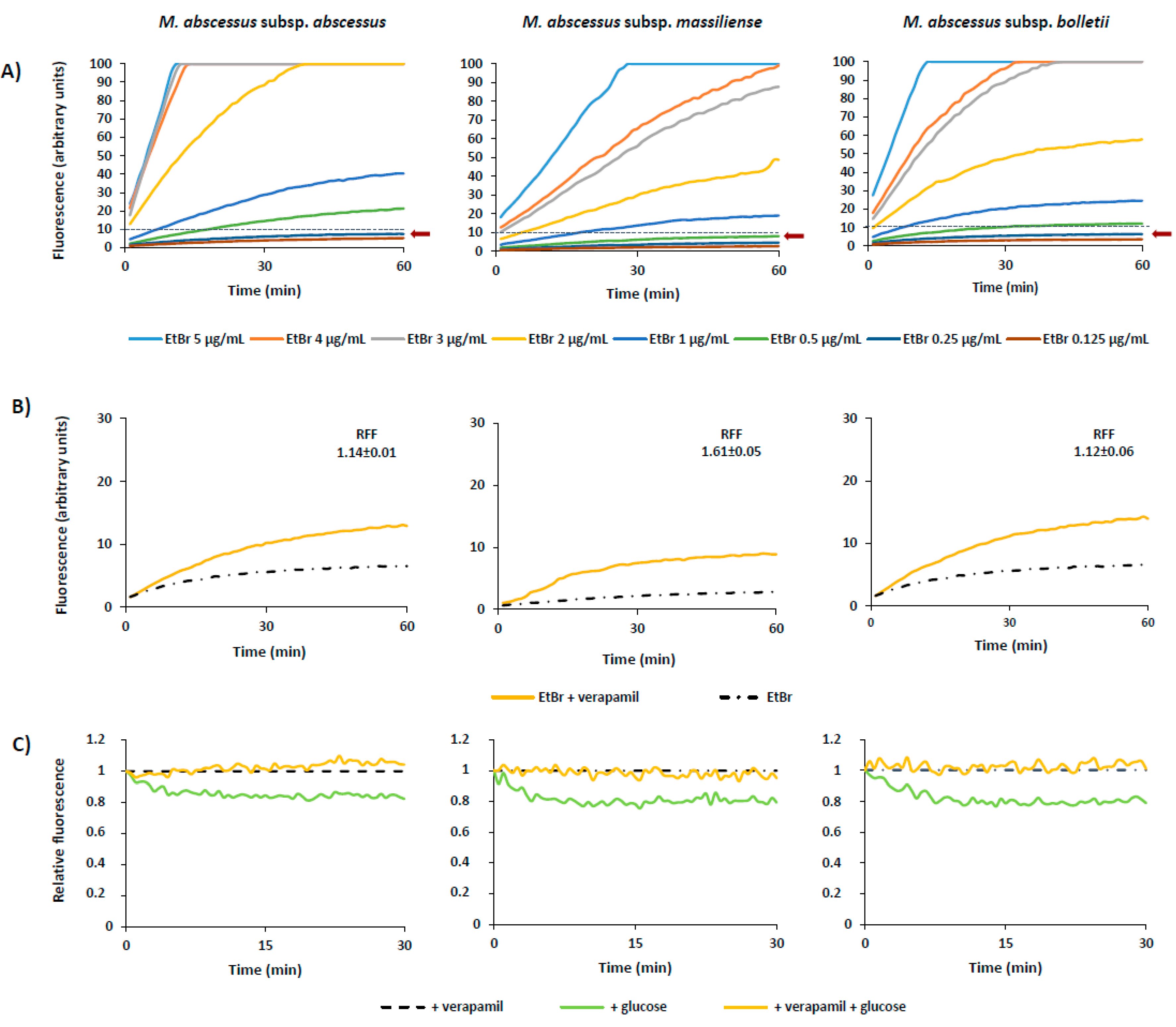
2.3. Assessment of Efflux Activity by Real-Time Fluorometry
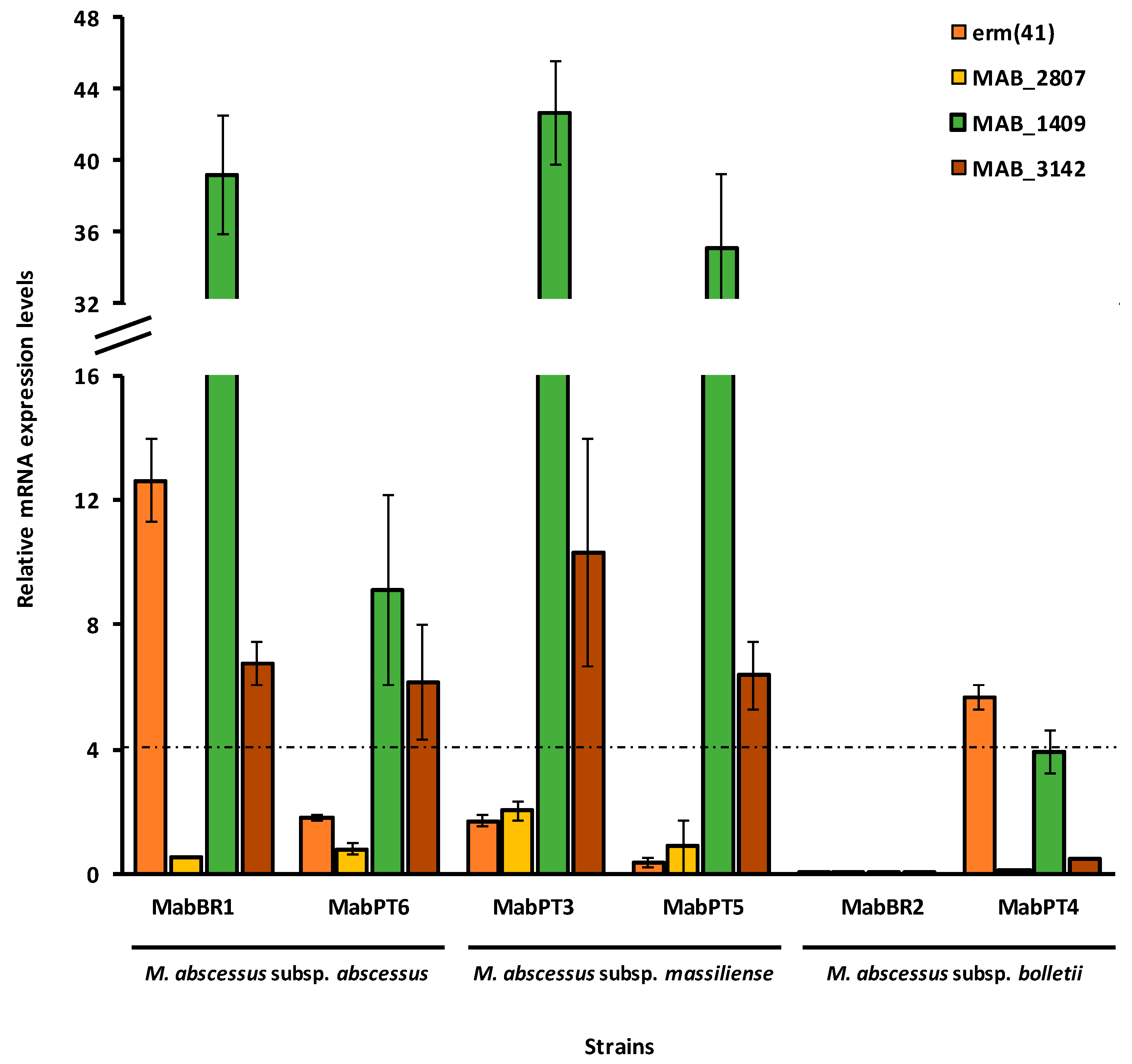
2.4. Quantification of Efflux Pump mRNA Levels by RT-qPCR
3. Discussion
4. Materials and Methods
4.1. M. abscessus Complex Strains
4.2. Compounds and Reagents
4.3. Detection of Mutations Associated with Clarithromycin and Amikacin Resistance
4.4. Susceptibility Testing
4.4.1. Growth of the Strains
4.4.2. MIC Determination of Compounds
4.4.3. MIC Determination of Antibiotics and Ethidium Bromide in the Presence of Verapamil
4.4.4. Semi-Automated Fluorometric Method
4.5. Efflux Pump Gene Expression
4.5.1. Sample Preparation
4.5.2. RNA Extraction
4.5.3. RT-qPCR Assay
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mougari, F.; Guglielmetti, L.; Raskine, L.; Sermet-Gaudelus, I.; Veziris, N.; Cambau, E. Infections caused by Mycobacterium abscessus: Epidemiology, diagnostic tools and treatment. Expert Rev. Anti Infect. Ther. 2016, 14, 1139–1154. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, D.; Wu, M.; Teo, J.; Dick, T. Vancomycin and clarithromycin show synergy against Mycobacterium abscessus in vitro. Antimicrob. Agents Chemother. 2017, 61, e01298–e01317. [Google Scholar] [CrossRef] [PubMed]
- Roux, A.-L.; Catherinot, E.; Ripoll, F.; Soismier, N.; Macheras, E.; Ravilly, S.; Bellis, G.; Vibet, M.-A.; Le Roux, E.; Lemonnier, L.; et al. Multicenter study of prevalence of nontuberculous mycobacteria in patients with cystic fibrosis in France. J. Clin. Microbiol. 2009, 47, 4124–4128. [Google Scholar] [CrossRef] [PubMed]
- Mirsaeidi, M.; Farshidpour, M.; Allen, M.; Ebrahimi, G.; Falkinham, J. Highlight on advances in nontuberculous mycobacterial disease in North America. Biomed. Res. Int. 2014, 919474, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Leão, S.; Tortoli, E.; Euzéby, J.; Garcia, M. Proposal that Mycobacterium massiliense and Mycobacterium bolletii be united and reclassified as Mycobacterium abscessus subsp. bolletii comb. nov., designation of Mycobacterium abscessus subsp. abscessus subsp. nov. and emended description of Mycobacterium abscessus. Int. J. Syst. Evol. Microbiol. 2011, 61, 2311–2313. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Kim, B.; Kook, Y.; Yun, Y.-J.; Shin, J.; Hwan, J.; Kim, B.-J.; Kook, Y.-H. Mycobacterium massiliense is differentiated from Mycobacterium abscessus and Mycobacterium bolletii by erythromycin ribosome methyltransferase gene (erm) and clarithromycin susceptibility patterns. Microbiol. Immunol. 2010, 54, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Bouzinbi, N.; Chaturvedi, V.; Godreuil, S.; Kremer, L. In vitro evaluation of a new drug combination against clinical isolates belonging to the Mycobacterium abscessus complex. Clin. Microbiol. Infect. 2014, 20, O1124–O1127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lu, J.; Liu, M.; Wang, Y.; Zhao, Y.; Pang, Y. In vitro activity of clarithromycin in combination with other antimicrobial agents against Mycobacterium abscessus and Mycobacterium massiliense. Int. J. Antimicrob. Agents 2017, 49, 383–386. [Google Scholar] [CrossRef]
- Griffith, D.; Aksamit, T.; Brown-Elliott, B.; Catanzaro, A.; Daley, C.; Gordin, F.; Holland, S.; Horsburgh, R.; Huitt, G.; Iademarco, M.; et al. An official ATS/IDSA statement: Diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir. Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef]
- Clinical Laboratory Standards Institute. Susceptibility Testing of Mycobacteria, Nocardiae, and Other Aerobic Actinomycetes; Approved Standard; CLSI document M24-A2; Clinical Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2011. [Google Scholar]
- Wallace, R.; Meier, A.; Brown, B.; Zhang, Y.; Sander, P.; Onyi, G.; Böttger, E. Genetic basis for clarithromycin resistance among isolates of Mycobacterium chelonae and Mycobacterium abscessus. Antimicrob. Agents Chemother. 1996, 40, 1676–1681. [Google Scholar] [CrossRef]
- Nash, K.; Brown-Elliott, B.; Wallace, R. A novel gene, erm (41), confers inducible macrolide resistance to clinical isolates of Mycobacterium abscessus but is absent from Mycobacterium chelonae. Antimicrob. Agents Chemother. 2009, 53, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Bastian, S.; Veziris, N.; Roux, A.; Brossier, F.; Gaillard, J.; Jarlier, V.; Cambau, E. Assessment of clarithromycin susceptibility in strains belonging to the Mycobacterium abscessus group by erm41 and rrl sequencing. Antimicrob. Agents Chemother. 2011, 55, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Mougari, F.; Amarsy, R.; Veziris, N.; Bastian, S.; Brossier, F.; Berçot, B.; Raskine, L.; Cambau, E. Standardized interpretation of antibiotic susceptibility testing and resistance genotyping for Mycobacterium abscessus with regard to subspecies and erm41 sequevar. J. Antimicrob. Chemother. 2016, 71, 2208–2212. [Google Scholar] [CrossRef] [PubMed]
- Maurer, F.; Bruderer, V.; Castelberg, C.; Ritter, C.; Scherbakov, D.; Bloemberg, G.; Böttger, E. Aminoglycoside-Modifying enzymes determine the innate susceptibility to aminoglycoside antibiotics in rapidly growing mycobacteria. J. Antimicrob. Chemother. 2015, 70, 1412–1419. [Google Scholar] [CrossRef] [PubMed]
- Dubée, V.; Bernut, A.; Cortes, M.; Lesne, T.; Dorchene, D.; Lefebvre, A.; Hugonnet, J.-E.; Gutmann, F.; Mainardi, J.-L.; Herrmann, J.-L.; et al. β-Lactamase inhibition by avibactam in Mycobacterium abscessus. J. Antimicrob. Chemother. 2014, 70, 1051–1058. [Google Scholar] [CrossRef]
- Oh, W.; Stout, J.; Yew, W. Advances in the management of pulmonary disease due to Mycobacterium abscessus complex. Int. J. Tuberc. Lung Dis. 2014, 18, 1141–1148. [Google Scholar] [CrossRef]
- Diel, R.; Ringshausen, F.; Richter, E.; Welker, L.; Schmitz, J.; Nienhaus, A. Microbiological and clinical outcomes of treating non-Mycobacterium avium complex nontuberculous mycobacterial pulmonary disease: A systematic review and meta-analysis. Chest 2017, 152, 120–142. [Google Scholar] [CrossRef] [PubMed]
- Schmalstieg, A.; Srivastava, S.; Belkaya, S.; Deshpande, D.; Meek, C.; Leff, R.; van Oers, N.; Gumbo, T. The antibiotic resistance arrow of time: Efflux pump induction is a general first step in the evolution of mycobacterial drug resistance. Antimicrob. Agents Chemother. 2012, 56, 4806–4815. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.; Couto, I.; Perdigão, J.; Rodrigues, L.; Portugal, I.; Baptista, P.; Veigas, B.; Amaral, L.; Viveiros, M. Contribution of efflux to the emergence of isoniazid and multidrug resistance in Mycobacterium tuberculosis. PLoS ONE 2012, 7, e34538. [Google Scholar] [CrossRef]
- Ramis, I.; Vianna, J.; Silva, L., Jr.; von Groll, A.; Ramos, D.; Zanatta, N.; Viveiros, M.; da Silva, P.E.A. In silico and in vitro evaluation of tetrahydropyridine compounds as efflux inhibitors in Mycobacterium abscessus. Tuberculosis 2019, 118, 101853. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Richard, M.; Roquet-Banères, F.; Viljoen, A.; Kremer, L. The TetR-family transcription factor MAB_2299c regulates the expression of two distinct MmpS-MmpL efflux pumps involved in cross-resistance to clofazimine and bedaquiline in Mycobacterium abscessus. Antimicrob. Agents Chemother. 2019, AAC.01000-19. [Google Scholar] [CrossRef]
- Da Silva, P.E.A.; Machado, D.; Ramos, D.; Couto, I.; von Groll, A.; Viveiros, M. Efflux pumps in mycobacteria: Antimicrobial resistance, physiological functions, and role in pathogenicity. In Efflux-Mediated Antimicrobial Resistance in Bacteria; Li, X.-Z., Elkins, C.A., Zgurskaya, H.I., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 527–559. [Google Scholar] [CrossRef]
- Marquez, B. Bacterial efflux systems and efflux pumps inhibitors. Biochimie 2005, 87, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, H.M. Flow cytometry of bacterial membrane potential and permeability. Methods Mol. Med. 2008, 142, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Viveiros, M.; Rodrigues, L.; Martins, M.; Couto, I.; Spengler, G.; Martins, A.; Amaral, L. Evaluation of efflux activity of bacteria by a semi-automated fluorometric system. Methods Mol. Biol. 2010, 642, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.; Piddock, L. How to measure export via bacterial multidrug resistance efflux pumps. MBio 2016, 7, e00840–e00916. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.; Perdigão, J.; Portugal, I.; Pieroni, M.; Silva, P.; Couto, I.; Viveiros, M. Efflux activity differentially modulates the levels of isoniazid and rifampicin resistance among multidrug resistant and monoresistant Mycobacterium tuberculosis strains. Antibiotics 2018, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Tyagi, S.; Almeida, D.; Maiga, M.; Ammerman, N.; Bishai, W. Acceleration of tuberculosis treatment by adjunctive therapy with verapamil as an efflux inhibitor. Am. J. Respir. Crit. Care Med. 2013, 188, 600–607. [Google Scholar] [CrossRef]
- Rodrigues, L.; Sampaio, D.; Couto, I.; Machado, D.; Kern, W.; Amaral, L.; Viveiros, M. The role of efflux pumps in macrolide resistance in Mycobacterium avium complex. Int. J. Antimicrob. Agents 2009, 34, 529–533. [Google Scholar] [CrossRef]
- Versalovic, J.; Shortridge, D.; Kibler, K.; Griffy, M.; Beyer, J.; Flamm, R.; Tanaka, S.; Graham, D.; Go, M. Mutations in 23S rRNA are associated with clarithromycin resistance in Helicobacter pylori. Antimicrob. Agents Chemother. 1996, 40, 477–480. [Google Scholar] [CrossRef]
- Machado, D.; Coelho, T.; Perdigão, J.; Pereira, C.; Couto, I.; Portugal, I.; Maschmann, R.; Ramos, D.; von Groll, A.; Rossetti, M.; et al. Interplay between mutations and efflux in drug resistant clinical isolates of Mycobacterium tuberculosis. Front. Microbiol. 2017, 8, 711. [Google Scholar] [CrossRef]
- Piddock, L. Multidrug-resistance efflux pumps? not just for resistance. Nat. Rev. Microbiol. 2006, 4, 629. [Google Scholar] [CrossRef] [PubMed]
- Koh, W.; Jeon, K.; Lee, N.; Kim, B.; Kook, Y.; Lee, S.; Park, Y.; Kim, C.; Shin, S.; Huitt, G.; et al. Clinical significance of differentiation of Mycobacterium massiliense from Mycobacterium abscessus. Am. J. Respir. Crit. Care Med. 2011, 183, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Akiyama, Y.; Kurashima, A.; Nagai, H.; Tsuyuguchi, K.; Fujii, T.; Yano, S.; Shigeto, E.; Kuraoka, T.; Kajiki, A.; et al. Clinical and microbiological differences between Mycobacterium abscessus and Mycobacterium massiliense lung diseases. J. Clin. Microbiol. 2012, 50, 3556–3561. [Google Scholar] [CrossRef] [PubMed]
- Nash, K.; Inderlied, C. Genetic basis of macrolide resistance in Mycobacterium avium isolated from patients with disseminated disease. Antimicrob. Agents Chemother. 1995, 29, 2625–2630. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, G.; Moellering, R., Jr. Antimicrobial combinations. In Antibiotics in Laboratory Medicine, 4th ed.; Lorian, V., Ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1996; pp. 330–396. [Google Scholar]
- Caviedes, L.; Delgado, J.; Gilman, R. Tetrazolium microplate assay as a rapid and inexpensive colorimetric method for determination of antibiotic susceptibility of Mycobacterium tuberculosis. J. Clin. Microbiol. 2002, 40, 1873–1874. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Flores, R.; Gupta, S.; Tamez-Guerra, R.; Mehta, R. Determination of MICs for Mycobacterium avium-M. intracellulare complex in liquid medium by a colorimetric method. J. Clin. Microbiol. 1995, 33, 1842–1846. [Google Scholar] [PubMed]
- Gröblacher, B.; Kunert, O.; Bucar, F. Compounds of Alpinia katsumadai as potential efflux inhibitors in Mycobacterium smegmatis. Bioorg. Med. Chem. 2012, 20, 2701–2706. [Google Scholar] [CrossRef]
- Coelho, T.; Machado, D.; Couto, I.; Maschmann, R.; Ramos, D.; von Groll, A.; Rossetti, M.; da Silva, P.A.; Viveiros, M. Enhancement of antibiotic activity by efflux inhibitors against multidrug resistant Mycobacterium tuberculosis clinical isolates from Brazil. Front. Microbiol. 2015, 6, 330. [Google Scholar] [CrossRef]
- Machado, L.; Spengler, G.; Evaristo, M.; Handzlik, J.; Molnár, J.; Viveiros, M.; Kiec-Kononowicz, K.; Amaral, L. Biological activity of twenty-three hydantoin derivatives on intrinsic efflux pump system of Salmonella enterica serovar Enteritidis NCTC 13349. In Vivo 2011, 25, 769–772. [Google Scholar]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strain | Species/Subspecies | Genetic Background | MICs (µg/mL), Day 3 * | |||||
|---|---|---|---|---|---|---|---|---|
| CLA | AMK | Compounds | ||||||
| erm(41) | rrl | rs | CLA | AMK | VP | EtBr | ||
| MabATCC19977T | M. abscessus subsp. abscessus | T28 | wt | wt | 2 (S) | 16 (S) | 1024 | 128 |
| MabBR1 | M. abscessus subsp. abscessus | T28 | wt | wt | 2 (S) | 8 (S) | 4096 | 256 |
| MabBR2 | M. abscessus subsp. bolletii | T28 | wt | wt | 1 (S) | 4 (S) | 1024 | 64 |
| MabBR3 | M. abscessus subsp. massiliense | T28 | A2059C | wt | 256 (R) | 16 (S) | 1024 | 32 |
| MabPT1 | M. abscessus subsp. abscessus | T28 | wt | wt | 0.25 (S) | 4 (S) | 1024 | 64 |
| MabPT2 | M. abscessus subsp. bolletii | T28 | wt | wt | 2 (S) | 4 (S) | 2048 | 128 |
| MabPT3 | M. abscessus subsp. massiliense | T28 | wt | wt | 0.5 (S) | 4 (S) | 1024 | 256 |
| MabPT4 | M. abscessus subsp. bolletii | T28 | wt | wt | 2 (S) | 4 (S) | 1024 | 128 |
| MabPT5 | M. abscessus subsp. massiliense | T28 | wt | wt | 1 (S) | 4 (S) | 1024 | 256 |
| MabPT6 | M. abscessus subsp. abscessus | T28 | wt | wt | 1 (S) | 4 (S) | 2048 | 64 |
| Strain | Mutations | CLA MICs (µg/mL) | CLA Susceptibility Profile at Day 14 * | |||
|---|---|---|---|---|---|---|
| erm(41) | rrl | Day 5 | Day 14 | |||
| MabATCC19977T | T28 | wt | 4 (I) | 16 (R) | Resistant | Inducible |
| MabBR1 | T28 | wt | 2 (S) | 4 (I) | Intermediate | Non-inducible |
| MabBR2 | T28 | wt | 16 (R) | 256 (R) | Resistant | Non-inducible |
| MabBR3 | T28 | A2059C | 256 (R) | 256 (R) | Resistant | Non-inducible |
| MabPT1 | T28 | wt | 0.25 (S) | 2 (S) | Susceptible | Non-inducible |
| MabPT2 | T28 | wt | 4 (I) | 16 (R) | Resistant | Inducible |
| MabPT3 | T28 | wt | 0.5 (S) | 0.5 (S) | Susceptible | Non-inducible |
| MabPT4 | T28 | wt | 16 (R) | 64 (R) | Resistant | Non-inducible |
| MabPT5 | T28 | wt | 1 (S) | 1 (S) | Susceptible | Non-inducible |
| MabPT6 | T28 | wt | 2 (S) | 16 (R) | Resistant | Inducible |
| Strain | MIC (µg/mL) (MF) | |||||
|---|---|---|---|---|---|---|
| Day 3 | Day 5 | Day 14 | ||||
| CLA | CLA+VP | CLA | CLA+VP | CLA | CLA+VP | |
| MabATCC19977T | 2 (S) | 0.5 (S) (↓4) | 4 (I) | 1 (S) (↓4) | 16 (R) | 16 (R) |
| MabBR1 | 2 (S) | 1 (S) (↓2) | 2 (S) | 2 (S) | 4 (I) | 4 (I) |
| MabBR2 | 1 (S) | 0.25 (S) (↓4) | 16 (R) | 2 (S) (↓8) | 256 (R) | 128 (R) (↓2) |
| MabBR3 | 256 (R) | 256 (R) | 256 (R) | 256 (R) | 256 (R) | 256 (R) |
| MabPT1 | 0.25 (S) | 0.0625 (S) (↓4) | 0.25 (S) | 0.0625 (S) (↓4) | 2 (S) | 0.5 (S) (↓4) |
| MabPT2 | 2 (S) | ≤0.0625 (S) (≥↓32) | 4 (I) | ≤0.0625 (S) (≥↓64) | 16 (R) | 16 (R) |
| MabPT3 | 0.5 (S) | 0.0625 (S) (↓4) | 0.5 (S) | 0.25 (S) (↓2) | 0.5 (S) | 0.5 (S) |
| MabPT4 | 2 (S) | 0.5 (S) (↓4) | 16 (R) | 4 (I) (↓4) | 64 (R) | 32 (R) (↓2) |
| MabPT5 | 1 (S) | 0.5 (S) (↓2) | 1 (S) | 0.5 (S) (↓2) | 1 (S) | 0.5 (S) (↓2) |
| MabPT6 | 1 (S) | ≤0.03125 (S) (≥↓32) | 2 (S) | 0.125 (S) (↓16) | 16 (R) | 8 (R) (↓2) |
| Strains | CEq (μg/mL) | RFFVP |
|---|---|---|
| MabATCC19977T | 0.25 | 1.15 ± 0.03 |
| MabBR1 | 0.125 | 0.78 ± 0.01 |
| MabBR2 | 0.25 | 1.12 ± 0.06 |
| MabBR3 | 0.125 | 0.19 ± 0.05 |
| MabPT1 | 0.25 | 1.14 ± 0.01 |
| MabPT2 | 0.25 | 0.63 ± 0.01 |
| MabPT3 | 0.5 | 1.61 ± 0.05 |
| MabPT4 | 0.125 | 1.61 ± 0.01 |
| MabPT5 | 0.5 | 0.26 ± 0.00 |
| MabPT6 | 0.25 | 1.64 ± 0.11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vianna, J.S.; Machado, D.; Ramis, I.B.; Silva, F.P.; Bierhals, D.V.; Abril, M.A.; von Groll, A.; Ramos, D.F.; Lourenço, M.C.S.; Viveiros, M.; et al. The Contribution of Efflux Pumps in Mycobacterium abscessus Complex Resistance to Clarithromycin. Antibiotics 2019, 8, 153. https://doi.org/10.3390/antibiotics8030153
Vianna JS, Machado D, Ramis IB, Silva FP, Bierhals DV, Abril MA, von Groll A, Ramos DF, Lourenço MCS, Viveiros M, et al. The Contribution of Efflux Pumps in Mycobacterium abscessus Complex Resistance to Clarithromycin. Antibiotics. 2019; 8(3):153. https://doi.org/10.3390/antibiotics8030153
Chicago/Turabian StyleVianna, Júlia S., Diana Machado, Ivy B. Ramis, Fábia P. Silva, Dienefer V. Bierhals, Michael Andrés Abril, Andrea von Groll, Daniela F. Ramos, Maria Cristina S. Lourenço, Miguel Viveiros, and et al. 2019. "The Contribution of Efflux Pumps in Mycobacterium abscessus Complex Resistance to Clarithromycin" Antibiotics 8, no. 3: 153. https://doi.org/10.3390/antibiotics8030153
APA StyleVianna, J. S., Machado, D., Ramis, I. B., Silva, F. P., Bierhals, D. V., Abril, M. A., von Groll, A., Ramos, D. F., Lourenço, M. C. S., Viveiros, M., & da Silva, P. E. A. (2019). The Contribution of Efflux Pumps in Mycobacterium abscessus Complex Resistance to Clarithromycin. Antibiotics, 8(3), 153. https://doi.org/10.3390/antibiotics8030153