Origanum vulgare Essential Oil vs. a Commercial Mixture of Essential Oils: In Vitro Effectiveness on Salmonella spp. from Poultry and Swine Intensive Livestock



 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Compositional Analysis
2.2. Antibiograms
2.3. Broth Microdilution Susceptibility Testing
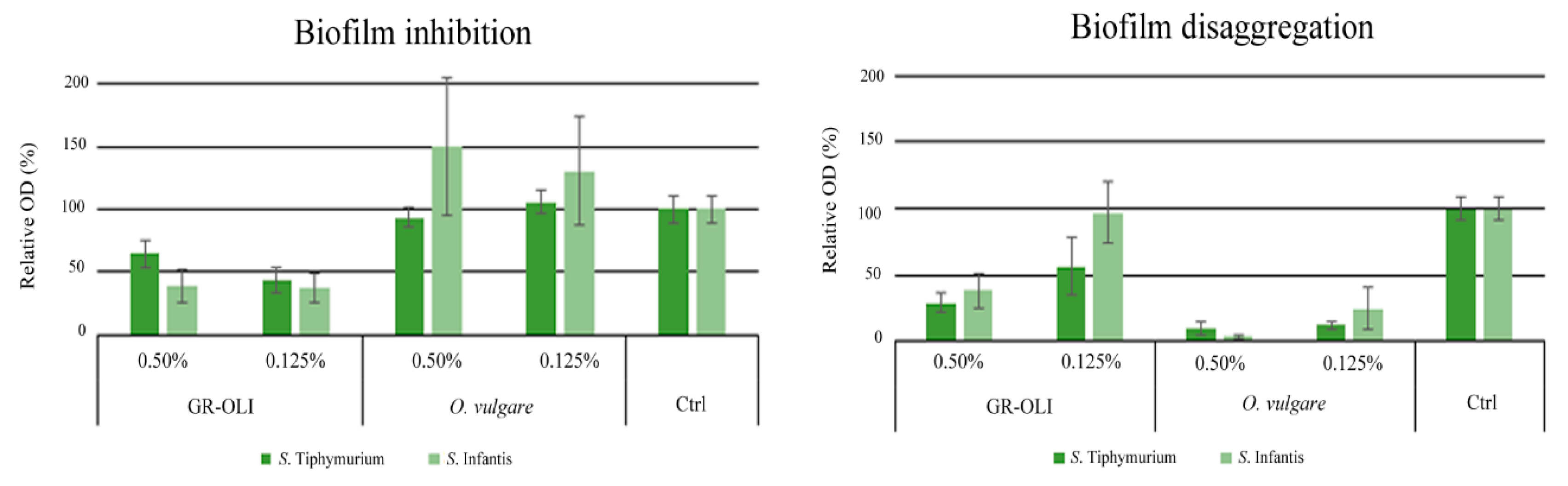
2.4. Biofilm Formation Assay
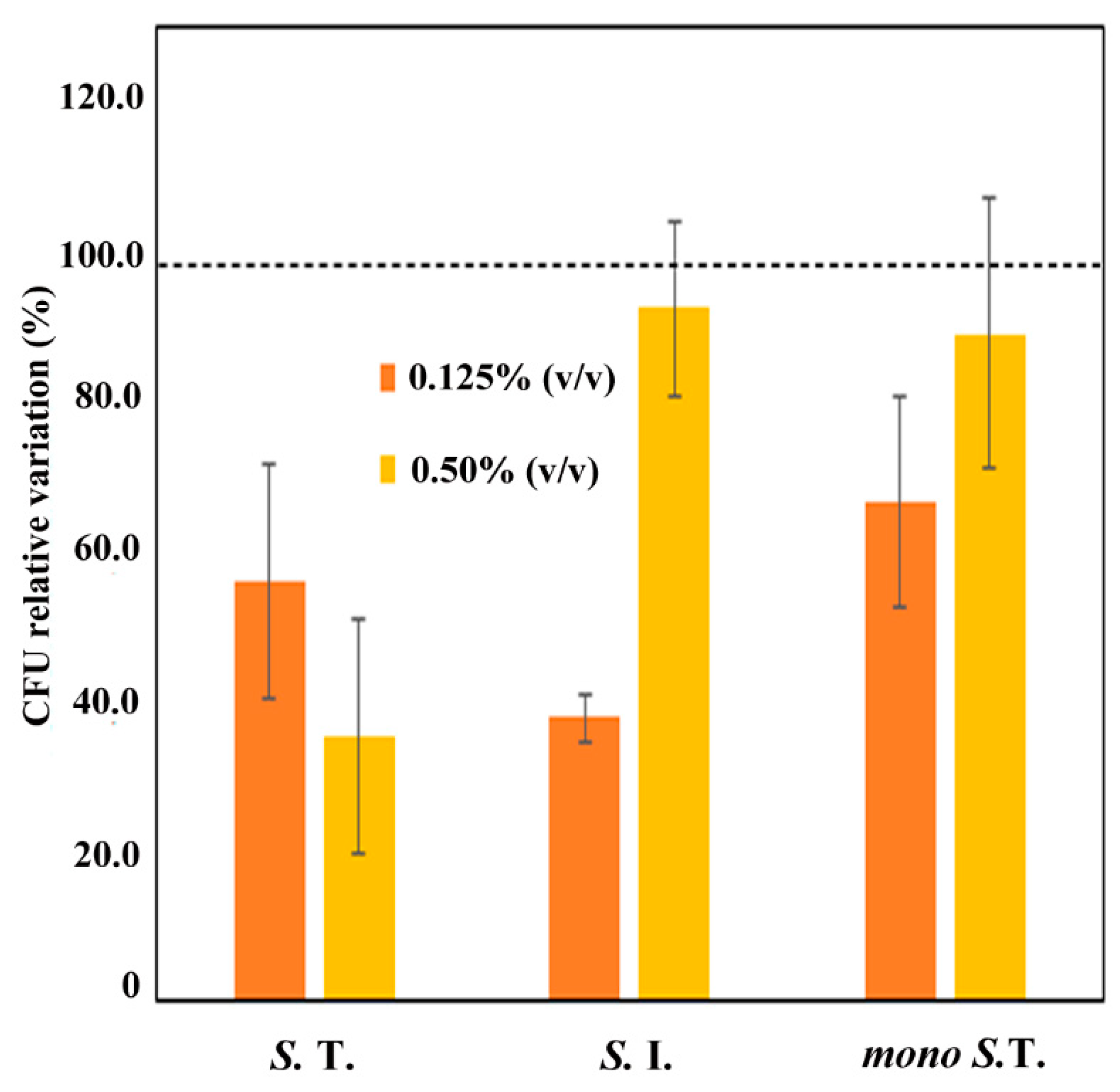
2.5. Cell Adhesion Assay
2.6. Growth Curves
2.7. Checkerboard Titration Method
3. Discussion
4. Materials and Methods
4.1. Natural Substances, Antibiotics, and Reagents
4.2. Bacterial Strains and Growth Media
4.3. GC-MS Analysis
4.4. GC-FID Analysis
4.5. Qualitative and Semi-Quantitative Analysis
4.6. Antimicrobial Susceptibility Testing against Antibiotics
4.7. Broth Microdilution Susceptibility Testing against Natural Products
4.8. Biofilm Assay
4.9. Cell Adhesion Assay
4.10. Growth Curves
4.11. Checkerboard Titration Method
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foley, S.L.; Lynne, A.M.; Nayak, R. Salmonella challenges: Prevalence in swine and poultry and potential pathogenicity of such isolates. J. Anim. Sci. 2008, 86, E149–E162. [Google Scholar] [CrossRef] [PubMed]
- Boyen, F.; Haesebrouck, F.; Maes, D.; Van Immerseel, F.; Ducatelle, R.; Pasmans, F. Non-typhoidal Salmonella infections in pigs: A closer look at epidemiology, pathogenesis and control. Vet. Microbiol. 2008, 130, 1–19. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, 5926. [Google Scholar]
- EFSA. Salmonella control in poultry flocks and its public health impact. EFSA J. 2019, 17. [Google Scholar] [CrossRef]
- Pornsukarom, S.; Thakur, S. Horizontal Dissemination of Antimicrobial Resistance Determinants in Multiple Salmonella Serotypes following Isolation from the Commercial Swine Operation Environment after Manure Application. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Opinion of the scientific panel on biological hazards on “risk assessment and mitigation options of Salmonella in pig production”. EFSA J. 2006, 4, 341. [Google Scholar]
- European Union Ban on Antibiotics as Growth Promoters in Animal Feed Enters into Effect. 2006. Available online: https://ec.europa.eu/commission/presscorner/detail/en/IP_05_1687 (accessed on 31 October 2020).
- Thacker, P.A. Alternatives to antibiotics as growth promoters for use in swine production: A review. J. Anim. Sci. Biotechnol. 2013, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential oils as alternatives to antibiotics in swine production. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2018, 4, 126–136. [Google Scholar] [CrossRef]
- International Organization for Standardization. ISO 9235:2013(en): Aromatic Natural Raw Materials—Vocabulary; International Organization for Standardization (ISO): Geneva, Switzerland, 2013. [Google Scholar]
- Amerah, A.M.; Mathis, G.; Hofacre, C.L. Effect of xylanase and a blend of essential oils on performance and Salmonella colonization of broiler chickens challenged with Salmonella Heidelberg. Poult. Sci. 2012, 91, 943–947. [Google Scholar] [CrossRef]
- Walia, K.; Argüello, H.; Lynch, H.; Leonard, F.C.; Grant, J.; Yearsley, D.; Kelly, S.; Duffy, G.; Gardiner, G.E.; Lawlor, P.G. Effect of strategic administration of an encapsulated blend of formic acid, citric acid, and essential oils on Salmonella carriage, seroprevalence, and growth of finishing pigs. Prev. Vet. Med. 2017, 137, 28–35. [Google Scholar] [CrossRef]
- Zhang, S.; Shen, Y.R.; Wu, S.; Xiao, Y.Q.; He, Q.; Shi, S.R. The dietary combination of essential oils and organic acids reduces Salmonella enteritidis in challenged chicks. Poult. Sci. 2019, 98, 6349–6355. [Google Scholar] [CrossRef]
- Si, W.; Gong, J.; Tsao, R.; Zhou, T.; Yu, H.; Poppe, C.; Johnson, R.; Du, Z. Antimicrobial activity of essential oils and structurally related synthetic food additives towards selected pathogenic and beneficial gut bacteria. J. Appl. Microbiol. 2006, 100, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Oussalah, M.; Caillet, S.; Salmiéri, S.; Saucier, L.; Lacroix, M. Antimicrobial effects of alginate-based films containing essential oils on Listeria monocytogenes and Salmonella typhimurium present in bologna and ham. J. Food Prot. 2007, 70, 901–908. [Google Scholar] [CrossRef]
- Fratianni, F.; De Martino, L.; Melone, A.; De Feo, V.; Coppola, R.; Nazzaro, F. Preservation of chicken breast meat treated with thyme and balm essential oils. J. Food Sci. 2010, 75, M528–M535. [Google Scholar] [CrossRef] [PubMed]
- Casco, G.; Taylor, T.M.; Alvarado, C. Evaluation of novel micronized encapsulated essential oil-containing phosphate and lactate blends for growth inhibition of Listeria monocytogenes and Salmonella on poultry bologna, pork ham, and roast beef ready-to-eat deli loaves. J. Food Prot. 2015, 78, 698–706. [Google Scholar] [CrossRef]
- Boskovic, M.; Djordjevic, J.; Ivanovic, J.; Janjic, J.; Zdravkovic, N.; Glisic, M.; Glamoclija, N.; Baltic, B.; Djordjevic, V.; Baltic, M. Inhibition of Salmonella by thyme essential oil and its effect on microbiological and sensory properties of minced pork meat packaged under vacuum and modified atmosphere. Int. J. Food Microbiol. 2017, 258, 58–67. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Safety and efficacy of an essential oil from Origanum vulgare ssp. hirtum (Link) Ietsw for all animal species. EFSA 2019, 17, e05909. [Google Scholar]
- Rodriguez-Garcia, I.; Silva-Espinoza, B.A.; Ortega-Ramirez, L.A.; Leyva, J.M.; Siddiqui, M.W.; Cruz-Valenzuela, M.R.; Gonzalez-Aguilar, G.A.; Ayala-Zavala, J.F. Oregano Essential Oil as an Antimicrobial and Antioxidant Additive in Food Products. Crit. Rev. Food Sci. Nutr. 2016, 56, 1717–1727. [Google Scholar] [CrossRef]
- Numpaque, M.A.; Oviedo, L.A.; Gil, J.H.; García, C.M.; Durango, D.L. Thymol and carvacrol: Biotransformation and antifungal activity against the plant pathogenic fungi Colletotrichum acutatum and Botryodiplodia theobromae. Trop. Plant Pathol. 2011, 36, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Ahmad, A.; Ahmad Khan, L.; Padoa, C.J.; van Vuuren, S.; Manzoor, N. Effect of two monoterpene phenols on antioxidant defense system in Candida albicans. Microb. Pathog. 2015, 80, 50–56. [Google Scholar] [CrossRef]
- Gumus, T.; Demirci, A.S.; Sagdic, O.; Arici, M. Inhibition of heat resistant molds: Aspergillus fumigatus and Paecilomyces variotii by some plant essential oils. Food Sci. Biotechnol. 2010, 19, 1241–1244. [Google Scholar] [CrossRef]
- Guarda, A.; Rubilar, J.F.; Miltz, J.; Galotto, M.J. The antimicrobial activity of microencapsulated thymol and carvacrol. Int. J. Food Microbiol. 2011, 146, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Van de Vel, E.; Sampers, I.; Raes, K. A review on influencing factors on the minimum inhibitory concentration of essential oils. Crit. Rev. Food Sci. Nutr. 2019, 59, 357–378. [Google Scholar] [CrossRef] [PubMed]
- WHO Website. Available online: https://www.who.int/ (accessed on 31 October 2020).
- CDC Website. Available online: https://www.cdc.gov/ (accessed on 31 October 2020).
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- International Organization for Standardization (ISO). ISO 6579:2002: Microbiology of Food and Animal Feeding Stuffs–Horizontal Method for the Detection of Salmonella spp.–Amendment 1: Annex D: Detection of Salmonella spp. in Animal Faeces and in Environmental Samples from the Primary Production Stage; International Organization for Standardization (ISO): Geneva, Switzerland, 2007. [Google Scholar]
- International Organization for Standardization (ISO). ISO/TR 6579-3:2014: Microbiology of the Food Chain–Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella. Part 3: Guidelines For serotyping of Salmonella spp.; International Organization for Standardization (ISO): Geneva, Switzerland, 2014. [Google Scholar]
- Grimont, P.A.D.; Weill, F.X. Antigenic Formulae of the Salmonella Serovars. Available online: https://www.pasteur.fr/ip/portal/action/WebdriveActionEvent/oid/01s-000036-089 (accessed on 31 October 2020).
- Adams, R. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy. Carol Stream 2007, 16, 65–120. [Google Scholar]
- Rodriguez-Tudela, J.L.; Arendrup, M.C.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Cuenca-Estrella, M.; Dannaoui, E.; Denning, D.W.; Donnelly, J.P.; Dromer, F.; et al. EUCAST definitive document EDef 7.1: Method for the determination of broth dilution MICs of antifungal agents for fermentative yeasts. Clin. Microbiol. Infect. 2008, 14, 398–405. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| % | |||
|---|---|---|---|
| Components | LRI | O. vulgare | GR-OLI |
| α−thujene | 927 | 0.82 | 0.13 |
| α−pinene | 934 | 0.98 | 2.48 |
| camphene | 948 | 0.13 | 0.24 |
| sabinene | 974 | n.d. | 0.38 |
| β−pinene | 976 | 0.15 | 2.20 |
| octen-3-ol | 980 | 0.46 | n.d. |
| 2-octanone | 988 | 0.22 | n.d. |
| β-myrcene | 993 | 1.51 | 0.90 |
| 3-octanol | 998 | 0.05 | n.d. |
| α−phellandrene | 1005 | 0.18 | n.d. |
| α− terpinene | 1017 | 1.06 | 1.24 |
| p-cymene | 1026 | 7.04 | 10.62 |
| limonene | 1029 | 0.47 | 15.32 |
| 1,8-cineol | 1034 | n.d. | 11.95 |
| cis-ocimene | 1040 | n.d. | 1.36 |
| trans ocimene | 1049 | 0.05 | 0.22 |
| γ−terpinene | 1060 | 5.50 | 3.80 |
| trans sabinene hydrate | 1067 | 0.18 | n.d. |
| cis linalool oxide | 1073 | n.d. | 0.11 |
| terpinolene | 1089 | 0.15 | 0.63 |
| linalool | 1102 | 1.48 | 8.73 |
| fenchol | 1115 | n.d. | 0.21 |
| camphor | 1146 | n.d. | 0.45 |
| borneol | 1167 | 0.19 | 0.44 |
| terpinen-4-ol | 1179 | 0.53 | 7.12 |
| p-cimen-8-ol | 1188 | n.d. | 0.10 |
| α-terpineol | 1193 | n.d. | 2.75 |
| linalyl acetate | 1263 | n.d. | 5.03 |
| thymol | 1296 | 2.78 | 5.98 |
| carvacrol | 1315 | 66.98 | 12.50 |
| neryl acetate | 1369 | n.d. | 0.14 |
| geranyl acetate | 1387 | n.d. | 0.21 |
| β−caryophyllene | 1427 | 1.64 | 0.65 |
| α-trans bergamotene | 1442 | n.d. | 0.08 |
| aromadendrene | 1446 | n.d. | 0.24 |
| β-farnesene | 1462 | n.d. | 0.27 |
| γ-cadinene | 1512 | n.d. | 0.09 |
| caryophyllene oxide | 1594 | 0.14 | n.d. |
| α-bisabolool | 1694 | n.d. | 0.39 |
| D. | Origin | Sample Source | SP. | AMC | TZP | CTX | CAZ | FEP | ETP | IPM | MEM | AMK | GEN | CIP | SXT | GR-OLI | O. vulgare |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | Swine | Feces | S.T. | S | S | S | S | S | S | S | S | S | S | R | S | 4 | 0.25 |
| 7 | Swine | Feces | S.T. | S | S | S | S | S | S | S | S | S | S | R | S | 8 | 2 |
| 12 | Swine | Feces | S.T. | S | S | S | S | S | S | S | S | S | S | R | S | 8 | 0.5 |
| 13 | Swine | Feces | S.T. | S | S | S | S | S | S | S | S | S | S | S | S | 4 | 0.5 |
| 17 | Swine | Feces | S.T. | S | S | S | S | S | S | S | S | S | R | R | S | 16 | 2 |
| 18 | Swine | Feces | S.T. | S | S | S | S | S | S | S | S | S | S | R | S | 16 | 2 |
| 21 | Swine | Feces | S.T. | S | S | S | S | S | S | S | S | S | S | R | S | 8 | 2 |
| 24 | Swine | Feces | S.T. | R | S | S | S | S | S | S | S | S | S | S | R | 16 | 1 |
| 30 | Swine | Feces | S.T. | S | S | S | S | S | S | S | S | S | S | S | S | 16 | 0.5 |
| 31 | Swine | Feces | S.T. | R | S | S | S | S | S | S | S | S | S | S | R | 8 | 1 |
| 32 | Swine | Feces | S.T. | R | S | S | S | S | S | S | S | S | R | R | S | 8 | 1 |
| 33 | Swine | Feces | S.T. | S | S | S | S | S | S | S | S | S | S | S | S | 8 | 1 |
| 34 | Swine | Feces | S.T. | R | S | S | S | S | S | S | S | S | S | S | S | 2 | <0.25 |
| 19 | Swine | Feces | m.S.T. | R | I | S | S | S | S | S | S | S | S | R | S | 4 | 1 |
| 27 | Swine | Feces | m.S.T. | R | S | S | S | S | S | S | S | S | S | S | S | 4 | 1 |
| 28 | Swine | Feces | m.S.T. | R | S | S | S | S | S | S | S | S | S | R | S | 16 | 1 |
| 29 | Swine | Feces | m.S.T. | R | S | S | I | S | S | S | S | S | S | S | R | 0.5 | <0.25 |
| 4 | Chicken | Boot swabs | S.I. | S | S | S | S | S | S | S | S | S | S | R | R | 8 | <0.25 |
| 10 | Chicken | Boot swabs | S.I. | S | S | R | I | I | S | S | S | S | S | R | S | 8 | 1 |
| 25 | Chicken | Boot swabs | S.I. | R | S | S | S | S | S | S | S | S | S | R | R | 16 | 1 |
| 26 | Chicken | Boot swabs | S.I. | R | S | R | I | R | S | S | S | S | S | R | S | 16 | 1 |
| 35 | Chicken | Boot swabs | S.I. | R | S | R | I | R | S | S | S | S | S | R | R | 4 | 0.5 |
| 36 | Chicken | Boot swabs | S.I. | R | S | R | I | R | S | S | S | S | S | R | R | >16 | 1 |
| 37 | Chicken | Boot swabs | S.I. | R | S | R | I | R | S | S | S | S | S | R | R | 16 | <0.25 |
| 38 | Chicken | Boot swabs | S.I. | R | S | S | S | S | S | S | S | S | S | R | R | 16 | 1 |
| 39 | Chicken | Boot swabs | S.I. | R | S | S | S | S | S | S | S | S | S | R | R | 16 | 1 |
| 40 | Chicken | Boot swabs | S.I. | S | S | R | I | R | S | S | S | S | S | R | R | 4 | 0.5 |
| 41 | Chicken | Boot swabs | S.I. | R | S | R | I | R | S | S | S | S | S | R | R | >16 | 1 |
| 42 | Chicken | Boot swabs | S.I. | S | S | S | S | S | S | S | S | S | S | R | S | 16 | <0.25 |
| MIC90 | 16 | 2 |
| Strain | S. Typhimurium | S. Infantis | ||||
|---|---|---|---|---|---|---|
| Product | 10 h | 15 h | 20 h | 10 h | 15 h | 20 h |
| Ctrl | 100 a | 100 a | 100 a | 100 a | 100 a | 100 a |
| O. vulgare EO | 57 b | 63 c | 66 b | 64 b | 67 b | 67 b |
| GR-OLI | 65 b | 82 b | 99 a | 66 b | 64 b | 64 b |
| Strain | MBCAMC | MBCCTX | MBCCIP | MBCGR | MBCAMC/GR | MBCCTX/GR | MBCCIP/GR | FBCIAMC/GR | FBCICTX/GR | FBCICIP/GR |
|---|---|---|---|---|---|---|---|---|---|---|
| 26 | 32 | >128 | 0.5 | 12.5 | 0.25/1.56 | 0.5/1.56 | 0.01/3.125 | 0.133 (s) | 0.270 (s) | 0.128 (s) |
| 35 | 64 | 128 | 0.5 | 12.5 | 0.125/1.56 | 0.25/3.12 | 0.03/6.25 | 0.127 (s) | 0.530 (a) | 0.321 (s) |
| 36 | 64 | 128 | 0.5 | 12.5 | 0.25/6.25 | 0.5/1.56 | 0.005/6.25 | 0.504 (a) | 0.505 (a) | 0.128 (s) |
| 37 | 64 | >128 | 0.5 | 12.5 | 0.125/12.5 | 0.25/3.12 | 0.03/3.125 | 1.002 (i) | 0.280 (s) | 0.251 (s) |
| Name | O-Antigens | H-Antigens |
|---|---|---|
| Salmonella Typhimurium | 1,4,[5],12 | I:1,2 |
| Salmonella Typhimurium monophasic variant | 1,4,[5],12 | i:- |
| Salmonella Infantis | 6,7,14 | r:1,5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Vito, M.; Cacaci, M.; Barbanti, L.; Martini, C.; Sanguinetti, M.; Benvenuti, S.; Tosi, G.; Fiorentini, L.; Scozzoli, M.; Bugli, F.; et al. Origanum vulgare Essential Oil vs. a Commercial Mixture of Essential Oils: In Vitro Effectiveness on Salmonella spp. from Poultry and Swine Intensive Livestock. Antibiotics 2020, 9, 763. https://doi.org/10.3390/antibiotics9110763
Di Vito M, Cacaci M, Barbanti L, Martini C, Sanguinetti M, Benvenuti S, Tosi G, Fiorentini L, Scozzoli M, Bugli F, et al. Origanum vulgare Essential Oil vs. a Commercial Mixture of Essential Oils: In Vitro Effectiveness on Salmonella spp. from Poultry and Swine Intensive Livestock. Antibiotics. 2020; 9(11):763. https://doi.org/10.3390/antibiotics9110763
Chicago/Turabian StyleDi Vito, Maura, Margherita Cacaci, Lorenzo Barbanti, Cecilia Martini, Maurizio Sanguinetti, Stefania Benvenuti, Giovanni Tosi, Laura Fiorentini, Maurizio Scozzoli, Francesca Bugli, and et al. 2020. "Origanum vulgare Essential Oil vs. a Commercial Mixture of Essential Oils: In Vitro Effectiveness on Salmonella spp. from Poultry and Swine Intensive Livestock" Antibiotics 9, no. 11: 763. https://doi.org/10.3390/antibiotics9110763
APA StyleDi Vito, M., Cacaci, M., Barbanti, L., Martini, C., Sanguinetti, M., Benvenuti, S., Tosi, G., Fiorentini, L., Scozzoli, M., Bugli, F., & Mattarelli, P. (2020). Origanum vulgare Essential Oil vs. a Commercial Mixture of Essential Oils: In Vitro Effectiveness on Salmonella spp. from Poultry and Swine Intensive Livestock. Antibiotics, 9(11), 763. https://doi.org/10.3390/antibiotics9110763