Inhibition of Pneumolysin Cytotoxicity by Hydrolysable Tannins

 , ,
, , 
Abstract
:1. Introduction
2. Results
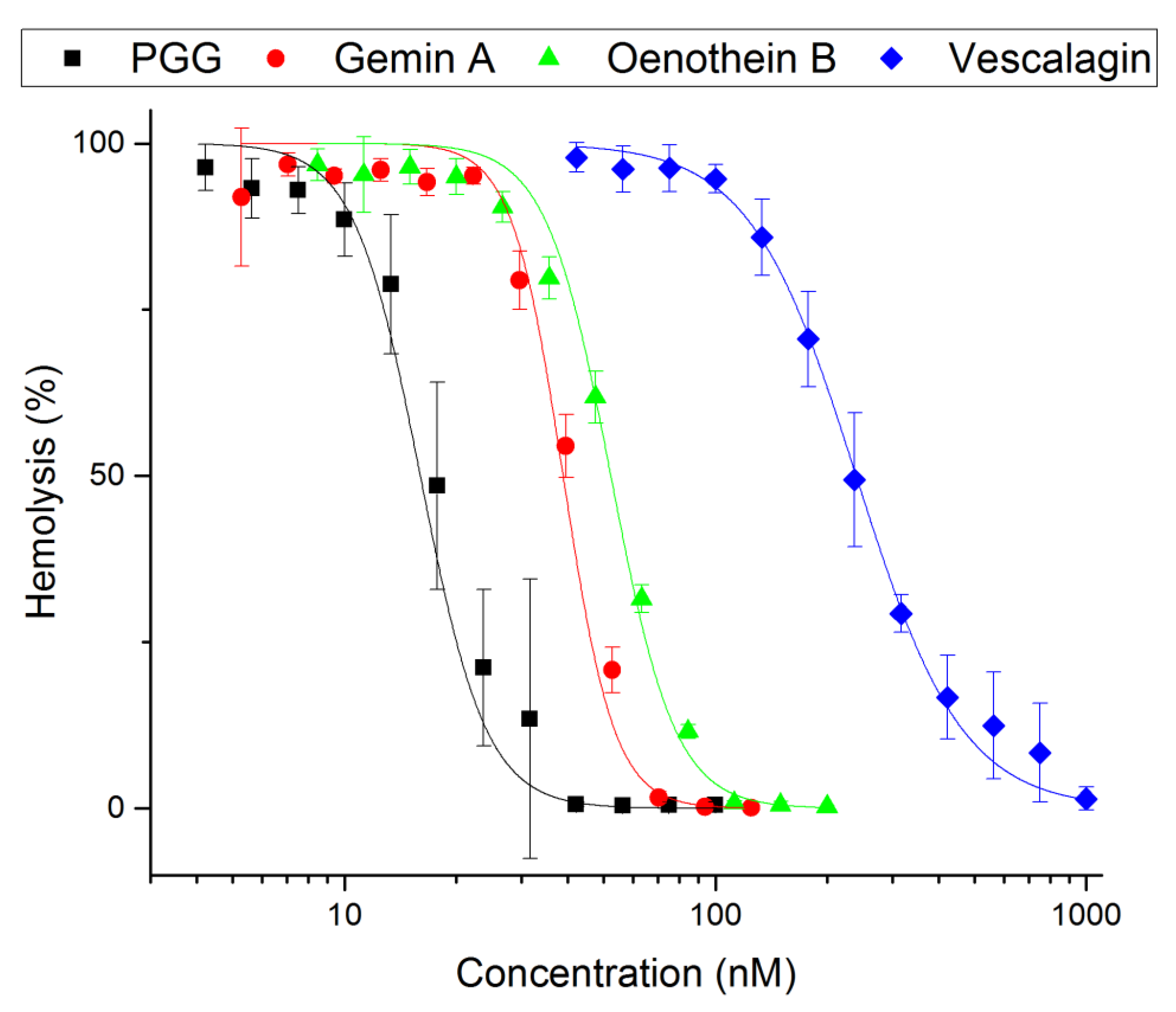
2.1. Inhibition of Hemolysis by Hydrolysable Tannins
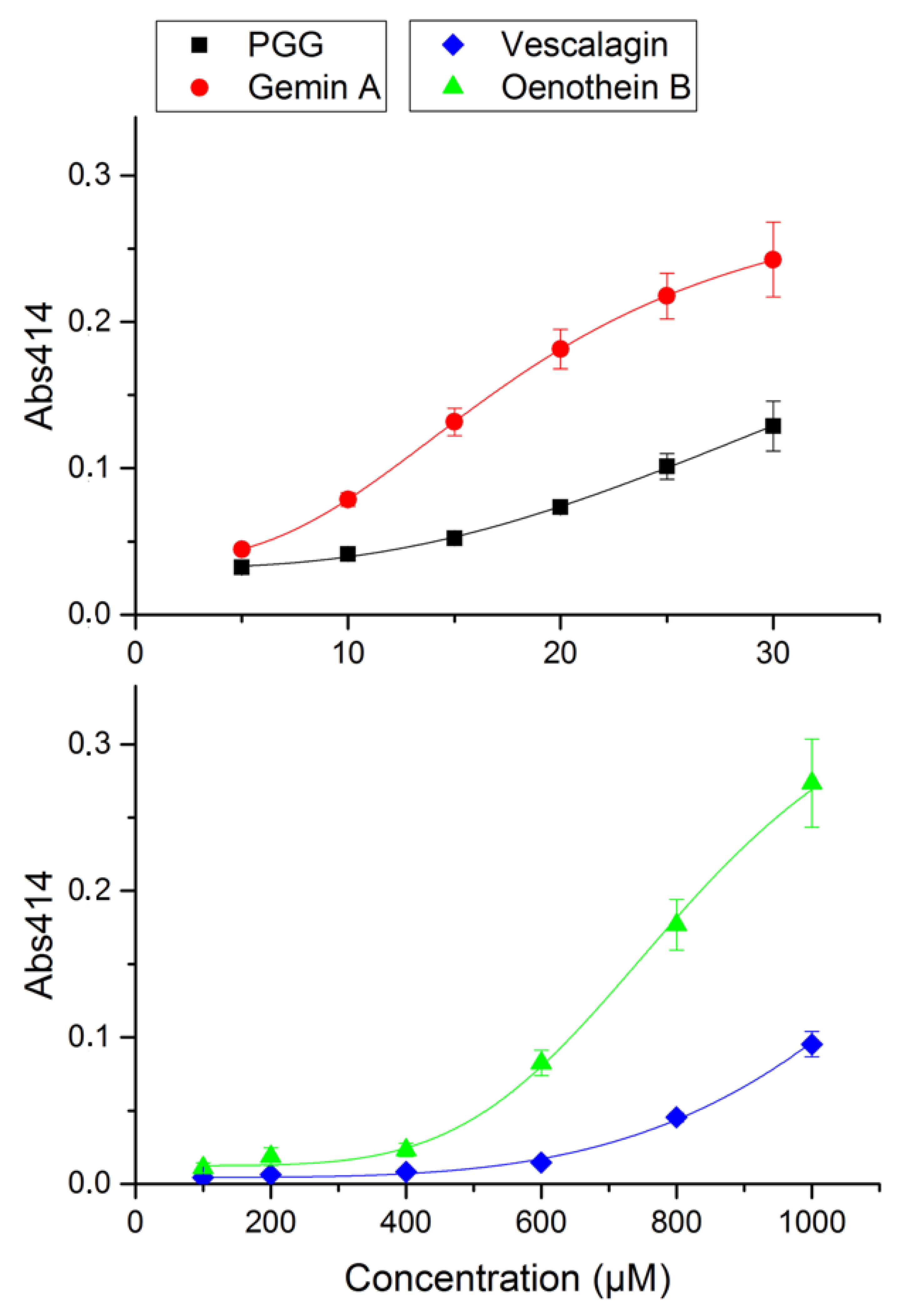
2.2. Inhibitory Power of PGG and Gemin A
2.3. PGG Abolishes Oligomerization of Pneumolysin
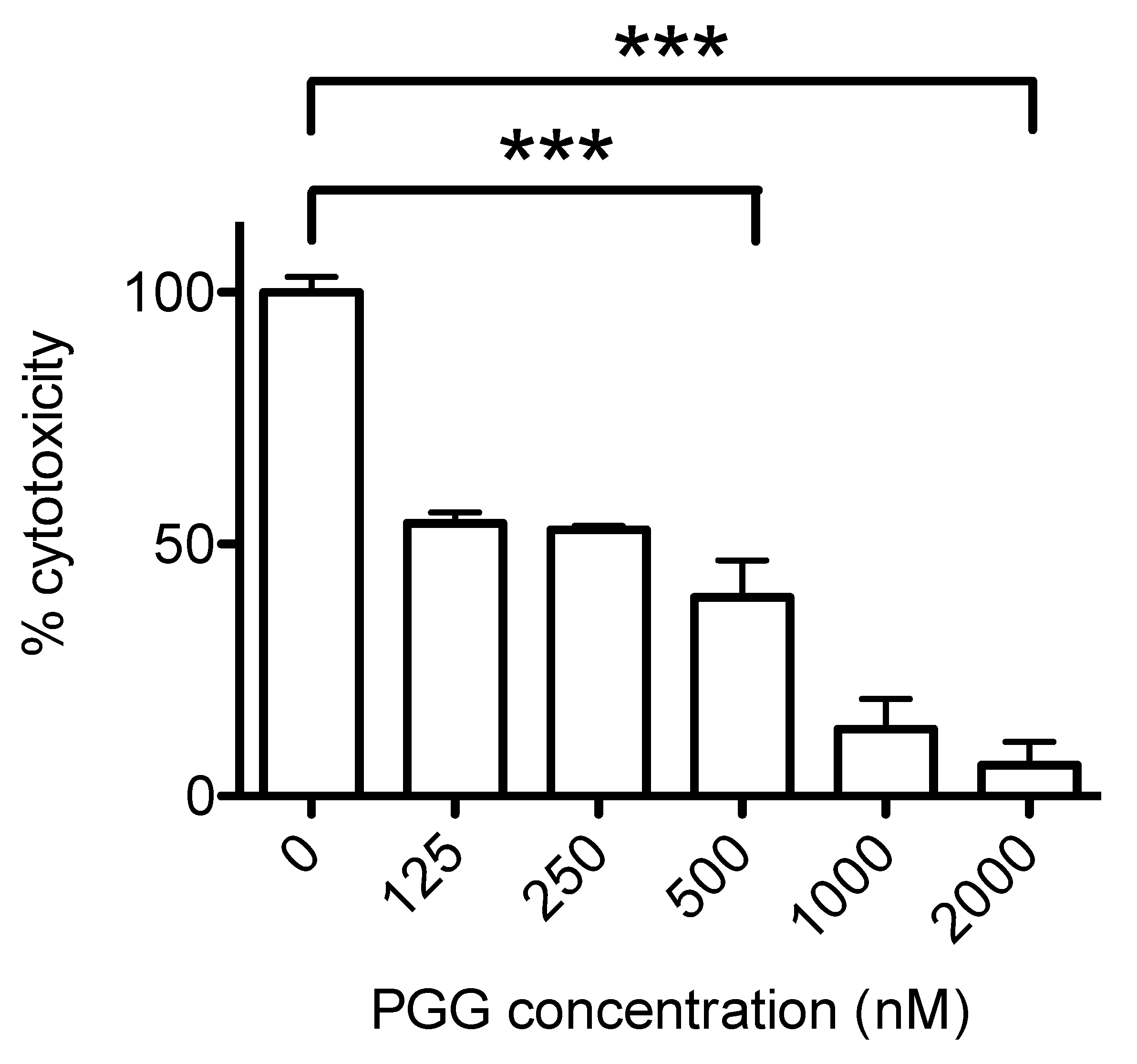
2.4. Inhibition of Ply Cytotoxicity by PGG
2.5. In Silico Studies of PGG Interaction with Ply
3. Discussion
4. Materials and Methods
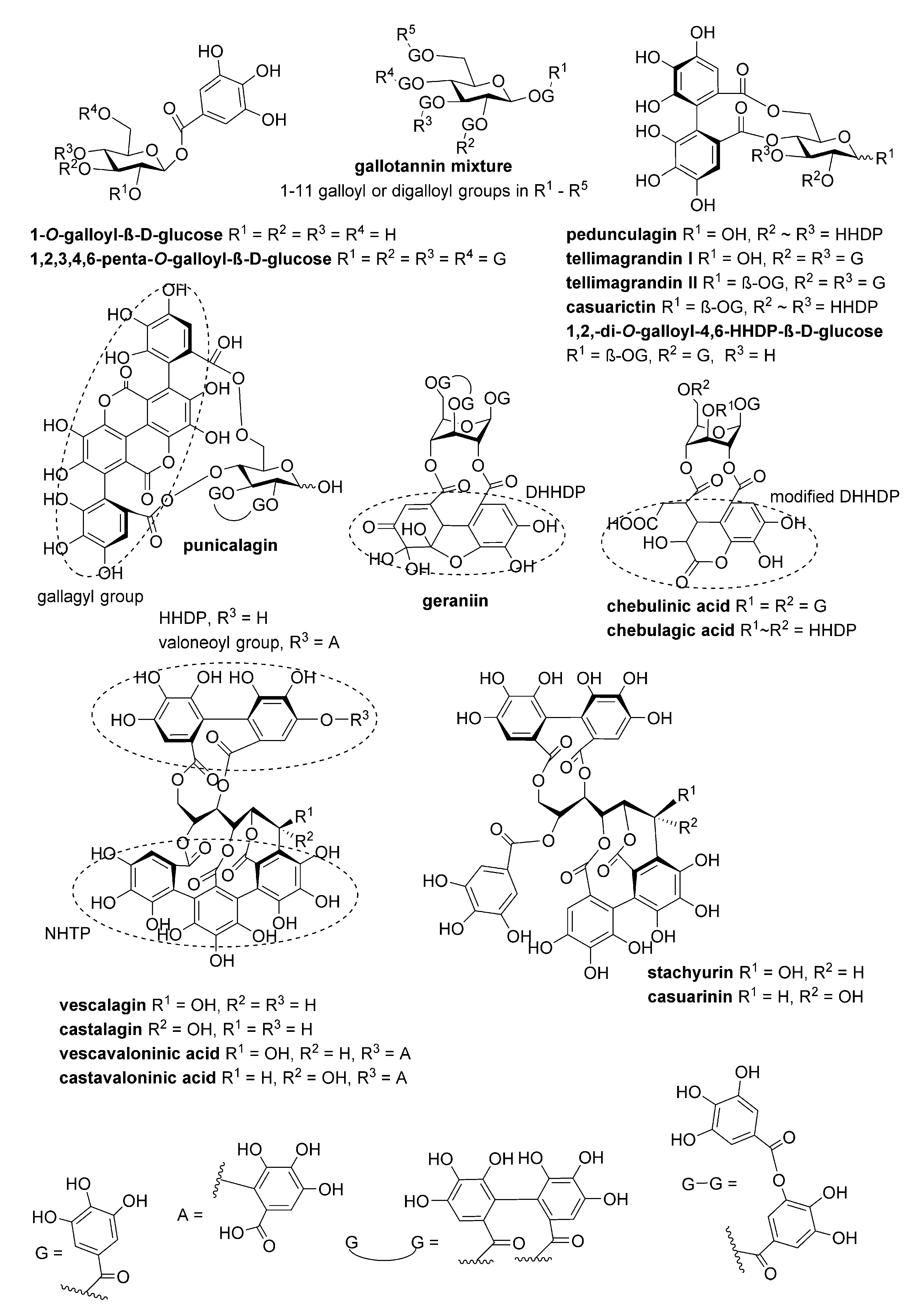
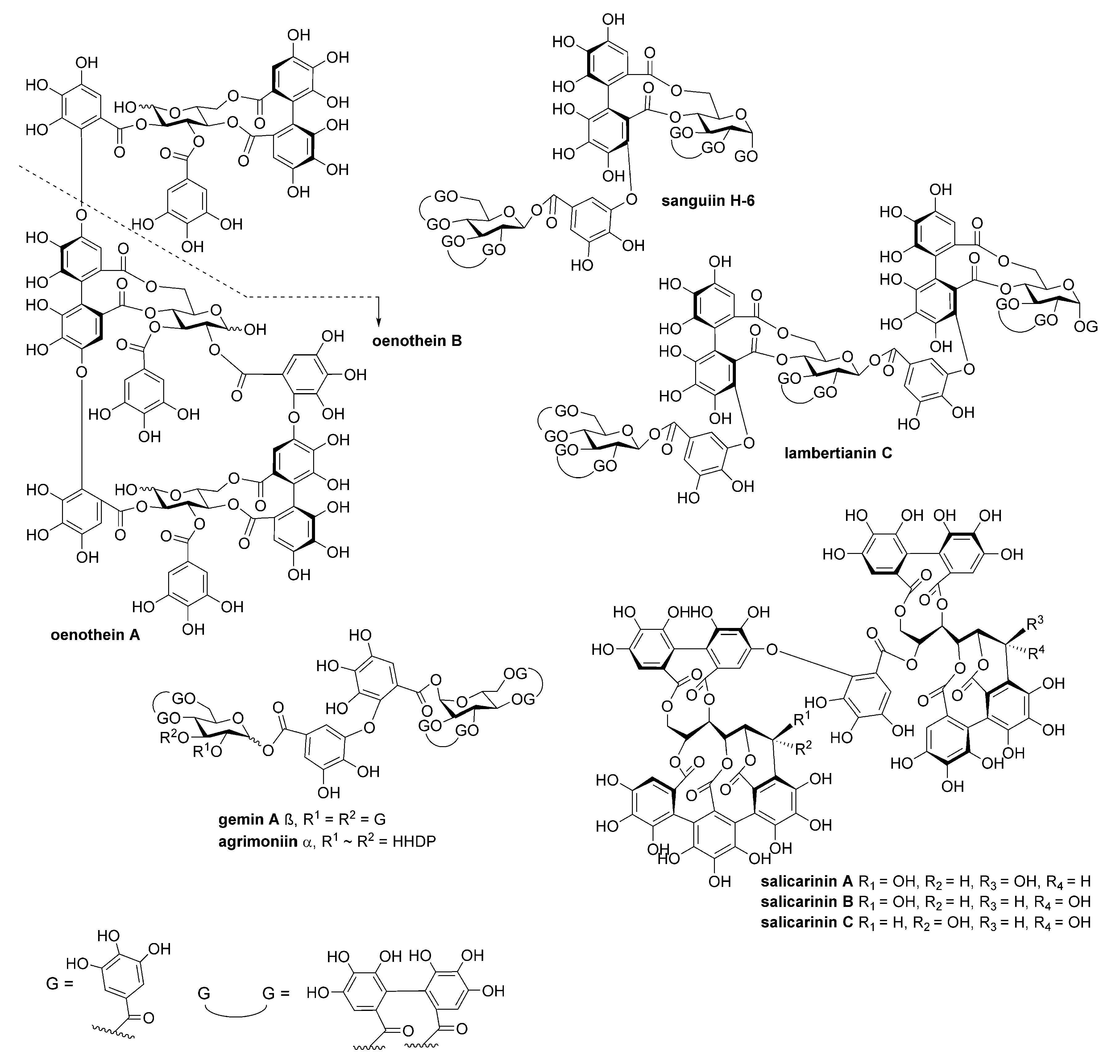
4.1. Tannins
4.2. Cloning and Expression of Recombinant Pneumolysin
4.3. Hemolysis Inhibition Assays with Tannins
4.4. Precipitation Assay
4.5. Liposomes
4.6. Oligomerization Assay
4.7. A549 Cell Cytotoxicity Assay
4.8. Molecular Modelling
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | MW (Da) | IC50 (nM) |
|---|---|---|
| 1,2,3,4,6-penta-O-galloyl-β-D-glucose | 940.67 | 10 |
| Tellimagrandin II | 938.66 | 10 |
| Gemin A | 1873.28 | 20 |
| Lambertianin C | 2805.9 | 20 |
| Sanguiin H-6 | 1871.27 | 20 |
| Agrimoniin | 1871.27 | 30 |
| Gallotannin mixture | 1396.99 * | 40 |
| Casuarictin | 936.64 | 40 |
| Oenothein A | 2353.62 | 40 |
| Salicarinin B | 1869.25 | 50 |
| Oenothein B | 1569.08 | 60 |
| Tellimagrandin I | 786.55 | 60 |
| Punicalagin | 1084.71 | 60 |
| 1,2,-di-O-galloyl-4,6-HHDP-β-D-glucose | 786.55 | 70 |
| Salicarinin A | 1869.25 | 80 |
| Castalagin | 934.63 | 80 |
| Casuarinin | 936.64 | 90 |
| Geraniin | 952.64 | 100 |
| Salicarinin C | 1869.25 | 100 |
| Stachyurin | 936.64 | >100 |
| Pedunculagin | 784.54 | >100 |
| Vescalagin | 934.63 | >100 |
| Castavaloninic acid | 1102.73 | >100 |
| Chebulagic acid | 954.66 | >100 |
| Vescavaloninic acid | 1102.73 | >100 |
| Chebulinic acid | 956.67 | >100 |
| 1-O-galloyl-β-D-glucose | 332.26 | >100 |
References
- Troeger, C.; Blacker, B.; Khalil, I.A.; Rao, P.C.; Cao, J.; Zimsen, S.R.M.; Albertson, S.B.; Deshpande, A.; Farag, T.; Abebe, Z.; et al. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1191–1210. [Google Scholar] [CrossRef] [Green Version]
- Feldman, C.; Anderson, R. The Role of Streptococcus pneumoniae in Community-Acquired Pneumonia. Semin. Respir. Crit. Care Med. 2016, 41, 455–469. [Google Scholar]
- Mehr, S.; Wood, N. Streptococcus pneumoniae—A review of carriage, infection, serotype replacement and vaccination. Paediatr. Respir. Rev. 2012, 13, 258–264. [Google Scholar] [CrossRef]
- Chang, Y.W.; Huang, W.C.; Lin, C.Y.; Wang, W.H.; Hung, L.C.; Chen, Y.H. Tellimagrandin ii, a type of plant polyphenol extracted from trapa bispinosa inhibits antibiotic resistance of drug-resistant Staphylococcus aureus. Int. J. Mol. Sci. 2019, 20, 5790. [Google Scholar] [CrossRef] [Green Version]
- Ogunniyi, A.D.; LeMessurier, K.S.; Graham, R.M.A.; Watt, J.M.; Briles, D.E.; Stroeher, U.H.; Paton, J.C. Contributions of pneumolysin, pneumococcal surface protein A (PspA), and PspC to pathogenicity of Streptococcus pneumoniae D39 in a mouse model. Infect. Immun. 2007, 75, 1843–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, A.M.; Ogunniyi, A.D.; Miller, D.C.; Paton, J.C. Comparative virulence of Streptococcus pneumoniae strains with insertion-duplication, point, and deletion mutations in the pneumolysin gene. Infect. Immun. 1999, 67, 981–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benton, K.A.; Paton, J.C.; Briles, D.E. The hemolytic and complement-activating properties of pneumolysin do not contribute individually to virulence in a pneumococcal bacteremia model. Microb. Pathog. 1997, 23, 201–209. [Google Scholar] [CrossRef]
- Benton, K.A.; Paton, J.C.; Briles, D.E. Differences in virulence for mice among Streptococcus pneumoniae strains of capsular types 2, 3, 4, 5, and 6 are not attributable to differences in pneumolysin production. Infect. Immun. 1997, 65, 1237–1244. [Google Scholar] [CrossRef] [Green Version]
- Orihuela, C.J.; Gao, G.; Francis, K.P.; Yu, J.; Tuomanen, E.I. Tissue-specific contributions of pneumococcal virulence factors to pathogenesis. J. Infect. Dis. 2004, 190, 1661–1669. [Google Scholar] [CrossRef]
- Weiser, J.N.; Ferreira, D.M.; Paton, J.C. Streptococcus pneumoniae: Transmission, colonization and invasion. Nat. Rev. Microbiol. 2018, 16, 355–367. [Google Scholar] [CrossRef]
- Gilbert, R.J.C. Chapter 5: Cholesterol-dependent cytolysins. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2010. [Google Scholar]
- Marshall, J.E.; Faraj, B.H.A.; Gingras, A.R.; Lonnen, R.; Sheikh, M.A.; El-Mezgueldi, M.; Moody, P.C.E.; Andrew, P.W.; Wallis, R. The crystal structure of pneumolysin at 2.0 Å resolution reveals the molecular packing of the pre-pore complex. Sci. Rep. 2015, 5, 13293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guiral, S.; Mitchell, T.J.; Martin, B.; Claverys, J.P. Competence-programmed predation of noncompetent cells in the human pathogen Streptococcus pneumoniae: Genetic requirements. Proc. Natl. Acad. Sci. USA 2005, 102, 8710–8715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandara, M.; Skehel, J.M.; Kadioglu, A.; Collinson, I.; Nobbs, A.H.; Blocker, A.J.; Jenkinson, H.F. The accessory Sec system (SecY2A2) in Streptococcus pneumoniae is involved in export of pneumolysin toxin, adhesion and biofilm formation. Microbes Infect. 2017, 19, 402–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, K.E.; Camilli, A. Pneumolysin localizes to the cell wall of Streptococcus pneumoniae. J. Bacteriol. 2009, 191, 2163–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, R.; Feldman, C. Pneumolysin as a potential therapeutic target in severe pneumococcal disease Introduction: The burden of pneumococcal infection. J. Infect. 2017, 74, 527–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimoto, A.T.; Rosch, J.W.; Tuomanen, E.I. Pneumolysin: Pathogenesis and Therapeutic Target. Front. Microbiol. 2020, 11, 1543. [Google Scholar] [CrossRef] [PubMed]
- Zafar, M.A.; Wang, Y.; Hamaguchi, S.; Weiser, J.N. Host-to-Host Transmission of Streptococcus pneumoniae Is Driven by Its Inflammatory Toxin, Pneumolysin. Cell Host Microbe 2017, 21, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Song, M.; Teng, Z.; Li, M.; Niu, X.; Wang, J.; Deng, X. Epigallocatechin gallate inhibits Streptococcus pneumoniae virulence by simultaneously targeting pneumolysin and sortase A. J. Cell. Mol. Med. 2017, 20, 2586–2598. [Google Scholar] [CrossRef]
- Salminen, J.P.; Karonen, M.; Sinkkonen, J. Chemical ecology of tannins: Recent developments in tannin chemistry reveal new structures and structure-activity patterns. Chem. A Eur. J. 2011, 17, 2806–2816. [Google Scholar] [CrossRef]
- Engström, M.T.; Arvola, J.; Nenonen, S.; Virtanen, V.T.J.; Leppä, M.M.; Tähtinen, P.; Salminen, J.P. Structural Features of Hydrolyzable Tannins Determine Their Ability to Form Insoluble Complexes with Bovine Serum Albumin. J. Agric. Food Chem. 2019, 67, 6798–6808. [Google Scholar] [CrossRef] [PubMed]
- Leppä, M.M.; Karonen, M.; Tähtinen, P.; Engström, M.T.; Salminen, J.P. Isolation of chemically well-defined semipreparative liquid chromatography fractions from complex mixtures of proanthocyanidin oligomers and polymers. J. Chromatogr. A 2018, 1576, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, H.; Wang, J.; Guo, Y.; Liu, B.; Deng, X.; Niu, X. Verbascoside alleviates pneumococcal pneumonia by reducing pneumolysin oligomerss. Mol. Pharmacol. 2016, 89, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Shewell, L.K.; Harvey, R.M.; Higgins, M.A.; Day, C.J.; Hartley-Tassell, L.E.; Chen, A.Y.; Gillen, C.M.; James, D.B.A.; Alonzo, F.; Torres, V.J.; et al. The cholesterol-dependent cytolysins pneumolysin and streptolysin O require binding to red blood cell glycans for hemolytic activity. Proc. Natl. Acad. Sci. USA 2014, 111, E5312–E5320. [Google Scholar] [CrossRef] [Green Version]
- Shewell, L.K.; Day, C.J.; Jen, F.E.C.; Haselhorst, T.; Atack, J.M.; Reijneveld, J.F.; Everest-Dass, A.; James, D.B.A.; Boguslawski, K.M.; Brouwer, S.; et al. All major cholesterol-dependent cytolysins use glycans as cellular receptors. Sci. Adv. 2020, 6, eaaz4926. [Google Scholar] [CrossRef] [PubMed]
- Paton, J.C.; Kim, J.-Y.; Rhee, D.-K.; Briles, D.E.; Pyo, S. Streptococcus pneumoniae induces pyroptosis through the regulation of autophagy in murine microglia. Oncotarget 2015, 6, 44161–44178. [Google Scholar] [CrossRef] [Green Version]
- Jacques, L.C.; Panagiotou, S.; Baltazar, M.; Senghore, M.; Khandaker, S.; Xu, R.; Bricio-Moreno, L.; Yang, M.; Dowson, C.G.; Everett, D.B.; et al. Increased pathogenicity of pneumococcal serotype 1 is driven by rapid autolysis and release of pneumolysin. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wolfmeier, H.; Radecke, J.; Schoenauer, R.; Koeffel, R.; Babiychuk, V.S.; Drücker, P.; Hathaway, L.J.; Mitchell, T.J.; Zuber, B.; Draeger, A.; et al. Active release of pneumolysin prepores and pores by mammalian cells undergoing a Streptococcus pneumoniae attack. Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 2498–2509. [Google Scholar] [CrossRef] [Green Version]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. Br. J. Pharmacol. 2017, 174, 1244–1262. [Google Scholar] [CrossRef] [Green Version]
- Dettweiler, M.; Marquez, L.; Lin, M.; Sweeney-Jones, A.M.; Chhetri, B.K.; Zurawski, D.V.; Kubanek, J.; Quave, C.L. Pentagalloyl glucose from Schinus terebinthifolia inhibits growth of carbapenem-resistant Acinetobacter baumannii. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Madar Johansson, M.; Bélurier, E.; Papageorgiou, A.C.; Sundin, A.P.; Rahkila, J.; Kallonen, T.; Nilsson, U.J.; Maatsola, S.; Nyholm, T.K.M.; Käpylä, J.; et al. The binding mechanism of the virulence factor Streptococcus suis adhesin P subtype to globotetraosylceramide is associated with systemic disease. J. Biol. Chem. 2020, 295, 14305–14324. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.F.; Budin, I.; Szostak, J.W. Vesicle extrusion through polycarbonate track-etched membranes using a hand-held mini-extruder. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 9780124200678. [Google Scholar]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 1. Method and Assessment of Docking Accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Halgren, T.A.; Murphy, R.B.; Friesner, R.A.; Beard, H.S.; Frye, L.L.; Pollard, W.T.; Banks, J.L. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 2. Enrichment Factors in Database Screening. J. Med. Chem. 2004, 47, 1750–1759. [Google Scholar] [CrossRef]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Antechamber, An Accessory Software Package For Molecular Mechanical Calculations. J. Am. Chem. Soc 2001, 222, U403. [Google Scholar]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [Green Version]







| Compound | Hemolysis Inhibition | Minimum Precipitating Concentration | ||
|---|---|---|---|---|
| IC50 [nM] | Inhibitory Power a | (µM) | Relative Power | |
| PGG | 18 ± 0.7 | 13 | 14 | 58 |
| Gemin A | 41 ± 1.0 | 5.9 | 6.2 | 130 |
| Oenothein B | 55 ± 2.1 | 4.4 | 510 | 1.6 |
| Vescalagin | 240 ± 5.3 | 1 | 820 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maatsola, S.; Kurkinen, S.; Engström, M.T.; Nyholm, T.K.M.; Pentikäinen, O.; Salminen, J.-P.; Haataja, S. Inhibition of Pneumolysin Cytotoxicity by Hydrolysable Tannins. Antibiotics 2020, 9, 930. https://doi.org/10.3390/antibiotics9120930
Maatsola S, Kurkinen S, Engström MT, Nyholm TKM, Pentikäinen O, Salminen J-P, Haataja S. Inhibition of Pneumolysin Cytotoxicity by Hydrolysable Tannins. Antibiotics. 2020; 9(12):930. https://doi.org/10.3390/antibiotics9120930
Chicago/Turabian StyleMaatsola, Santeri, Sami Kurkinen, Marica T. Engström, Thomas K. M. Nyholm, Olli Pentikäinen, Juha-Pekka Salminen, and Sauli Haataja. 2020. "Inhibition of Pneumolysin Cytotoxicity by Hydrolysable Tannins" Antibiotics 9, no. 12: 930. https://doi.org/10.3390/antibiotics9120930
APA StyleMaatsola, S., Kurkinen, S., Engström, M. T., Nyholm, T. K. M., Pentikäinen, O., Salminen, J.-P., & Haataja, S. (2020). Inhibition of Pneumolysin Cytotoxicity by Hydrolysable Tannins. Antibiotics, 9(12), 930. https://doi.org/10.3390/antibiotics9120930