Bacterial Biofilm and its Role in the Pathogenesis of Disease



Abstract
:1. Introduction
2. The Auditory System
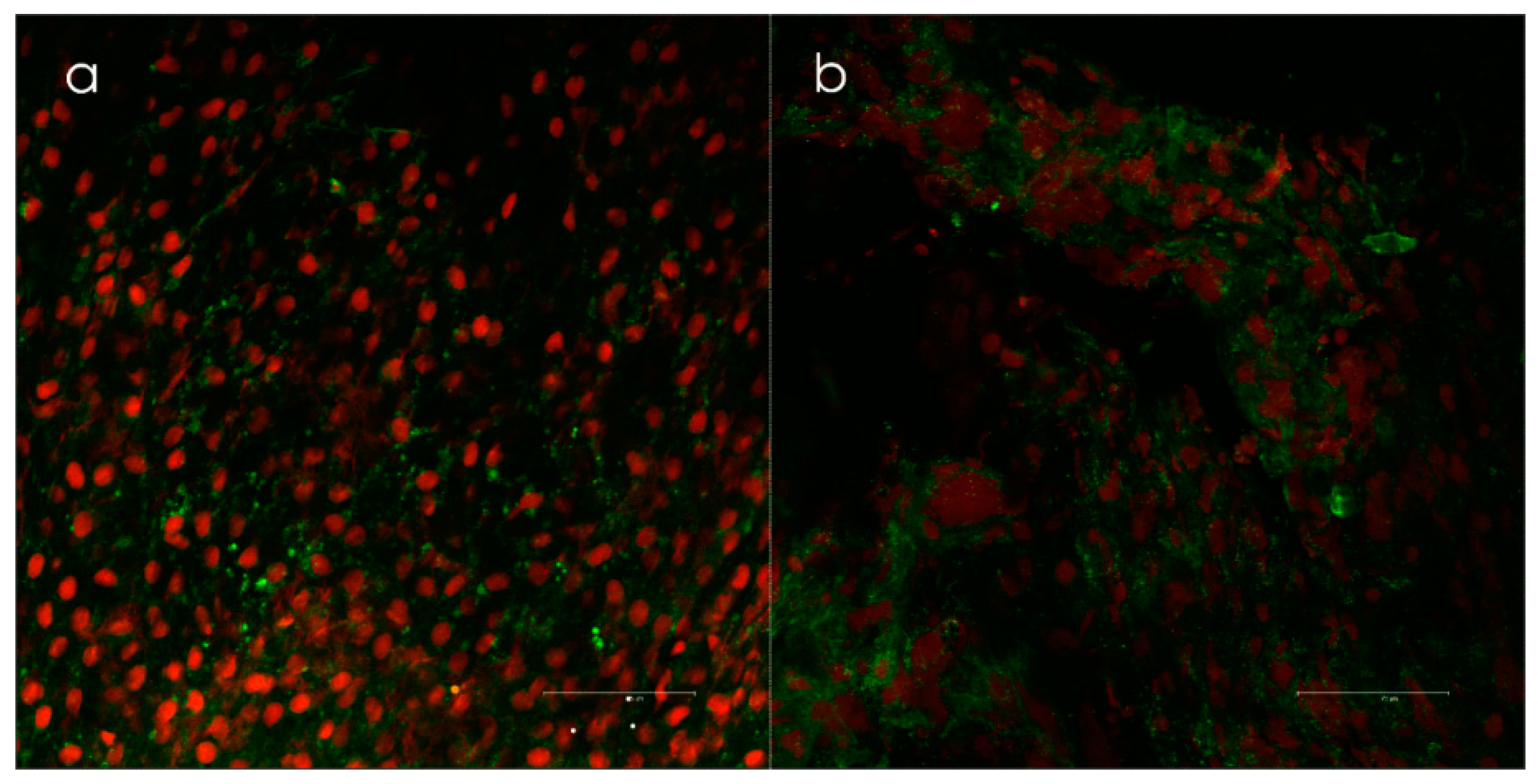
Otitis Media
3. The Cardiovascular System
3.1. Infective Endocarditis (IE)
3.2. Atherosclerosis
4. The Digestive System
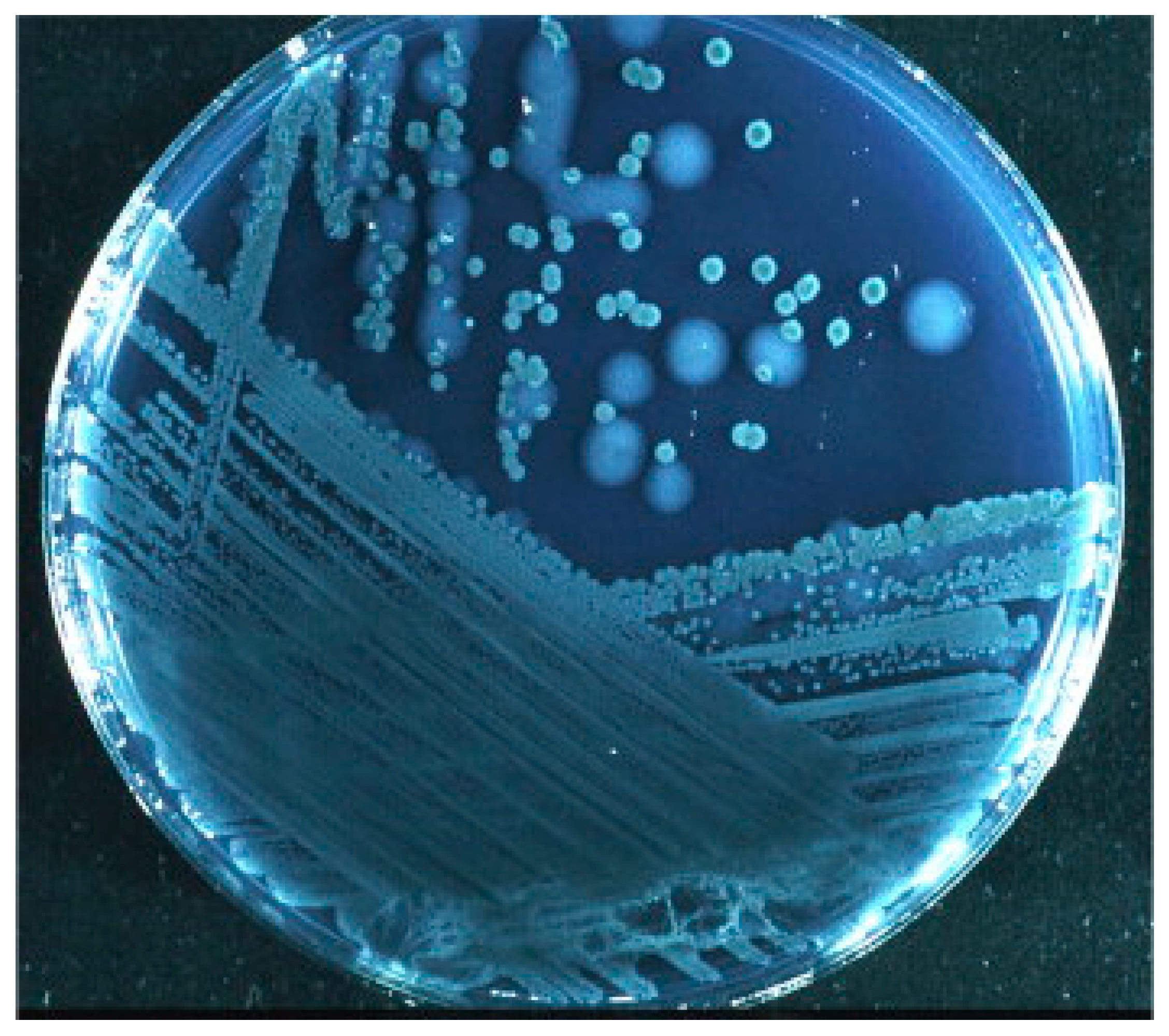
4.1. Sialolithiasis
4.2. Recalcitrant Typhoid Fever and Predisposition to Hepatobiliary Cancers
4.3. Inflammatory Bowel Disease and Colorectal Cancer
5. The Integumentary System
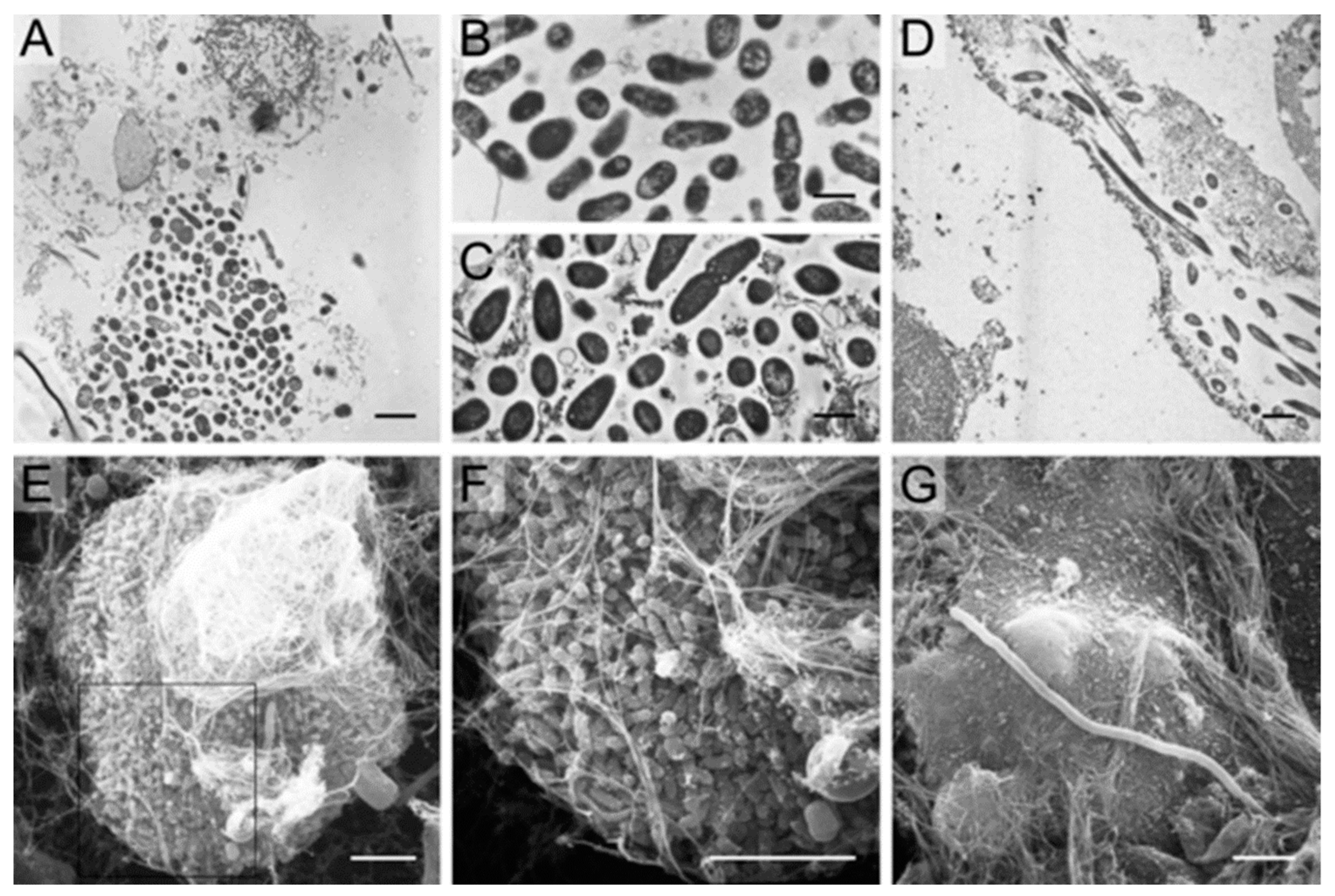
Wound Infections
6. The Reproductive System
6.1. Bacterial Vaginosis
6.2. Chronic Endometritis
6.3. Mastitis
7. The Respiratory System
7.1. Chronic Rhinosinusitis (CRS)
7.2. Pharyngitis and Laryngitis
7.3. Pertussis and other Bordertella Infections
7.4. Cystic Fibrosis (CF)
8. The Urinary System
8.1. Chronic Bacterial Prostatitis
8.2. Urinary Tract Infections (UTI)
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Høiby, N. A short history of microbial biofilms and biofilm infections. APMIS 2017, 125, 272–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoodley, P.; Hall-Stoodley, L. Evolving concepts in biofilm infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T. The role of bacterial biofilms in chronic infections. APMIS 2013, 121, 1–58. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Moser, C.; Pedersen, H.T.; Lerche, C.J.; Kolpen, M.; Line, L.; Thomsen, K.; Høiby, N.; Jensen, P. Østrup Biofilms and host response-helpful or harmful. APMIS 2017, 125, 320–338. [Google Scholar] [CrossRef] [Green Version]
- Auinger, P.; Lanphear, B.P.; Kalkwarf, H.J.; Mansour, M.E. Trends in otitis media among children in the United States. Pediatrics 2003, 112, 514–520. [Google Scholar] [CrossRef] [Green Version]
- DeAntonio, R.; Yarzabal, J.-P.; Cruz, J.P.; Schmidt, J.E.; Kleijnen, J. Epidemiology of otitis media in children from developing countries: A systematic review. Int. J. Pediatr. Otorhinolaryngol. 2016, 85, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Rovers, M.M.; Schilder, A.G.; Zielhuis, A.G.; Rosenfeld, R.M. Otitis media. Lancet 2004, 363, 465–473. [Google Scholar] [CrossRef]
- Schilder, A.G.; Chonmaitree, T.; Cripps, A.W.; Rosenfeld, R.M.; Casselbrant, M.L.; Haggard, M.P.; Venekamp, R.P. Otitis media. Nat. Rev. Dis. Primers 2016. [Google Scholar] [CrossRef] [PubMed]
- Monasta, L.; Ronfani, L.; Marchetti, F.; Montico, M.; Brumatti, L.V.; Bavcar, A.; Grasso, D.; Barbiero, C.; Tamburlini, G. Burden of Disease Caused by Otitis Media: Systematic Review and Global Estimates. PLoS ONE 2012, 7, e36226. [Google Scholar] [CrossRef] [PubMed]
- Chonmaitree, T.; Trujillo, R.; Jennings, K.; Alvarez-Fernandez, P.; Patel, J.A.; Loeffelholz, M.J.; Nokso-Koivisto, J.; Matalon, R.; Pyles, R.B.; Miller, A.L.; et al. Acute Otitis Media and Other Complications of Viral Respiratory Infection. Pediatrics 2016, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faden, H.; Duffy, L.; Wasielewski, R.; Wolf, J.; Krystofik, D.; Tung, Y. Relationship between Nasopharyngeal Colonization and the Development of Otitis Media in Children. J. Infect. Dis. 1997, 175, 1440–1445. [Google Scholar] [CrossRef] [PubMed]
- Giebink, G.S.; Juhn, S.K.; Weber, M.L.; Le, C.T. The bacteriology and cytology of chronic otitis media with effusion. Pediatr. Infect. Dis. J. 1982, 1, 98–103. [Google Scholar] [CrossRef]
- Hotomi, M.; Tabata, T.; Kakiuchi, H.; Kunimoto, M. Detection of Haemophilus influenzae in middle ear of otitis media with effusion by polymerase chain reaction. Int. J. Pediatr. Otorhinolaryngol. 1993, 27, 119–126. [Google Scholar] [CrossRef]
- Post, J.C.; Preston, A.R.; Aul, J.J.; Larkins-Pettigrew, M.; Rydquist-White, J.; Anderson, K.W.; Wadowsky, R.M.; Reagan, D.R.; Walker, E.S.; Kingsley, A.L.; et al. Molecular analysis of bacterial pathogens in otitis media with effusion. JAMA 1995, 273, 1598–1604. [Google Scholar] [CrossRef]
- Fergie, N.; Bayston, R.; Pearson, J.; Birchall, J. Is otitis media with effusion a biofilm infection? Clin. Otolaryngol. 2004, 29, 38–46. [Google Scholar] [CrossRef]
- Ehrlich, G.D.; Veeh, R.; Wang, X.; Costerton, J.W.; Hayes, J.D.; Hu, F.Z.; Daigle, B.J.; Post, J.C. Mucosal Biofilm Formation on Middle-Ear Mucosa in the Chinchilla Model of Otitis Media. JAMA 2002, 287, 1710–1715. [Google Scholar] [CrossRef] [Green Version]
- Hall-Stoodley, L.; Hu, F.Z.; Gieseke, A.; Nistico, L.; Nguyen, D.; Hayes, J.; Forbes, M.; Greenberg, D.P.; Dice, B.; Burrows, A.; et al. Direct detection of bacterial biofilms on the middle-ear mucosa of children with chronic otitis media. JAMA 2006. [Google Scholar] [CrossRef] [Green Version]
- Post, J.C. Candidate’s Thesis: Direct Evidence of Bacterial Biofilms in Otitis Media. Laryngoscope 2001, 111, 2083–2094. [Google Scholar] [CrossRef]
- Daniel, M.; Imtiaz-Umer, S.; Fergie, N.; Birchall, J.; Bayston, R. Bacterial involvement in otitis media with effusion. Int. J. Pediatr. Otorhinolaryngol. 2012, 76, 1416–1422. [Google Scholar] [CrossRef]
- Van Hoecke, H.; De Paepe, A.-S.; Lambert, E.; Van Belleghem, J.D.; Cools, P.; Van Simaey, L.; Deschaght, P.; Vaneechoutte, M.; Dhooge, I. Haemophilus influenzae biofilm formation in chronic otitis media with effusion. Eur. Arch. Oto-Rhino-Laryngol. 2016, 273, 3553–3560. [Google Scholar] [CrossRef] [PubMed]
- Nistico, L.; Kreft, R.; Gieseke, A.; Coticchia, J.M.; Burrows, A.; Khampang, P.; Liu, Y.; Kerschner, J.E.; Post, J.C.; Lonergan, S.; et al. Adenoid Reservoir for Pathogenic Biofilm Bacteria. J. Clin. Microbiol. 2011, 49, 1411–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kania, R.E.; Lamers, G.E.M.; Vonk, M.J.; Dorpmans, E.; Struik, J.; Huy, P.T.; Hiemstra, P.; Bloemberg, G.V.; Grote, J.J. Characterization of Mucosal Biofilms on Human Adenoid Tissues. Laryngoscope 2008, 118, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Coticchia, J.M.; Chen, M.; Sachdeva, L.; Mutchnick, S. New Paradigms in the Pathogenesis of Otitis Media in Children. Front. Pediatr. 2013, 1. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Chang, A.; Xu, Q.; Casey, J.R.; Pichichero, M.E. Phylogenetic relatedness and diversity of non-typable Haemophilus influenzae in the nasopharynx and middle ear fluid of children with acute otitis media. J. Med. Microbiol. 2011, 60, 1841–1848. [Google Scholar] [CrossRef] [Green Version]
- Galli, J.; Calo, L.; Giuliani, M.; Sergi, B.; Lucidi, D.; Meucci, D.; Bassotti, E.; Sanguinetti, M.; Paludetti, G. Biofilm’s role in chronic cholesteatomatous otitis media: A pilot study. Otolaryngol.-Head Neck Surg. 2016. [Google Scholar] [CrossRef]
- Kaya, E.; Dag, I.; Incesulu, A.; Gürbüz, M.K.; Acar, M.; Birdane, L. Investigation of the Presence of Biofilms in Chronic Suppurative Otitis Media, Nonsuppurative Otitis Media, and Chronic Otitis Media with Cholesteatoma by Scanning Electron Microscopy. Sci. World J. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Lampikoski, H.; Aarnisalo, A.A.; Jero, J.; Kinnari, T.J. Mastoid Biofilm in Chronic Otitis Media. Otol. Neurotol. 2012, 33, 785–788. [Google Scholar] [CrossRef] [Green Version]
- Saunders, J.; Murray, M.; Alleman, A. Biofilms in chronic suppurative otitis media and cholesteatoma: Scanning electron microscopy findings. Am. J. Otolaryngol. 2011, 32, 32–37. [Google Scholar] [CrossRef]
- Kuo, C.-L.; Shiao, A.-S.; Yung, M.; Sakagami, M.; Sudhoff, H.; Wang, C.-H.; Hsu, C.-H.; Lien, C.-F. Updates and Knowledge Gaps in Cholesteatoma Research. BioMed Res. Int. 2015, 2015, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Louw, L. Acquired cholesteatoma pathogenesis: Stepwise explanations. J. Laryngol. Otol. 2010, 124, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Hoa, M.; Syamal, M.; Sachdeva, L.; Berk, R.; Coticchia, J. Demonstration of Nasopharyngeal and Middle Ear Mucosal Biofilms in an Animal Model of Acute Otitis Media. Ann. Otol. Rhinol. Laryngol. 2009, 118, 292–298. [Google Scholar] [CrossRef]
- Chole, R.A.; Faddis, B.T. Evidence for microbial biofilms in cholesteatomas. Arch. Otolaryngol.-Head Neck Surg. 2002, 128, 1129–1133. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Keyoumu, Y.; Long, L.; Zhang, H. Detection of bacterial biofilms in different types of chronic otitis media. Eur. Arch. Otorhinolaryngol. 2014. [Google Scholar] [CrossRef]
- Verhoeff, M.; Van Der Veen, E.L.; Rovers, M.M.; Sanders, E.A.; Schilder, A.G. Chronic suppurative otitis media: A review. Int. J. Pediatr. Otorhinolaryngol. 2006, 70, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.; Telischi, F.F.; Eshraghi, A.A.; Yan, D.; Yao, Q.; Lisi, C.V.; Mittal, J.; Narasimhan, G.; Grati, M.; Azad, R.K.; et al. Current concepts in the pathogenesis and treatment of chronic suppurative otitis media. J. Med. Microbiol. 2015, 64, 1103–1116. [Google Scholar] [CrossRef]
- Lee, M.R.; Pawlowski, K.S.; Luong, A.; Furze, A.D.; Roland, P.S. Biofilm presence in humans with chronic suppurative otitis media. Otolaryngol. Neck Surg. 2009, 141, 567–571. [Google Scholar] [CrossRef]
- Belfield, K.; Bayston, R.; Birchall, J.; Daniel, M. Do orally administered antibiotics reach concentrations in the middle ear sufficient to eradicate planktonic and biofilm bacteria? A review. Int. J. Pediatr. Otorhinolaryngol. 2015, 79, 296–300. [Google Scholar] [CrossRef]
- Ah-Tye, C.; Paradise, J.L.; Colborn, D.K. Otorrhea in young children after tympanostomy-tube placement for persistent middle-ear effusion: Prevalence, incidence, and duration. Pediatrics 2001, 107, 1251–1258. [Google Scholar] [CrossRef]
- Rosenfeld, R.M.; Schwartz, S.R.; Pynnonen, M.A.; Tunkel, D.E.; Hussey, H.M.; Fichera, J.S.; Grimes, A.M.; Hackell, J.M.; Harrison, M.F.; Haskell, H.; et al. Clinical practice guideline: Tympanostomy tubes in children. Otolaryngol.-Head Neck Surg. 2013. [Google Scholar] [CrossRef]
- Kay, D.J.; Nelson, M.; Rosenfeld, R.M. Meta-analysis of tympanostomy tube sequelae. Otolaryngol. Neck Surg. 2001, 124, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.J.; Lee, J.C.; Stevens, G.R.; Antonelli, P.J. Opening Plugged Tympanostomy Tubes: Effect of Biofilm Formation. Otolaryngol.-Head Neck Surg. 2006, 134, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Saidi, I.S.; Biedlingmaier, J.F.; Whelan, P. In Vivo Resistance to Bacterial Biofilm Formation on Tympanostomy Tubes as a Function of Tube Material. Otolaryngol.-Head Neck Surg. 1999, 120, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Idicula, W.K.; Jurcisek, J.A.; Cass, N.D.; Ali, S.; Goodman, S.D.; Elmaraghy, C.A.; Jatana, K.R.; Bakaletz, L.O. Identification of biofilms in post-tympanostomy tube otorrhea. Laryngoscope 2016, 126, 1946–1951. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.J.; Eavey, R.D.; Krouse, J.H.; Kiskaddon, R.M. Tympanostomy tubes: Experience with removal. Laryngoscope 1993. [Google Scholar] [CrossRef] [PubMed]
- Dohar, J.; Giles, W.; Roland, P.; Bikhazi, N.; Carroll, S.; Moe, R.; Reese, B.; Dupre, S.; Wall, M.; Stroman, D.; et al. Topical Ciprofloxacin/Dexamethasone Superior to Oral Amoxicillin/Clavulanic Acid in Acute Otitis Media with Otorrhea Through Tympanostomy Tubes. Pediatrics 2006, 118. [Google Scholar] [CrossRef]
- Parsek, M.R.; Singh, P.K. Bacterial Biofilms: An Emerging Link to Disease Pathogenesis. Annu. Rev. Microbiol. 2003, 57, 677–701. [Google Scholar] [CrossRef]
- Elgharably, H.; Hussain, S.T.; Shrestha, N.K.; Blackstone, E.H.; Pettersson, G.B. Current Hypotheses in Cardiac Surgery: Biofilm in Infective Endocarditis. Semin. Thorac. Cardiovasc. Surg. 2016, 28, 56–59. [Google Scholar] [CrossRef]
- Marrie, T.J.; Cooper, J.H.; Costerton, J.W. Ultrastructure of cardiac bacterial vegetations on native valves with emphasis on alterations in bacterial morphology following antibiotic treatment. Can. J. Cardiol. 1987, 3, 275–280. [Google Scholar]
- Long, B.; Koyfman, A. Infectious endocarditis: An update for emergency clinicians. Am. J. Emerg. Med. 2018, 36, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Falk, E.; Shah, P.K.; Fuster, V. Coronary Plaque Disruption. Circulation 1995, 92, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Seymour, G.; Ford, P.; Cullinan, M.; Leishman, S.; Yamazaki, K. Relationship between periodontal infections and systemic disease. Clin. Microbiol. Infect. 2007, 13, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snow, D.E.; Everett, J.; Mayer, G.; Cox, S.B.; Miller, B.; Rumbaugh, K.; Wolcott, R.A.; Wolcott, R.D. The presence of biofilm structures in atherosclerotic plaques of arteries from legs amputated as a complication of diabetic foot ulcers. J. Wound Care 2016, 25, S16–S22. [Google Scholar] [CrossRef] [Green Version]
- Lanter, B.B.; Sauer, K.; Davies, D.G. Bacteria Present in Carotid Arterial Plaques Are Found as Biofilm Deposits Which May Contribute to Enhanced Risk of Plaque Rupture. mBio 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Teymoortash, A.; Wollstein, A.C.; Lippert, B.M.; Peldszus, R.; Werner, J.A. Bacteria and pathogenesis of human salivary calculus. Acta Oto-Laryngol. 2002, 122, 210–214. [Google Scholar] [CrossRef]
- Schrøder, A.S.; Eickhardt, S.; Bjarnsholt, T.; Nørgaard, T.; Homøe, P. Morphological evidence of biofilm in chronic obstructive sialadenitis. J. Laryngol. Otol. 2018, 132, 611–614. [Google Scholar] [CrossRef]
- Kao, W.K.; Chole, R.A.; Ogden, M.A. Evidence of a microbial etiology for sialoliths. Laryngoscope 2019, 130, 69–74. [Google Scholar] [CrossRef]
- Fusconi, M.; Petrozza, V.; Schippa, S.; de Vincentiis, M.; Familiari, G.; Pantanella, F.; Cirenza, M.; Iebba, V.; Battaglione, E.; Greco, A.; et al. Bacterial biofilm in salivary gland stones: Cause or consequence? Otolaryngol.-Head Neck Surg. 2016. [Google Scholar] [CrossRef]
- Perez-Tanoira, R.; Aarnisalo, A.; Haapaniemi, A.; Saarinen, R.; Kuusela, P.; Kinnari, T.J. Bacterial biofilm in salivary stones. Eur. Arch. Oto-Rhino-Laryngol. 2019, 276, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.I.; Bartlett, J.A.; Corey, G.R. Extra-Intestinal Manifestations of Salmonella Infections. Medicine 1987, 66, 349–388. [Google Scholar] [CrossRef] [PubMed]
- Crum, N.F. Current trends in typhoid Fever. Curr. Gastroenterol. Rep. 2003, 5, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Luby, S.P.; Mintz, E.D. The global burden of typhoid fever. Bull. World Heal. Organ. 2004, 82, 346–353. [Google Scholar]
- Lai, C.W.; Chan, R.C.; Cheng, A.F.; Sung, J.Y.; Leung, J.W. Common bile duct stones: A cause of chronic salmonellosis. Am. J. Gastroenterol. 1992, 87, 1198–1199. [Google Scholar] [PubMed]
- Dutta, U.; Garg, P.K.; Kumar, R.; Tandon, R.K. Typhoid carriers among patients with gallstones are at increased risk for carcinoma of the gallbladder. Am. J. Gastroenterol. 2000, 95, 784–787. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.M.; Black, R.E.; Lanata, C. Precise Estimation of the Numbers of Chronic Carriers of Salmonella typhi in Santiago, Chile, an Endemic Area. J. Infect. Dis. 1982, 146, 724–726. [Google Scholar] [CrossRef]
- Gosbell, I.; Jones, P.D.; Matthews, A.; Yeo, B. surgical presentation of hepatobiliary disease due to salmonella typhi. ANZ J. Surg. 1995, 65, 898–899. [Google Scholar] [CrossRef]
- Sinnott, C.; Teall, A. Persistent gallbladder carriage of salmonella typhi. Lancet 1987, 329, 976. [Google Scholar] [CrossRef]
- Vaishnavi, C.; Kochhar, R.; Singh, G.; Kumar, S.; Singh, S.; Singh, K. Epidemiology of typhoid carriers among blood donors and patients with biliary, gastrointestinal and other related diseases. Microbiol. Immunol. 2005, 49, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Dinbar, A.; Altmann, G.; Tulcinsky, D. The treatment of chronic biliary Salmonella carriers. Am. J. Med. 1969, 47, 236–242. [Google Scholar] [CrossRef]
- Gonzalez-Escobedo, G.; Marshall, J.M.; Gunn, J.S. Chronic and acute infection of the gall bladder by salmonella typhi: Understanding the carrier state. Nat. Rev. Microbiol. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Escobedo, G.; Gunn, J.S. Gallbladder Epithelium as a Niche for Chronic Salmonella Carriage. Infect. Immun. 2013, 81, 2920–2930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, R.W.; Rosales-Reyes, R.; Ramírez-Aguilar, M.D.L.L.; Chapa-Azuela, O.; Alpuche-Aranda, C.; Gunn, J.S. Gallstones play a significant role in Salmonella spp. gallbladder colonization and carriage. Proc. Natl. Acad. Sci. USA 2010, 107, 4353–4358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prouty, A.M.; Schwesinger, W.H.; Gunn, J.S. Biofilm Formation and Interaction with the Surfaces of Gallstones by Salmonella spp. Infect. Immun. 2002, 70, 2640–2649. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.M.; Flechtner, A.D.; La Perle, K.M.; Gunn, J.S. Visualization of Extracellular Matrix Components within Sectioned Salmonella Biofilms on the Surface of Human Gallstones. PLoS ONE 2014, 9, e89243. [Google Scholar] [CrossRef] [Green Version]
- González, J.F.; Kurtz, J.; Bauer, D.L.; Hitt, R.; Fitch, J.; Wetzel, A.; La Perle, K.; White, P.; McLachlan, J.; Gunn, J.S. Establishment of Chronic Typhoid Infection in a Mouse Carriage Model Involves a Type 2 Immune Shift and T and B Cell Recruitment to the Gallbladder. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Neiger, M.R.; González, J.F.; Gonzalez-Escobedo, G.; Kuck, H.; White, P.; Gunn, J.S. Pathoadaptive Alteration of Salmonella Biofilm Formation in Response to the Gallbladder Environment. J. Bacteriol. 2019. [Google Scholar] [CrossRef] [Green Version]
- González, J.F.; Tucker, L.; Fitch, J.; Wetzel, A.; White, P.; Gunn, J.S. Human Bile-Mediated Regulation of Salmonella Curli Fimbriae. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [Green Version]
- Dongol, S.; Thompson, C.N.; Clare, S.; Nga, T.V.T.; Duy, P.T.; Karkey, A.; Arjyal, A.; Koirala, S.; Khatri, N.S.; Maskey, P.; et al. The Microbiological and Clinical Characteristics of Invasive Salmonella in Gallbladders from Cholecystectomy Patients in Kathmandu, Nepal. PLoS ONE 2012, 7, e47342. [Google Scholar] [CrossRef] [Green Version]
- Gunn, J.S.; Marshall, J.M.; Baker, S.; Dongol, S.; Charles, R.C.; Ryan, E.T. Salmonella chronic carriage: Epidemiology, diagnosis, and gallbladder persistence. Trends Microbiol. 2014, 22, 648–655. [Google Scholar] [CrossRef] [Green Version]
- González, J.F.; Alberts, H.; Lee, J.; Doolittle, L.; Gunn, J.S. Biofilm Formation Protects Salmonella from the Antibiotic Ciprofloxacin In Vitro and In Vivo in the Mouse Model of chronic Carriage. Sci. Rep. 2018, 8, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo, I.Z.; Quiroz, C.; Gutiérrez, M.A.; Arias, J.; Rentería, M. Fluoroquinolones in the treatment of typhoid fever and the carrier state. Eur. J. Clin. Microbiol. Infect. Dis. 1991, 10, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Ferreccio, C.; Morris, J.G.; Valdivieso, C.; Prenzel, I.; Sotomayor, V.; Drusano, G.L.; Levine, M.M. Efficacy of Ciprofloxacin in the Treatment of Chronic Typhoid Carriers. J. Infect. Dis. 1988, 157, 1235–1239. [Google Scholar] [CrossRef] [PubMed]
- Dejea, C.M.; Sears, C.L. Do biofilms confer a pro-carcinogenic state? Gut Microbes 2016, 7, 54–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Konstantinov, S.R.; Smits, R.; Peppelenbosch, M.P. Bacterial Biofilms in Colorectal Cancer Initiation and Progression. Trends Mol. Med. 2017, 23, 18–30. [Google Scholar] [CrossRef]
- Von Rosenvinge, E.C.; O’May, G.A.; Macfarlane, S.; Macfarlane, G.T.; Shirtliff, M.E. Microbial biofilms and gastrointestinal diseases. Pathog. Dis. 2013, 67, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.R.; Chang, D.K. Colorectal cancer in inflammatory bowel disease: The risk, pathogenesis, prevention and diagnosis. World J. Gastroenterol. 2014, 20, 9872–9881. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Loening-Baucke, V.; Herber, A. Mucosal flora in Crohn’s disease and ulcerative colitis—An overview. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2009, 60, 61–71. [Google Scholar]
- Swidsinski, A.; Weber, J.; Loening-Baucke, V.; Hale, L.P.; Lochs, H. Spatial Organization and Composition of the Mucosal Flora in Patients with Inflammatory Bowel Disease. J. Clin. Microbiol. 2005, 43, 3380–3389. [Google Scholar] [CrossRef] [Green Version]
- Martin, H.M.; Campbell, B.J.; Hart, C.; Mpofu, C.; Nayar, M.; Singh, R.; Englyst, H.; Williams, H.F.; Rhodes, J.M. Enhanced Escherichia coli adherence and invasion in Crohn’s disease and colon cancer 1. Gastroenterol. 2004, 127, 80–93. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in globocan 2012. Int. J. Cancer 2015. [Google Scholar] [CrossRef] [PubMed]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Canha, M.B. Role of colonic microbiota in colorectal carcinogenesis: A systematic review. Rev. Española Enferm. Dig. 2015, 107, 659–671. [Google Scholar]
- Kang, M.; Martin, A. Microbiome and colorectal cancer: Unraveling host-microbiota interactions in colitis-associated colorectal cancer development. Semin. Immunol. 2017, 32, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Tjalsma, H.; Boleij, A.; Marchesi, J.R.; Dutilh, B.E. A bacterial driver–passenger model for colorectal cancer: Beyond the usual suspects. Nat. Rev. Genet. 2012, 10, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Sears, C.L.; Geis, A.L.; Housseau, F. Bacteroides fragilis subverts mucosal biology: From symbiont to colon carcinogenesis. J. Clin. Investig. 2014, 124, 4166–4172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dejea, C.M.; Fathi, P.; Craig, J.M.; Boleij, A.; Taddese, R.; Geis, A.L.; Wu, X.; Shields, C.E.D.; Hechenbleikner, E.M.; Huso, D.L.; et al. Patients with familial adenomatous polyposis harbor colonic biofilms containing tumorigenic bacteria. Science 2018, 359, 592–597. [Google Scholar] [CrossRef] [Green Version]
- Kohoutová, D.; Šmajs, D.; Morávková, P.; Cyrany, J.; Moravkova, M.; Forstlova, M.; Cihak, M.; Rejchrt, S.; Bureš, J. Escherichia coli strains of phylogenetic group B2 and D and bacteriocin production are associated with advanced colorectal neoplasia. BMC Infect. Dis. 2014, 14, 733. [Google Scholar] [CrossRef] [Green Version]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis ofFusobacteriumpersistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef] [Green Version]
- Hold, G.L.; Allen-Vercoe, E. Gut microbial biofilm composition and organisation holds the key to CRC. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 329–330. [Google Scholar] [CrossRef] [PubMed]
- Dejea, C.M.; Wick, E.C.; Hechenbleikner, E.M.; White, J.R.; Welch, J.L.M.; Rossetti, B.J.; Peterson, S.N.; Snesrud, E.C.; Borisy, G.G.; Lazarev, M.; et al. Microbiota organization is a distinct feature of proximal colorectal cancers. Proc. Natl. Acad. Sci. USA 2014, 111, 18321–18326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Castro, E.J.T.; Shim, H.; Advincula, J.V.G.; Kim, Y.-W. Differences Regarding the Molecular Features and Gut Microbiota Between Right and Left Colon Cancer. Ann. Coloproctol. 2018, 34, 280–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomkovich, S.; Dejea, C.M.; Winglee, K.; Drewes, J.L.; Chung, L.; Housseau, F.; Pope, J.L.; Gauthier, J.; Sun, X.; Mühlbauer, M.; et al. Human colon mucosal biofilms from healthy or colon cancer hosts are carcinogenic. J. Clin. Investig. 2019, 130, 1699–1712. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chen, Y.; Fu, X.; Zhou, X.; Peng, Y.; Shi, L.; Chen, T.; Wu, Y.; And, T.C. InvasiveFusobacterium nucleatummay play a role in the carcinogenesis of proximal colon cancer through the serrated neoplasia pathway. Int. J. Cancer 2016, 139, 1318–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fijan, S.; Frauwallner, A.; Langerholc, T.; Krebs, B.; Née Younes, J.A.T.H.; Heschl, A.; Turk, D.M.; Rogelj, I. Efficacy of Using Probiotics with Antagonistic Activity against Pathogens of Wound Infections: An Integrative Review of Literature. BioMed Res. Int. 2019, 2019, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Union of Wound Healing Societies. Principles of Best Practice: Wound Infection in Clinical Practice. An International Consensus; World Union of Wound Healing Societies: London, UK, 2008; Available online: https://www.mepltd.co.uk (accessed on 13 January 2020).
- Percival, S.L.; Hill, K.E.; Williams, D.W.; Hooper, S.J.; Thomas, D.W.; Costerton, J.W. A review of the scientific evidence for biofilms in wounds. Wound Repair Regen. 2012, 20, 647–657. [Google Scholar] [CrossRef]
- Bsc, C.E.D.; Wilson, M.J.; Hill, E.K.; Stephens, P.; Hill, C.M.; Harding, K.G.; Thomas, D.W.; Fds, C.M.H.; Frcs, K.G.H. Use of molecular techniques to study microbial diversity in the skin: Chronic wounds reevaluated. Wound Repair Regen. 2001, 9, 332–340. [Google Scholar]
- Brackman, G.; De Meyer, L.; Nelis, H.; Coenye, T. Biofilm inhibitory and eradicating activity of wound care products againstStaphylococcus aureusandStaphylococcus epidermidisbiofilms in anin vitrochronic wound model. J. Appl. Microbiol. 2013, 114, 1833–1842. [Google Scholar] [CrossRef]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E.D.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in chronic wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- Davis, S.C.; Ricotti, C.; Cazzaniga, A.; Welsh, E.; Eaglstein, W.H.; Mertz, P.M. Microscopic and physiologic evidence for biofilm-associated wound colonization in vivo. Wound Repair Regen. 2008, 16, 23–29. [Google Scholar] [CrossRef]
- Harrison-Balestra, C.; Cazzaniga, A.L.; Davis, S.C.; Mertz, P.M. A wound-isolated pseudomonas aeruginosa grows a biofilm in vitro within 10 h and is visualized by light microscopy. Dermatol. Surg. 2003, 29, 631–635. [Google Scholar] [PubMed]
- Haesler, E.; Swanson, T.; Ousey, K.; Carville, K. Clinical indicators of wound infection and biofilm: Reaching international consensus. J. Wound Care 2019, 28, s4–s12. [Google Scholar] [CrossRef]
- Jung, H.-S.; Ehlers, M.M.; Lombaard, H.; Redelinghuys, M.J.; Kock, M.M. Etiology of bacterial vaginosis and polymicrobial biofilm formation. Crit. Rev. Microbiol. 2017, 210, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, D.; Castro, J.; Palmeira-de-Oliveira, A.; Martinez-de-Oliveira, J.; Cerca, N. Bacterial vaginosis biofilms: Challenges to current therapies and emerging solutions. Front. Microbiol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Amsel, R.; Totten, P.A.; Spiegel, C.A.; Chen, K.C.; Eschenbach, D.; Holmes, K.K. Nonspecific vaginitis. Am. J. Med. 1983, 74, 14–22. [Google Scholar] [CrossRef]
- Swidsinski, A.; Mendling, W.; Loening-Baucke, V.; Ladhoff, A.; Swidsinski, S.; Hale, L.P.; Lochs, H. Adherent Biofilms in Bacterial Vaginosis. Obstet. Gynecol. 2005, 106, 1013–1023. [Google Scholar] [CrossRef] [Green Version]
- Machado, A.; Cerca, N. The influence of biofilm formation by Gardnerella vaginalis and other anaerobes on bacterial vaginosis. J. Infect. Dis. 2015, 212, 1856–1861. [Google Scholar] [CrossRef] [Green Version]
- Verstraelen, H.; Swidsinski, A. The biofilm in bacterial vaginosis: Implications for epidemiology, diagnosis and treatment: 2018 update. Curr. Opin. Infect. Dis. 2019, 32, 38–42. [Google Scholar] [CrossRef]
- Swidsinski, A.; Doerffel, Y.; Loening-Baucke, V.; Swidsinski, S.; Verstraelen, H.; Vaneechoutte, M.; Lemm, V.; Schilling, J.; Mendling, W. Gardnerella Biofilm Involves Females and Males and Is Transmitted Sexually. Gynecol. Obstet. Investig. 2010, 70, 256–263. [Google Scholar] [CrossRef]
- Castro, J.; Alves, P.; Sousa, C.; Cereija, T.; França, Â.; Jefferson, K.K.; Cerca, N. Using an in-vitro biofilm model to assess the virulence potential of Bacterial Vaginosis or non-Bacterial Vaginosis Gardnerella vaginalis isolates. Sci. Rep. 2015, 5, 11640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, J.; França, A.; Bradwell, K.R.; Serrano, M.G.; Jefferson, K.K.; Cerca, N. Comparative transcriptomic analysis of Gardnerella vaginalis biofilms vs. planktonic cultures using RNA-seq. NPJ Biofilm. Microbiomes 2017, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, J.; Machado, D.; Cerca, N. Unveiling the role of Gardnerella vaginalis in polymicrobial Bacterial Vaginosis biofilms: The impact of other vaginal pathogens living as neighbors. ISME J. 2019, 13, 1306–1317. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.; Franasiak, J.M. Endometrial microbiota—New player in town. Fertil. Steril. 2017, 108, 32–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, C.M.; Haick, A.; Nkwopara, E.; Garcia, R.; Rendi, M.; Agnew, K.; Fredricks, D.N.; Eschenbach, D. Colonization of the upper genital tract by vaginal bacterial species in nonpregnant women. Am. J. Obstet. Gynecol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, I.; Codoñer, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazán, J.; Alonso, R.; Alamá, P.; Remohí, J.; Pellicer, A.; et al. Evidence that the endometrial microbiota has an effect on implantation success or failure. Am. J. Obstet. Gynecol. 2016, 215, 684–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cicinelli, E.; De Ziegler, D.; Nicoletti, R.; Tinelli, R.; Saliani, N.; Resta, L.; Bellavia, M.; De Vito, D. Poor Reliability of Vaginal and Endocervical Cultures for Evaluating Microbiology of Endometrial Cavity in Women with Chronic Endometritis. Gynecol. Obstet. Investig. 2009, 68, 108–115. [Google Scholar] [CrossRef]
- Greenwood, S.M.; Moran, J.J. Chronic endometritis: Morphologic and clinical observations. Obstet. Gynecol. 1981, 58, 176–184. [Google Scholar]
- Swidsinski, A.; Verstraelen, H.; Loening-Baucke, V.; Swidsinski, S.; Mendling, W.; Halwani, Z. Presence of a Polymicrobial Endometrial Biofilm in Patients with Bacterial Vaginosis. PLoS ONE 2013, 8, e53997. [Google Scholar] [CrossRef] [Green Version]
- Causey, R.C. Making sense of equine uterine infections: The many faces of physical clearance. Vet. J. 2006, 172, 405–421. [Google Scholar] [CrossRef]
- Riddle, W.; Leblanc, M.; Stromberg, A. Relationships between uterine culture, cytology and pregnancy rates in a Thoroughbred practice. Theriogenology 2007, 68, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Ferris, R.A.; McCue, P.M.; Borlee, G.I.; Loncar, K.D.; Hennet, M.L.; Borlee, B.R. In Vitro Efficacy of Nonantibiotic Treatments on Biofilm Disruption of Gram-Negative Pathogens and an In Vivo Model of Infectious Endometritis Utilizing Isolates from the Equine Uterus. J. Clin. Microbiol. 2016, 54, 631–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferris, R.A.; McCue, P.M.; Borlee, G.I.; Glapa, K.E.; Martin, K.H.; Mangalea, M.R.; Hennet, M.L.; Wolfe, L.M.; Broeckling, C.D.; Borlee, B.R. Model of Chronic Equine Endometritis Involving a Pseudomonas aeruginosa Biofilm. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahanfar, S.; Ng, C.J.; Teng, C.L. Antibiotics for mastitis in breastfeeding women. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef] [Green Version]
- Halasa, T.; Huijps, K.; Østerås, O.; Hogeveen, H. Economic effects of bovine mastitis and mastitis management: A review. Vet. Q. 2007, 29, 18–31. [Google Scholar] [CrossRef]
- Almeida, R.A.; Matthews, K.R.; Cifrian, E.; Guidry, A.J.; Oliver, S.P. Staphylococcus aureus Invasion of Bovine Mammary Epithelial Cells. J. Dairy Sci. 1996, 79, 1021–1026. [Google Scholar] [CrossRef]
- Dego, O.K.; Van Dijk, J.; Nederbragt, H. Factors involved in the early pathogenesis of bovineStaphylococcus aureusmastitis with emphasis on bacterial adhesion and invasion. A review. Vet. Q. 2002, 24, 181–198. [Google Scholar] [CrossRef]
- Hensen, S.M.; Pavicic, M.J.A.M.; Lohuis, J.A.C.M.; de Hoog, J.A.M.; Poutrel, B. Location of staphylococcus aureus within the experimentally infected bovine udder and the expression of capsular polysaccharide type 5 in situ. J. Dairy Sci. 2000, 83, 1966–1975. [Google Scholar] [CrossRef]
- Seixas, R.; Varanda, D.; Bexiga, R.; Tavares, L.; Oliveira, M. Biofilm-formation by Staphylococcus aureus and Staphylococcus epidermidis isolates from subclinical mastitis in conditions mimicking the udder environment. Pol. J. Vet. Sci. 2015, 18, 787–792. [Google Scholar] [CrossRef] [Green Version]
- Xue, T.; Chen, X.; Shang, F. Short communication: Effects of lactose and milk on the expression of biofilm-associated genes in Staphylococcus aureus strains isolated from a dairy cow with mastitis. J. Dairy Sci. 2014, 97, 6129–6134. [Google Scholar] [CrossRef]
- Tremblay, Y.D.; Lamarche, D.; Chever, P.; Haine, D.; Messier, S.; Jacques, M. Characterization of the ability of coagulase-negative staphylococci isolated from the milk of Canadian farms to form biofilms. J. Dairy Sci. 2013, 96, 234–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simojoki, H.; Hyvönen, P.; Ferrer, C.P.; Taponen, S.; Pyorala, S. Is the biofilm formation and slime producing ability of coagulase-negative staphylococci associated with the persistence and severity of intramammary infection? Vet. Microbiol. 2012, 158, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Vautor, E.; Abadie, G.; Pont, A.; Thiéry, R. Evaluation of the presence of the bap gene in Staphylococcus aureus isolates recovered from human and animals species. Vet. Microbiol. 2008, 127, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darwish, S.F.; Asfour, H.A.E. Investigation of Biofilm Forming Ability in Staphylococci Causing Bovine Mastitis Using Phenotypic and Genotypic Assays. Sci. World J. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cucarella, C.; Solano, C.; Valle, J.; Amorena, B.; Lasa, I.; Penades, J.R. Bap, a Staphylococcus aureus Surface Protein Involved in Biofilm Formation. J. Bacteriol. 2001, 183, 2888–2896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuniga, E.; Melville, P.A.; Saidenberg, A.B.; Laes, M.A.; Gonsales, F.F.; Salaberry, S.R.; Gregori, F.; Brandão, P.E.; Dos Santos, F.G.; Lincopan, N.E.; et al. Occurrence of genes coding for MSCRAMM and biofilm-associated protein Bap in Staphylococcus spp. isolated from bovine subclinical mastitis and relationship with somatic cell counts. Microb. Pathog. 2015, 89, 1–6. [Google Scholar] [CrossRef]
- Valle, J.; Latasa, C.; Gil, C.; Toledo-Arana, A.; Solano, C.; Penades, J.R.; Lasa, I. Bap, a Biofilm Matrix Protein of Staphylococcus aureus Prevents Cellular Internalization through Binding to GP96 Host Receptor. PLOS Pathog. 2012, 8, e1002843. [Google Scholar] [CrossRef] [Green Version]
- Gogoi-Tiwari, J.; Williams, V.; Waryah, C.B.; Eto, K.Y.; Tau, M.; Costantino, P.; Tiwari, H.K.; Mukkur, T. Comparative studies of the immunogenicity and protective potential of biofilmvsplanktonicStaphylococcus aureusvaccine against bovine mastitis using non-invasive mouse mastitis as a model system. Biofouling 2015, 31, 543–554. [Google Scholar] [CrossRef]
- Perez, M.M.; Prenafeta, A.; Valle, J.; Penades, J.; Rota, C.; Solano, C.; Marco, J.; Grillo, M.J.; Lasa, I.; Irache, J.M.; et al. Protection from staphylococcus aureus mastitis associated with poly-n-acetyl beta-1,6 glucosamine specific antibody production using biofilm-embedded bacteria. Vaccine 2009. [Google Scholar] [CrossRef] [Green Version]
- Fokkens, W.J.; Lund, V.J.; Mullol, J.; Bachert, C.; Alobid, I.; Baroody, F.; Cohen, N.; Cervin, A.; Douglas, R.; Gevaert, P.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2012. Rhinol. Suppl. 2012, 23, 1–298. [Google Scholar]
- Tan, K.S.; Yan, Y.; Ong, H.H.; Chow, V.T.K.; Shi, L.; Wang, D.-Y. Impact of Respiratory Virus Infections in Exacerbation of Acute and Chronic Rhinosinusitis. Curr. Allergy Asthma Rep. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Bleier, B.S.; Wei, Y. Current Understanding of the Acute Exacerbation of Chronic Rhinosinusitis. Front. Microbiol. 2019, 9, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foreman, A.; Boase, S.; Psaltis, A.; Wormald, P.-J. Role of Bacterial and Fungal Biofilms in Chronic Rhinosinusitis. Curr. Allergy Asthma Rep. 2012, 12, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.J.; Lund, V.J. Biofilms and chronic rhinosinusitis: Systematic review of evidence, current concepts and directions for research. Rhinol. J. 2007, 45, 3–13. [Google Scholar]
- Ramakrishnan, Y.; Shields, R.C.; ElBadawey, M.R.; Wilson, A.J. Biofilms in chronic rhinosinusitis: What is new and where next? J. Laryngol. Otol. 2015, 129, 744–751. [Google Scholar] [CrossRef]
- Danielsen, K.; Eksland, O.; Fridrich-Aas, K.; Orszagh, V.; Bachmann-Harildstad, G.; Burum-Auensen, E. Bacterial biofilms in patients with chronic rhinosinusitis: A confocal scanning laser microscopy study. Rhinol. J. 2014, 52, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Galli, J.; Calò, L.; Ardito, F.; Imperiali, M.; Bassotti, E.; Passali, G.C.; La Torre, G.; Paludetti, G.; Fadda, G. Damage to Ciliated Epithelium in Chronic Rhinosinusitis: What is the Role of Bacterial Biofilms? Ann. Otol. Rhinol. Laryngol. 2008, 117, 902–908. [Google Scholar] [CrossRef]
- Ramadan, H.H.; Sanclement, J.A.; Thomas, J.G. Chronic rhinosinusitis and biofilms. Otolaryngol.-Head Neck Surg. 2005, 132, 414–417. [Google Scholar] [CrossRef]
- You, H.; Zhuge, P.; Li, N.; Shao, L.; Shi, H.; Du, H. Factors affecting bacterial biofilm expression in chronic rhinosinusitis and the influences on prognosis. Am. J. Otolaryngol. 2011, 32, 583–590. [Google Scholar] [CrossRef]
- Wood, A.J.; Fraser, J.; Swift, S.; Amirapu, S.; Douglas, R.G. Are biofilms associated with an inflammatory response in chronic rhinosinusitis? Int. Forum Allergy Rhinol. 2011, 1, 335–339. [Google Scholar] [CrossRef]
- Tan, N.C.-W.; Foreman, A.; Jardeleza, C.; Douglas, R.; Vreugde, S.; Wormald, P.-J. IntracellularStaphylococcus aureus: The Trojan horse of recalcitrant chronic rhinosinusitis? Int. Forum Allergy Rhinol. 2013, 3, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Glowacki, R.; Tomaszewski, K.A.; Strek, P.; Tomaszewska, I.M.; Zgorska-Swierzy, K.; Markiewicz, B.; Skladzien, J. The influence of bacterial biofilm on the clinical outcome of chronic rhinosinusitis: A prospective, double-blind, scanning electron microscopy study. Eur. Arch. Otorhinolaryngol. 2014, 271, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Linkin, D.R.; Finkelman, B.S.; O’Malley, B.W.; Thaler, E.R.; Doghramji, L.; Kennedy, D.W.; Cohen, N.A.; Palmer, J.N. Asthma and biofilm-forming bacteria are independently associated with revision sinus surgeries for chronic rhinosinusitis. J. Allergy Clin. Immunol. 2011, 128, 221–223. [Google Scholar] [CrossRef]
- Psaltis, A.J.; Weitzel, E.K.; Ha, K.R.; Wormald, P.-J. The Effect of Bacterial Biofilms on Post-sinus Surgical Outcomes. Am. J. Rhinol. 2008, 22, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Singhal, D.; Psaltis, A.J.; Foreman, A.; Wormald, P.-J. The Impact of Biofilms on Outcomes after Endoscopic Sinus Surgery. Am. J. Rhinol. Allergy 2010, 24, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Maina, I.W.; Patel, N.N.; Cohen, N.A. Understanding the Role of Biofilms and Superantigens in Chronic Rhinosinusitis. Curr. Otorhinolaryngol. Rep. 2018, 6, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Kadioglu, A.; Weiser, J.N.; Paton, J.C.; Andrew, P.W. The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat. Rev. Genet. 2008, 6, 288–301. [Google Scholar] [CrossRef]
- Gilley, R.P.; Orihuela, C.J. Pneumococci in biofilms are non-invasive: Implications on nasopharyngeal colonization. Front. Microbiol. 2014, 4, 163. [Google Scholar] [CrossRef]
- Marks, L.R.; Davidson, B.A.; Knight, P.R.; Hakansson, A.P. Interkingdom Signaling Induces Streptococcus pneumoniae Biofilm Dispersion and Transition from Asymptomatic Colonization to Disease. mBio 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Zautner, A.E. Adenotonsillar Disease. Recent Pat. Inflamm. Allergy Drug Discov. 2012, 6, 121–129. [Google Scholar] [CrossRef]
- Al-Mazrou, K.A.; Al-Khattaf, A.S. Adherent Biofilms in Adenotonsillar Diseases in Children. Arch. Otolaryngol.-Head Neck Surg. 2008, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chole, R.A.; Faddis, B.T. Anatomical evidence of microbial biofilms in tonsillar tissues: A possible mechanism to explain chronicity. Arch. Otolaryngol.-Head Neck Surg. 2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, R.R.; Picciafuoco, S.; Paraje, M.G.; Villegas, N.A.; Miranda, J.A.; Albesa, I.; Cremonezzi, D.; Commisso, R.; Paglini-Oliva, P. Relevance of biofilms in pediatric tonsillar disease. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.L.; Connolly, K.L.; Kirse, D.J.; Evans, A.K.; Poehling, A.K.; Peters, T.R.; Reid, S.D. Detection of group A Streptococcus in tonsils from pediatric patients reveals high rate of asymptomatic streptococcal carriage. BMC Pediatr. 2012, 12. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.H.; Kim, S.T.; Kang, I.G.; Lee, J.H.; Cha, H.E.; Kim, N.Y. Comparison of tonsillar biofilms between patients with recurrent tonsillitis and a control group. Acta Oto-Laryngol. 2012, 132, 1115–1120. [Google Scholar] [CrossRef]
- Hoa, M.; Tomovic, S.; Nistico, L.; Hall-Stoodley, L.; Stoodley, P.; Sachdeva, L.; Berk, R.; Coticchia, J.M. Identification of adenoid biofilms with middle ear pathogens in otitis-prone children utilizing sem and fish. Int. J. Pediatr. Otorhinolaryngol. 2009, 73, 1242–1248. [Google Scholar] [CrossRef]
- Belcher, R.; Virgin, F. The Role of the Adenoids in Pediatric Chronic Rhinosinusitis. Med. Sci. 2019, 7, 35. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, T.; Riani, C.; Koczan, D.; Standar, K.; Kreikemeyer, B.; Podbielski, A. Protective mechanisms of respiratory tract streptococci against streptococcus pyogenes biofilm formation and epithelial cell infection. Appl. Environ. Microbiol. 2013, 79, 1265–1276. [Google Scholar] [CrossRef] [Green Version]
- Kinnari, T.J. The role of biofilm in chronic laryngitis and in head and neck cancer. Curr. Opin. Otolaryngol. Head Neck Surg. 2015, 23, 448–453. [Google Scholar] [CrossRef]
- Melvin, J.A.; Scheller, E.V.; Miller, J.F.; Cotter, P.A. Bordetella pertussis pathogenesis: Current and future challenges. Nat. Rev. Genet. 2014, 12, 274–288. [Google Scholar] [CrossRef] [Green Version]
- Cattelan, N.; Dubey, P.; Arnal, L.; Yantorno, O.M.; Deora, R. Bordetella biofilms: A lifestyle leading to persistent infections. Pathog. Dis. 2016, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattelan, N.; Jennings-Gee, J.; Dubey, P.; Yantorno, O.M.; Deora, R. Hyperbiofilm Formation by Bordetella pertussis Strains Correlates with Enhanced Virulence Traits. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paddock, C.D.; Sanden, G.N.; Cherry, J.D.; Gal, A.A.; Langston, C.; Tatti, K.M.; Wu, K.-H.; Goldsmith, C.S.; Greer, P.W.; Montague, J.L.; et al. Pathology and Pathogenesis of FatalBordetella pertussisInfection in Infants. Clin. Infect. Dis. 2008, 47, 328–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gouw, D.; Serra, O.D.; De Jonge, I.M.; Hermans, P.W.; Wessels, H.J.; Zomer, A.; Yantorno, O.M.; Diavatopoulos, A.D.; Mooi, F.R. The vaccine potential of Bordetella pertussis biofilm-derived membrane proteins. Emerg. Microbes Infect. 2014, 3. [Google Scholar] [CrossRef]
- Dorji, D.; Graham, R.M.; Singh, A.K.; Ramsay, J.P.; Price, P.; Lee, S. Immunogenicity and protective potential of Bordetella pertussis biofilm and its associated antigens in a murine model. Cell. Immunol. 2019, 337, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Høiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas aeruginosabiofilms in cystic fibrosis. Futur. Microbiol. 2010, 5, 1663–1674. [Google Scholar] [CrossRef]
- Ciofu, O.; Tolker-Nielsen, T.; Jensen, P.Ø.; Wang, H.; Høiby, N. Antimicrobial resistance, respiratory tract infections and role of biofilms in lung infections in cystic fibrosis patients. Adv. Drug Deliv. Rev. 2015, 85, 7–23. [Google Scholar] [CrossRef]
- Hector, A.; Frey, N.; Hartl, M. Update on host-pathogen interactions in cystic fibrosis lung disease. Mol. Cell. Pediatr. 2016, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Starner, T.D.; Zhang, N.; Kim, G.; Apicella, M.A.; McCray, P.B., Jr. Haemophilus influenzae forms biofilms on airway epithelia: Implications in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2006, 174, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Baltimore, R.S.; Christie, C.D.C.; Smith, G.J.W. Immunohistopathologic Localization of Pseudomonas aeruginosa in Lungs from Patients with Cystic Fibrosis: Implications for the Pathogenesis of Progressive Lung Deterioration. Am. Rev. Respir. Dis. 1989, 140, 1650–1661. [Google Scholar] [CrossRef]
- Høiby, N.; Johansen, H.K.; Moser, C.; Song, Z.; Ciofu, O.; Kharazmi, A. Pseudomonas aeruginosa and the in vitro and in vivo biofilm mode of growth. Microbes Infect. 2001, 3, 23–35. [Google Scholar] [CrossRef]
- Lam, J.; Chan, R.; Lam, K.; Costerton, J.W. Production of mucoid microcolonies by Pseudomonas aeruginosa within infected lungs in cystic fibrosis. Infect. Immun. 1980, 28, 546–556. [Google Scholar]
- Singh, P.K.; Schaefer, A.L.; Parsek, M.R.; Moninger, T.O.; Welsh, M.J.; Greenberg, E.P. Quorum-sensing signals indicate that cystic fibrosis lungs are infected with bacterial biofilms. Nature 2000, 407, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Worlitzsch, D.; Tarran, R.; Ulrich, M.; Schwab, U.; Cekici, A.; Meyer, K.C.; Birrer, P.; Bellon, G.; Berger, J.; Weiss, T.; et al. Effects of reduced mucus oxygen concentration in airway Pseudomonas infections of cystic fibrosis patients. J. Clin. Investig. 2002, 109, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.S.; Hennigan, R.F.; Hilliard, G.M.; Ochsner, U.A.; Parvatiyar, K.; Kamani, M.C.; Allen, H.L.; DeKievit, T.R.; Gardner, P.R.; Schwab, U.; et al. Pseudomonas aeruginosa anaerobic respiration in biofilms: Relationships to cystic fibrosis pathogenesis. Dev. Cell 2002, 3, 593–603. [Google Scholar] [CrossRef] [Green Version]
- Høiby, N.; Bjarnsholt, T.; Moser, C.; Bassi, G.; Coenye, T.; Donelli, G.; Hall-Stoodley, L.; Hola, V.; Imbert, C.; Kirketerp-Møller, K.; et al. ESCMID* guideline for the diagnosis and treatment of biofilm infections 2014. Clin. Microbiol. Infect. 2015, 21, S1–S25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, B.I.; Han, D.-S.; Ha, U.-S.; Lee, S.-J.; Sohn, D.W.; Kim, H.W.; Han, C.-H.; Cho, Y.-H. Clinical courses following acute bacterial prostatitis. Prostate Int. 2013, 1, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Yoon, B.I.; Kim, S.; Han, D.-S.; Ha, U.-S.; Lee, S.-J.; Kim, H.W.; Han, C.-H.; Cho, Y.-H. Acute bacterial prostatitis: How to prevent and manage chronic infection? J. Infect. Chemother. 2012, 18, 444–450. [Google Scholar] [CrossRef]
- Wagenlehner, F.M.E.; Diemer, T.; Naber, K.G.; Weidner, W. Chronic bacterial prostatitis (NIH type II): Diagnosis, therapy and influence on the fertility status. Andrologia 2008, 40, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Weidner, W.; Schiefer, H.G.; Jantos, C.; Krauss, H.; Friedrich, H.J.; Altmannsberger, M. Chronic prostatitis: A thorough search for etiologically involved microorganisms in 1,461 patients. Infect. 1991, 19, S119–S125. [Google Scholar] [CrossRef] [PubMed]
- Bartoletti, R.; Cai, T.; Nesi, G.; Albanese, S.; Meacci, F.; Mazzoli, S.; Naber, K. The impact of biofilm-producing bacteria on chronic bacterial prostatitis treatment: Results from a longitudinal cohort study. World J. Urol. 2014, 32, 737–742. [Google Scholar] [CrossRef]
- Kanamaru, S.; Kurazono, H.; Terai, A.; Monden, K.; Kumon, H.; Mizunoe, Y.; Ogawa, O.; Yamamoto, S. Increased biofilm formation in Escherichia coli isolated from acute prostatitis. Int. J. Antimicrob. Agents 2006, 28, 21–25. [Google Scholar] [CrossRef]
- Soto, S.M.; Smithson, A.; Martinez, J.; Horcajada, J.P.; Mensa, J.; Vilá, J. Biofilm Formation in Uropathogenic Escherichia coli Strains: Relationship with Prostatitis, Urovirulence Factors and Antimicrobial Resistance. J. Urol. 2007, 177, 365–368. [Google Scholar] [CrossRef]
- Cai, T.; Tessarolo, F.; Caola, I.; Piccoli, F.; Nollo, G.; Caciagli, P.; Mazzoli, S.; Palmieri, A.; Verze, P.; Malossini, G.; et al. Prostate calcifications: A case series supporting the microbial biofilm theory. Investig. Clin. Urol. 2018, 59, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Nickel, J.C.; Olson, M.E.; Barabas, A.; Benediktsson, H.; Dasgupta, M.K.; Costerton, J. Pathogenesis of Chronic Bacterial Prostatitis in an Animal Model. BJU Int. 1990, 66, 47–54. [Google Scholar] [CrossRef]
- Mazzoli, S. Biofilms in chronic bacterial prostatitis (NIH-II) and in prostatic calcifications. FEMS Immunol. Med. Microbiol. 2010, 59, 337–344. [Google Scholar] [CrossRef]
- Longhi, C.; Comanducci, A.; Riccioli, A.; Ziparo, E.; Marazzato, M.; Aleandri, M.; Conte, A.L.; Lepanto, M.S.; Goldoni, P.; Conte, M.P. Features of uropathogenic Escherichia coli able to invade a prostate cell line. New Microbiol. 2016, 39, 146–149. [Google Scholar]
- Rudick, C.N.; Berry, R.E.; Johnson, J.R.; Johnston, B.; Klumpp, D.J.; Schaeffer, A.J.; Thumbikat, P. Uropathogenic escherichia coli induces chronic pelvic pain. Infect. Immun. 2011, 79, 628–635. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.G.; Palermo, J.J.; Schilling, J.D.; Roth, R.; Heuser, J.; Hultgren, S.J. Intracellular Bacterial Biofilm-Like Pods in Urinary Tract Infections. Science 2003, 301, 105–107. [Google Scholar] [CrossRef] [Green Version]
- Berry, R.E.; Klumpp, D.J.; Schaeffer, A.J. Urothelial Cultures Support Intracellular Bacterial Community Formation by Uropathogenic Escherichia coli. Infect. Immun. 2009, 77, 2762–2772. [Google Scholar] [CrossRef] [Green Version]
- Conte, M.P.; Aleandri, M.; Marazzato, M.; Conte, A.L.; Ambrosi, C.; Nicoletti, M.; Zagaglia, C.; Gambara, G.; Palombi, F.; De Cesaris, P.; et al. The Adherent/Invasive Escherichia coli Strain LF82 Invades and Persists in Human Prostate Cell Line RWPE-1, Activating a Strong Inflammatory Response. Infect. Immun. 2016, 84, 3105–3113. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.-C.; Han, X.-M.; Shi, M.; Pang, Z.-L. Persistence of uropathogenic Escherichia Coli in the bladders of female patients with sterile urine after antibiotic therapies. Acta Acad. Med. Wuhan 2016, 36, 710–715. [Google Scholar] [CrossRef]
- Olson, P.D.; Hunstad, D.A. Subversion of Host Innate Immunity by Uropathogenic Escherichia coli. Pathogens 2016, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Lakeman, M.M.; Roovers, J.-P.W. Urinary tract infections in women with urogynaecological symptoms. Curr. Opin. Infect. Dis. 2016, 29, 1–97. [Google Scholar] [CrossRef]
- Scott, V.C.S.; Haake, D.A.; Churchill, B.M.; Justice, S.S.; Kim, J.-H. Intracellular Bacterial Communities: A Potential Etiology for Chronic Lower Urinary Tract Symptoms. Urology 2015, 86, 425–431. [Google Scholar] [CrossRef] [Green Version]
- Robino, L.; Scavone, P.; Araújo, L.; Algorta, G.; Zunino, P.; Vignoli, R. Detection of intracellular bacterial communities in a child with Escherichia coli recurrent urinary tract infections. Pathog. Dis. 2013, 68, 78–81. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.G.; Dodson, K.W.; Hooton, T.M.; Hultgren, S.J. Intracellular bacterial communities of uropathogenic escherichia coli in urinary tract pathogenesis. Trends Microbiol. 2004, 12, 424–430. [Google Scholar] [CrossRef]
- Anderson, G.G.; Martin, S.M.; Hultgren, S.J. Host subversion by formation of intracellular bacterial communities in the urinary tract. Microbes Infect. 2004, 6, 1094–1101. [Google Scholar] [CrossRef]
- Justice, S.S.; Hung, C.; Theriot, J.A.; Fletcher, D.A.; Anderson, G.G.; Footer, M.J.; Hultgren, S.J. Differentiation and developmental pathways of uropathogenic Escherichia coli in urinary tract pathogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 1333–1338. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, C.K.; Hooton, T.M.; Martin, S.M.; Stamm, W.E.; Palermo, J.J.; Gordon, J.I.; Hultgren, S.J. Escherichia coli from urine of female patients with urinary tract infections is competent for intracellular bacterial community formation. Infect. Immun. 2007, 75, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Rosen, A.D.; Hooton, T.M.; Stamm, E.W.; Humphrey, A.P.; Hultgren, S.J. Detection of Intracellular Bacterial Communities in Human Urinary Tract Infection. PLoS Med. 2007, 4, e329. [Google Scholar] [CrossRef]
- Robino, L.; Scavone, P.; Araujo, L.; Algorta, G.; Zunino, P.; Pírez, M.C.; Vignoli, R. Intracellular Bacteria in the Pathogenesis of Escherichia coli Urinary Tract Infection in Children. Clin. Infect. Dis. 2014, 59, e158–e164. [Google Scholar] [CrossRef] [Green Version]
- Kelley, S.P.; Courtneidge, H.R.; Birch, E.R.; Contreras-Sanz, A.; Kelly, M.C.; Durodie, J.; Peppiatt-Wildman, C.M.; Farmer, C.K.; Delaney, M.P.; Malone-Lee, J.; et al. Urinary ATP and visualization of intracellular bacteria: A superior diagnostic marker for recurrent UTI in renal transplant recipients? Springerplus 2014, 3, 200. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
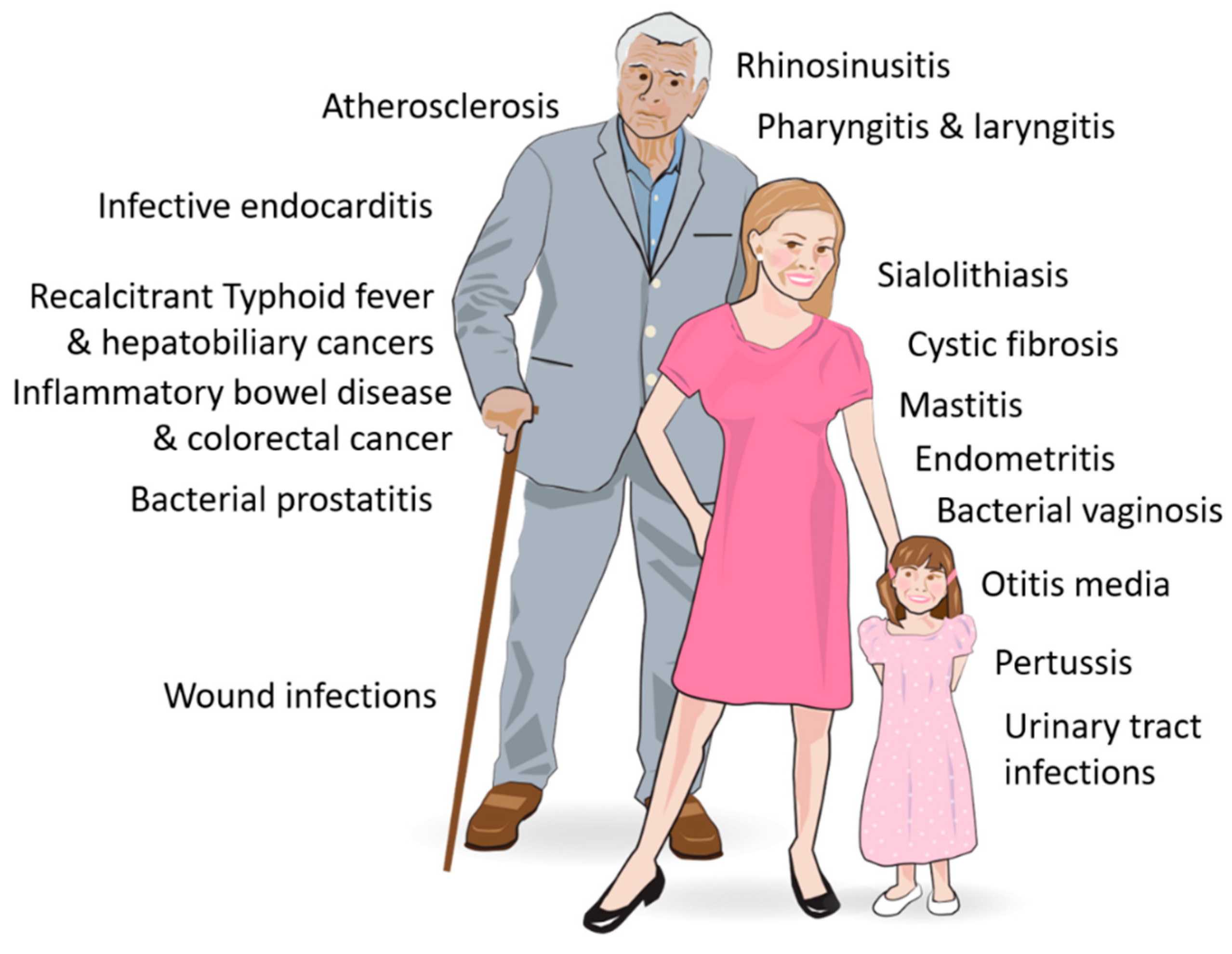
| Body System | Affected Organs | Disease |
|---|---|---|
| Auditory | Middle ear | Otitis media |
| Cardiovascular | Cardiac valves | Infective endocarditis |
| Arteries | Atherosclerosis | |
| Digestive | Salivary glands | Sialolithiasis (salivary duct stones) |
| Gall bladder | Recalcitrant typhoid fever and predisposition to hepatobiliary cancers | |
| Gastrointestinal tract, especially the small and large intestine | Inflammatory bowel disease and colorectal cancer | |
| Integumentary | Skin and underlying tissue | Wound infections |
| Reproductive | Vagina | Bacterial vaginosis |
| Uterus and fallopian tubes | Chronic endometritis | |
| Mammary glands (breasts) | Mastitis | |
| Respiratory | Nasal cavity and paranasal sinuses | Chronic rhinosinusitis |
| Throat, i.e., pharynx with tonsils and adenoids, and larynx with vocal cords | Pharyngitis and laryngitis | |
| Upper and lower airways | Pertussis (whooping cough) and other Bordertella infections | |
| Upper and lower airways | Cystic fibrosis | |
| Urinary | Prostate gland | Chronic bacterial prostatitis |
| Urethra, bladder, urethers, kidneys | Urinary tract infections |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics 2020, 9, 59. https://doi.org/10.3390/antibiotics9020059
Vestby LK, Grønseth T, Simm R, Nesse LL. Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics. 2020; 9(2):59. https://doi.org/10.3390/antibiotics9020059
Chicago/Turabian StyleVestby, Lene K., Torstein Grønseth, Roger Simm, and Live L. Nesse. 2020. "Bacterial Biofilm and its Role in the Pathogenesis of Disease" Antibiotics 9, no. 2: 59. https://doi.org/10.3390/antibiotics9020059
APA StyleVestby, L. K., Grønseth, T., Simm, R., & Nesse, L. L. (2020). Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics, 9(2), 59. https://doi.org/10.3390/antibiotics9020059