New Peptaibiotics and a Cyclodepsipeptide from Ijuhya vitellina: Isolation, Identification, Cytotoxic and Nematicidal Activities

 , ,
, ,  ,
,  and
and
Abstract
:
1. Introduction
2. Results and Discussion
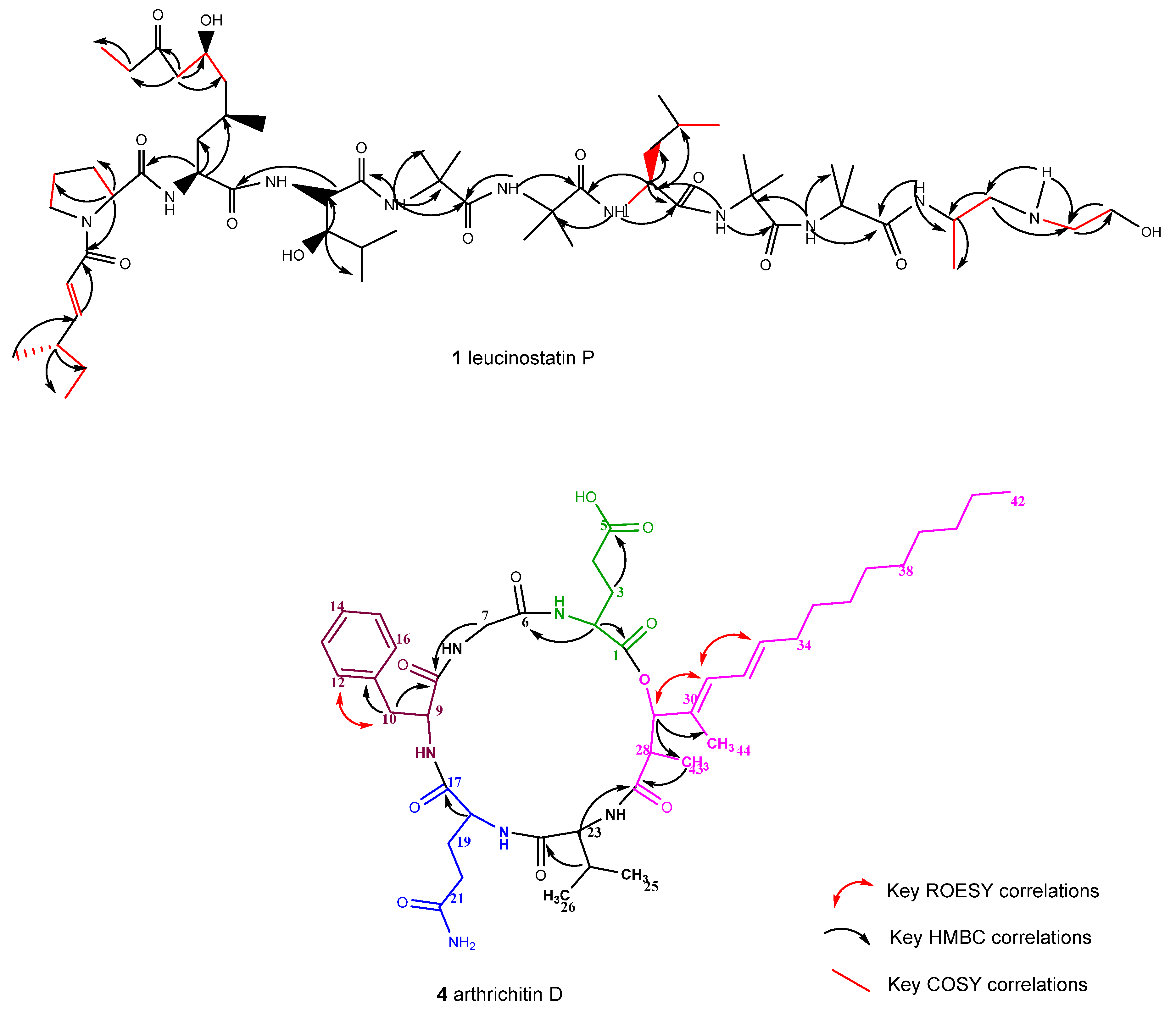
2.1. Isolation and Structure Elucidation of Secondary Metabolites
2.2. Nematicidal Activity of the Metabolites of I. vitellina
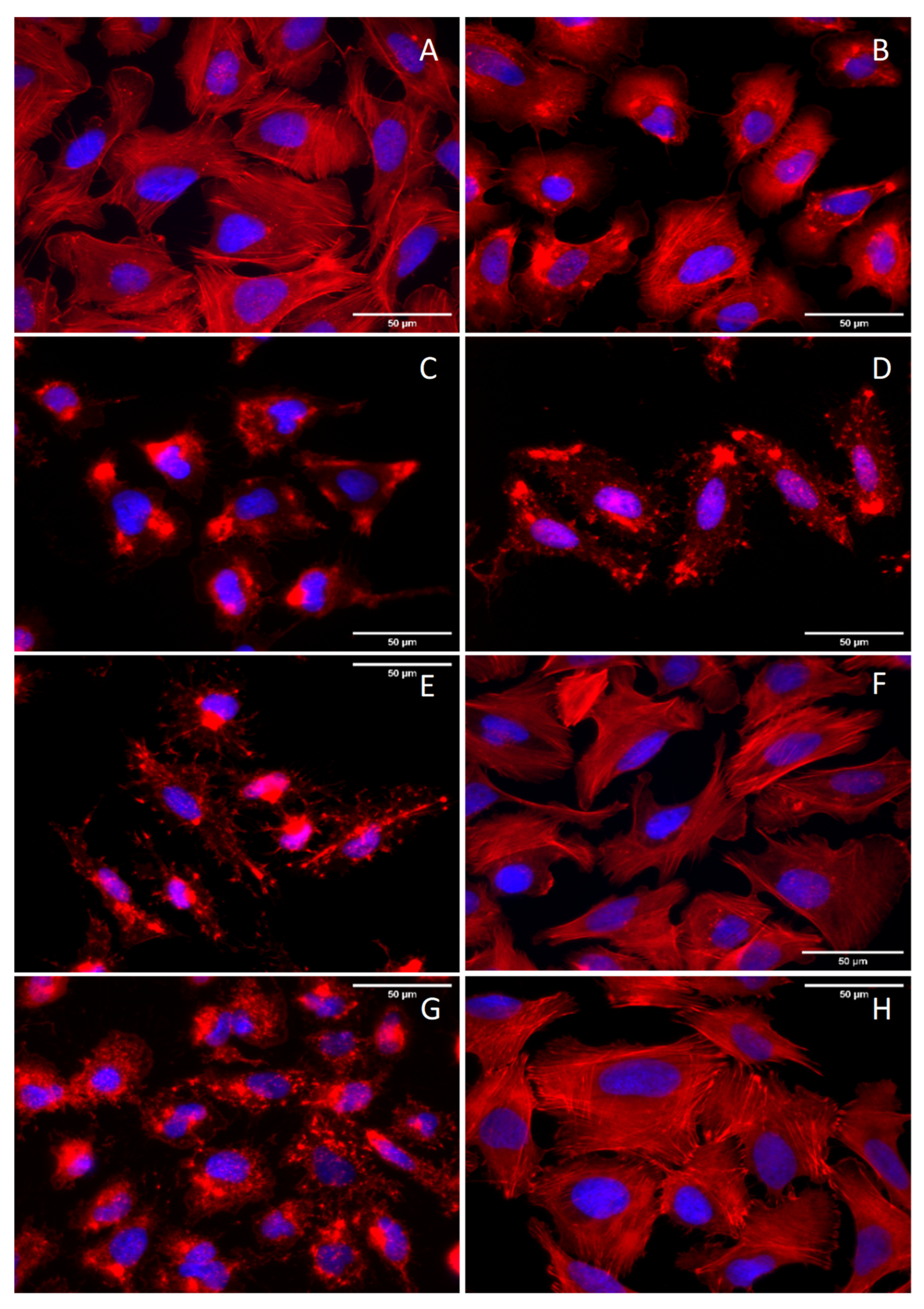
2.3. Assessment of Chaetoglobosin B Activity on Actin
2.4. Conclusion
3. Materials and Methods
3.1. General Information
3.2. Fermentation and Extraction
3.3. Isolation of 1–6
3.4. Marfay’s Analysis
3.5. Mosher Ester Hydrolysis and Absolute Configuration
3.6. Nematicidal Activity
3.7. Evaluation of Chaetoglobosin B for its Effect on Actin in Mammalian Cells
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, G.; Zhang, K.; Xu, J.; Dong, J.; Liu, Y. Nematicidal substances from fungi. Recent Pat. Biotechnol. 2007, 1, 212–233. [Google Scholar] [CrossRef]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.; Gaur, H.S.; Helder, J.; Jones, M.G.; Kikuchi, T.; Manzanilla-Lopez, R.; Palomares-Rius, J.E.; Wesemael, W.M.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2017, 14, 946–961. [Google Scholar] [CrossRef]
- Jones, M.G.K.; Nyarko, J.F. Molecular biology of root lesion nematodes (Pratylenchus spp.) and their interaction with host plants. Ann. Appl. Biol. 2014, 164, 163–181. [Google Scholar] [CrossRef]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Stadler, M.; Chuang, W.; Wu, S.; Ariyawansa, H.A. In vitro inferred interactions of selected entomopathogenic fungi from Taiwan and eggs of Meloidogyne graminicola. Mycol. Prog. 2020, 19, 97–109. [Google Scholar] [CrossRef]
- Degenkolb, T.; Vilcinskas, A. Metabolites from nematophagous fungi and nematicidal natural products from fungi as an alternative for biological control. Part I: Metabolites from nematophagous ascomycetes. Appl. Microbiol. Biotechnol. 2016, 100, 3799–3812. [Google Scholar] [CrossRef]
- Mayer, A.; Anke, H.; Sterner, O. Omphalotin, a new cyclic peptide with potent nematicidal activity from Omphalotus olearius I. Fermentation and biological activity. Nat. Prod. Lett. 1997, 10, 25–32. [Google Scholar] [CrossRef]
- Ashrafi, S.; Stadler, M.; Dababat, A.A.; Richert-Pöggeler, K.R.; Finckh, M.R.; Maier, W. Monocillium gamsii sp. nov. and Monocillium bulbillosum: Two nematode-associated fungi parasitising the eggs of Heterodera filipjevi. MycoKeys 2017, 27, 21–38. [Google Scholar] [CrossRef]
- Ashrafi, S.; Knapp, D.G.; Blaudez, D.; Chalot, M.; Maciá-Vicente, J.G.; Zagyva, I.; Dababat, A.A.; Maier, W.; Kovács, G.M. Inhabiting plant roots, nematodes, and truffles—Polyphilus, a new helotialean genus with two globally distributed species. Mycologia 2018, 110, 286–299. [Google Scholar] [CrossRef]
- Ashrafi, S.; Helaly, S.; Schroers, H.J.; Stadler, M.; Richart-Poeggeler, K.R.; Dababat, K.A.; Maier, W. Ijuhya vitellina sp. nov, a novel source for chaetoglobosin A, is a destructive parasite of the cereal cyst nematode Heterodera filipjevi. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Helaly, S.E.; Ashrafi, S.; Teponno, R.B.; Bernecker, S.; Dababat, A.A.; Maier, W.; Stadler, M. Nematicidal cyclic lipodepsipeptides and a xanthocillin derivative from a phaeosphariaceous fungus parasitizing eggs of the plant parasitic nematode Heterodera filipjevi. J. Nat. Prod. 2018, 81, 2228–2234. [Google Scholar] [CrossRef]
- Momose, I.; Onodera, T.; Doi, H.; Adachi, H.; Iijima, M.; Yamazaki, Y.; Sawa, R.; Kubota, Y.; Igarashi, M.; Kawada, M. Leucinostatin Y: A peptaibiotic produced by the entomoparasitic fungus Purpureocillium lilacinum 40-H-28. J. Nat. Prod. 2019, 82, 1120–1127. [Google Scholar] [CrossRef]
- Vertuani, G.; Boggian, M.; Scatturin, A.; Ricci, M.; Meli Balbocchino, B.; Tuttobello, L.; Rossi, C. Structure activity studies on chemically modified homologues of the antibiotic phytotoxic leucinostatin A. J. Antibiot. 1995, 48, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, E.K.S.; Roy, K.; Chatterjee, S.; Deshmukh, S.K.; Ganguli, B.N.; Fehlhaber, H.W.; Kogler, H. Arthrichitin. A new cell wall active metabolite from Arthrinium phaeospermum. J. Org. Chem. 1996, 61, 6591–6593. [Google Scholar] [CrossRef]
- Arai, T.; Mikami, Y.; Fukushima, K.; Utsumi, T.; Yazawa, K. A new antibiotic, leucinostatin, derived from Penicillium lilacinum. J. Antibiot. 1973, 26, 157–161. [Google Scholar] [CrossRef]
- Harrington, J.M. Antimicrobial peptide killing of African trypanosomes. Parasite Immunol. 2012, 33, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Csermely, P.; Radics, L.; Rossi, C.; Szamel, M.; Ricci, M.; Mihály, K.; Somogyi, J. The nonapeptide leucinostatin A acts as a weak ionophore and as an immunosuppressant on T lymphocytes. Biochim. Biophys. Acta 1994, 1221, 125–132. [Google Scholar] [CrossRef]
- Park, J.O.; Hargreaves, J.R.; Mcconville, E.J.; Stirling, G.R.; Ghisalberti, E.L.; Sivasithamparam, K. Production of leucinostatins and nematicidal activity of Australian isolates of Paecilomyces lilacinus (Thom) Samson. Lett. Appl. Microbiol. 2004, 38, 271–276. [Google Scholar] [CrossRef]
- Degenkolb, T.; Gams, W.; Brückner, H. Natural cyclopeptaibiotics and related cyclic tetrapeptides: Structural diversity and future prospects. Chem. Biodivers. 2008, 5, 693–706. [Google Scholar] [CrossRef]
- Shima, A.; Fukushima, K.; Arai, T.; Terada, H. Dual inhibitory effects of the peptide antibiotics leucinostatins on oxidative phosphorylation in mitochondria since leucinostatin. Cell Struct. Funct. 1990, 15, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Kretz, R.; Wendt, L.; Wongkanoun, S.; Luangsa-Ard, J.J.; Surup, F.; Helaly, S.E.; Noumeur, S.R.; Stadler, M.; Stradal, T.E.B. The effect of cytochalasans on the actin cytoskeleton of eukaryotic cells and preliminary structure–activity relationships. Biomolecules 2019, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Yahara, I.; Harada, F.; Sekita, S.; Yoshihira, K.; Natori, S. Correlation between effects of 24 different cytochalasins on cellular structures and cellular events and those on actin in vitro. J. Cell Biol. 1982, 92, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Scherlach, K.; Boettger, D.; Remme, N.; Hertweck, C. The chemistry and biology of cytochalasans. Nat. Prod. Rep. 2010, 27, 869–886. [Google Scholar] [CrossRef]
- Li, H.; Xiao, J.; Gao, Y.Q.; Tang, J.J.; Zhang, A.L.; Gao, J.M. Chaetoglobosins from Chaetomium globosum, an endophytic fungus in Ginkgo biloba, and their phytotoxic and cytotoxic activities. J. Agric. Food Chem. 2014, 62, 3734–3741. [Google Scholar] [CrossRef]
- Surup, F.; Chauhan, D.; Niggemann, J.; Bartok, E.; Herrmann, J.; Keck, M.; Zander, W.; Stadler, M.; Hornung, V.; Müller, R. Activation of the NLRP3 Inflammasome by hyaboron, a new asymmetric boron-containing macrodiolide from the Myxobacterium Hyalangium minutum. ACS Chem. Biol. 2018, 13, 2981–2988. [Google Scholar] [CrossRef]
- Schlingmann, G.; Milne, L.; Williams, D.R.; Carter, G.T. Cell wall active antifungal compounds produced by the marine fungus Hypoxylon oceanicum LL-15G256 II. J. Antibiot. 1998, 51, 303–316. [Google Scholar] [CrossRef]
- Hoye, T.R.; Jeffrey, C.S.; Shao, F. Mosher ester analysis for the determination of absolute configuration of stereogenic (chiral) carbinol carbons. Nat. Protoc. 2007, 2, 2451–2458. [Google Scholar] [CrossRef]
- Surup, F.; Pommerehne, K.; Schroers, H.J.; Stadler, M. Elsinopirins A–D, decalin polyketides from the ascomycete Elsinoë pyri. Biomolecules 2018, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Elhady, A.; Giné, A.; Topalovic, O.; Jacquiod, S.; Sørensen, S.J.; Sorribas, F.J.; Heuer, H. Microbiomes associated with infective stages of root-knot and lesion nematodes in soil. PLoS ONE 2017, 12, e0177145. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | LD90 [µg/mL] C. elegans | (LD90) µg/mL P. penetrans |
|---|---|---|
| Leucinostatin U (1) | 5 | 100 |
| Leucinostatin Q (2) | 7 | 100 |
| Leucinostatin P (3) | 7 | 100 |
| Arthrichitin D (4) | 100 | >100 |
| 19-O-acetyl-chaetoglobosin B (6) | 25 | >100 |
| Ivermectin (positive control) | 1 | 10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moussa, A.Y.; Lambert, C.; Stradal, T.E.B.; Ashrafi, S.; Maier, W.; Stadler, M.; Helaly, S.E. New Peptaibiotics and a Cyclodepsipeptide from Ijuhya vitellina: Isolation, Identification, Cytotoxic and Nematicidal Activities. Antibiotics 2020, 9, 132. https://doi.org/10.3390/antibiotics9030132
Moussa AY, Lambert C, Stradal TEB, Ashrafi S, Maier W, Stadler M, Helaly SE. New Peptaibiotics and a Cyclodepsipeptide from Ijuhya vitellina: Isolation, Identification, Cytotoxic and Nematicidal Activities. Antibiotics. 2020; 9(3):132. https://doi.org/10.3390/antibiotics9030132
Chicago/Turabian StyleMoussa, Ashaimaa Y., Christopher Lambert, Theresia E.B. Stradal, Samad Ashrafi, Wolfgang Maier, Marc Stadler, and Soleiman E. Helaly. 2020. "New Peptaibiotics and a Cyclodepsipeptide from Ijuhya vitellina: Isolation, Identification, Cytotoxic and Nematicidal Activities" Antibiotics 9, no. 3: 132. https://doi.org/10.3390/antibiotics9030132