Repurposing Disulfiram (Tetraethylthiuram Disulfide) as a Potential Drug Candidate against Borrelia burgdorferi In Vitro and In Vivo

 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Disulfiram Is a Potential Antibiotic on Log Phage and Stationary Cultures of B. burgdorferi B31 MI
2.2. DLS and AFM Imaging Analyses Demonstrate the Formation of Disulfiram Aggregates at High Concentrations
2.3. Disulfiram Treatment Reduces the B. burgdorferi Burden in Tissues Following Dissemination in Infected C3H/HeN Mice
2.4. Disulfiram Treatment Decreases Disease Pathology and Further Reduces Inflammatory Markers in the Heart of B. burgdorferi Infected C3H/HeN Mice
2.5. Disulfiram Treatment Reduces Antibody Titers in the B. burgdorferi Infected Mice
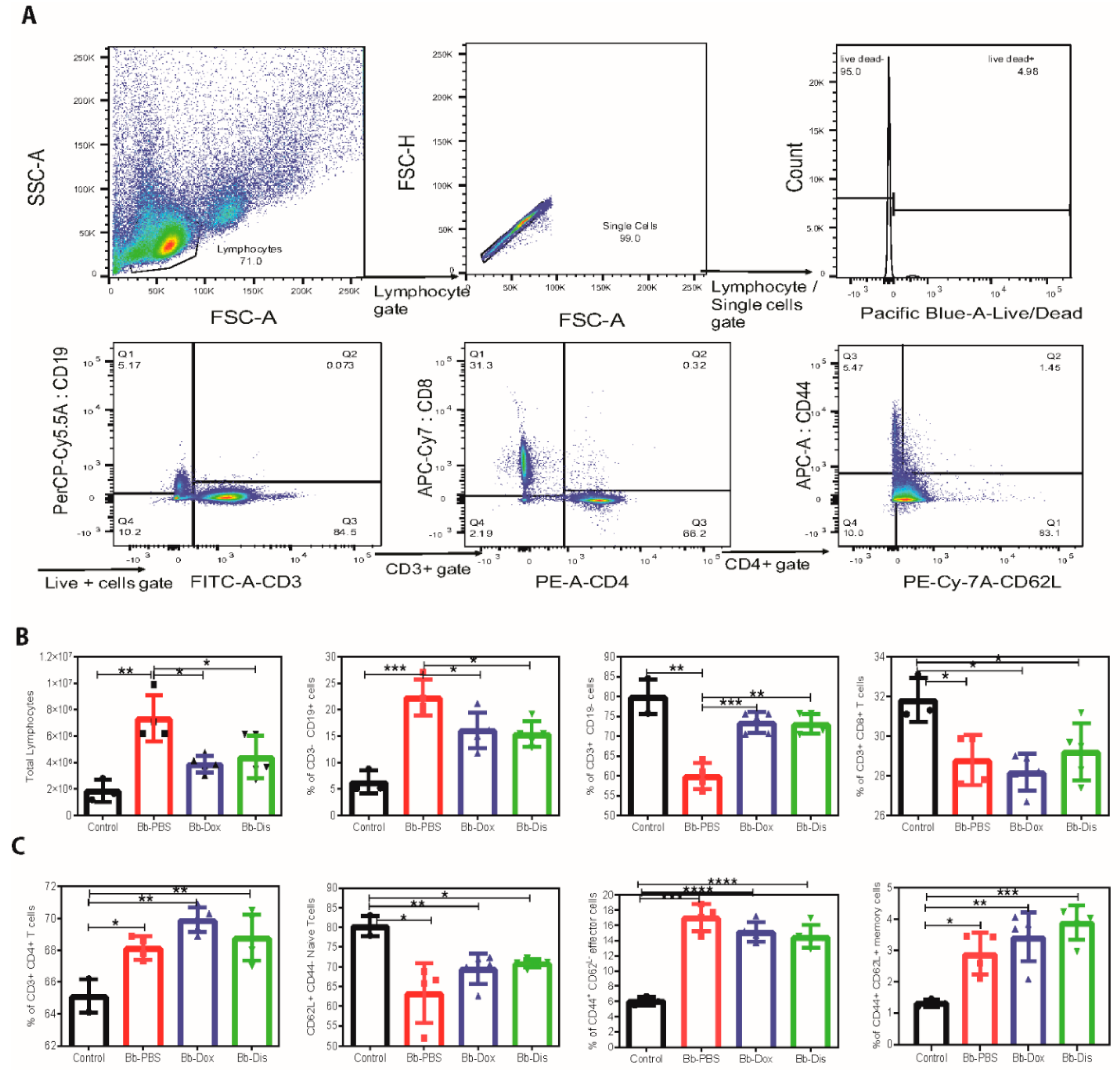
2.6. Disulfiram Reduces Lymphoadenopathy in B. burgdorferi Infected C3H/HeN Mice
3. Discussion
4. Materials and Methods
4.1. Culturing and Growth Conditions of B. burgdorferi B31 MI
4.2. Drug Formulations
4.3. In-Vitro Testing of Antibiotics by Microdilution and Dark Field/Fluorescent Methods
SYBR® Green I/PI by Fluorescent Microscopy
4.4. Dynamic Light Scattering
4.5. Atomic Force Microscopy
4.6. Animal Experiments Ethical Statement
4.7. In Vivo Testing of Drugs in Immunocompetent C3H/HeN Mice
4.8. Quantitative (Q-PCR) and Real-Time PCR (RT-PCR) Analysis
5. Histopathology
5.1. Quantification of Borrelia Specific and Total Immunoglobulins in Serum by ELISA
5.2. Flow Cytometry
6. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- CDC. Tickborne Diseases of The United States A reference Manual for Healthcare Providers; CDC: Atlanta, GA, USA, 2014. [Google Scholar]
- Steere, A.C. Lyme Disease. N. Engl. J. Med. 2001, 345, 115–125. [Google Scholar] [CrossRef] [PubMed]
- STEERE, A.C.; Batsford, W.P.; Weinberg, M.; Alexander, J.; Berger, H.J.; Wolfson, S.; Malawista, S.E. Lyme Carditis: Cardiac Abnormalities of Lyme Disease. Ann. Intern. Med. 1980, 93, 8. [Google Scholar] [CrossRef]
- Nadelman, R.B.; Nowakowski, J.; Fish, D.; Falco, R.C.; Freeman, K.; McKenna, D.; Welch, P.; Marcus, R.; Agüero-Rosenfeld, M.E.; Dennis, D.T.; et al. Prophylaxis with Single-Dose Doxycycline for the Prevention of Lyme Disease after an Ixodes scapularis Tick Bite. N. Engl. J. Med. 2001, 345, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Hodzic, E.; Feng, S.; Holden, K.; Freet, K.J.; Barthold, S.W. Persistence of Borrelia burgdorferi following antibiotic treatment in mice. Antimicrob. Agents Chemother. 2008, 52, 1728–1736. [Google Scholar] [CrossRef]
- Hodzic, E.; Imai, D.; Feng, S.; Barthold, S.W. Resurgence of Persisting Non-Cultivable Borrelia burgdorferi following Antibiotic Treatment in Mice. PLoS ONE 2014, 9, e86907. [Google Scholar] [CrossRef]
- Barthold, S.W.; Hodzic, E.; Imai, D.M.; Feng, S.; Yang, X.; Luft, B.J. Ineffectiveness of tigecycline against persistent Borrelia burgdorferi. Antimicrob. Agents Chemother. 2010, 54, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Straubinger, R.K.; Summers, B.A.; Chang, Y.F.; Appel, M.J. Persistence of Borrelia burgdorferi in experimentally infected dogs after antibiotic treatment. J. Clin. Microbiol. 1997, 35, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-F.; Ku, Y.-W.; Chang, C.-F.; Chang, C.-D.; McDonough, S.P.; Divers, T.; Pough, M.; Torres, A. Antibiotic treatment of experimentally Borrelia burgdorferi-infected ponies. Vet. Microbiol. 2005, 107, 285–294. [Google Scholar] [CrossRef]
- Embers, M.E.; Barthold, S.W.; Borda, J.T.; Bowers, L.; Doyle, L.; Hodzic, E.; Jacobs, M.B.; Hasenkampf, N.R.; Martin, D.S.; Narasimhan, S.; et al. Persistence of Borrelia burgdorferi in Rhesus Macaques following Antibiotic Treatment of Disseminated Infection. PLoS ONE 2012, 7, e29914. [Google Scholar] [CrossRef]
- Crossland, N.A.; Alvarez, X.; Embers, M.E. Late Disseminated Lyme Disease: Associated Pathology and Spirochete Persistence Posttreatment in Rhesus Macaques. Am. J. Pathol. 2018, 188, 672–682. [Google Scholar] [CrossRef]
- Feng, J.; Auwaerter, P.G.; Zhang, Y. Drug Combinations against Borrelia burgdorferi Persisters In Vitro: Eradication Achieved by Using Daptomycin, Cefoperazone and Doxycycline. PLoS ONE 2015, 10, e0117207. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Weitner, M.; Shi, W.; Zhang, S.; Zhang, Y. Eradication of Biofilm-Like Microcolony Structures of Borrelia burgdorferi by Daunomycin and Daptomycin but not Mitomycin C in Combination with Doxycycline and Cefuroxime. Front. Microbiol. 2016, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.R.; Boylan, J.A.; Frye, J.G.; Gherardini, F.C. Evidence of a conjugal erythromycin resistance element in the Lyme disease spirochete Borrelia burgdorferi. Int. J. Antimicrob. Agents 2007, 30, 496–504. [Google Scholar] [CrossRef]
- Pothineni, V.; Wagh, D.; Babar, M.M.; Inayathullah, M.; Solow-Cordero, D.; Kim, K.-M.; Samineni, A.; Parekh, M.B.; Tayebi, L.; Rajadas, J. Identification of new drug candidates against Borrelia burgdorferi using high-throughput screening. Drug Des. Devel. Ther. 2016, 10, 1307. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.P.; Mays, D.C.; Lipsky, J.J. Inhibition of Recombinant Human Mitochondrial and Cytosolic Aldehyde Dehydrogenases by Two Candidates for the Active Metabolites of Disulfiram. Biochemistry 1997, 36, 13748–13754. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Bethesda (MD): National Library of Medicine (US); 14 July 2019. Available online: https://clinicaltrials.gov/ct2/results?cond=&term=disulfiram&cntry=&state=&city=&dist= (accessed on 9 July 2019).
- Long, T.E. Repurposing Thiram and Disulfiram as Antibacterial Agents for Multidrug-Resistant Staphylococcus aureus Infections. Antimicrob. Agents Chemother. 2017, 61, e00898-17. [Google Scholar] [CrossRef]
- Horita, Y.; Takii, T.; Yagi, T.; Ogawa, K.; Fujiwara, N.; Inagaki, E.; Kremer, L.; Sato, Y.; Kuroishi, R.; Lee, Y.; et al. Antitubercular activity of disulfiram, an antialcoholism drug, against multidrug-and extensively drug-resistant Mycobacterium tuberculosis isolates. Antimicrob. Agents Chemother. 2012, 56, 4140–4145. [Google Scholar] [CrossRef]
- Galkin, A.; Kulakova, L.; Lim, K.; Chen, C.Z.; Zheng, W.; Turko, I.V.; Herzberg, O. Structural basis for inactivation of Giardia lamblia carbamate kinase by disulfiram. J. Biol. Chem. 2014, 289, 10502–10509. [Google Scholar] [CrossRef]
- Liegner, K.B. Disulfiram (Tetraethylthiuram Disulfide) in the Treatment of Lyme Disease and Babesiosis: Report of Experience in Three Cases. Antibiotics 2019, 8, 72. [Google Scholar] [CrossRef]
- Boylan, J.A.; Hummel, C.S.; Benoit, S.; Garcia-Lara, J.; Treglown-Downey, J.; Crane, E.J.; Gherardini, F.C. Borrelia burgdorferi bb0728 encodes a coenzyme A disulphide reductase whose function suggests a role in intracellular redox and the oxidative stress response. Mol. Microbiol. 2006, 59, 475–486. [Google Scholar] [CrossRef]
- Parsonage, D.; Desrosiers, D.C.; Hazlett, K.R.O.; Sun, Y.; Nelson, K.J.; Cox, D.L.; Radolf, J.D.; Poole, L.B. Broad specificity AhpC-like peroxiredoxin and its thioredoxin reductant in the sparse antioxidant defense system of Treponema pallidum. Proc. Natl. Acad. Sci. USA 2010, 107, 6240–6245. [Google Scholar] [CrossRef] [PubMed]
- Barth, K.; Malcolm, R. Disulfiram: An Old Therapeutic with New Applications. CNS Neurol. Disord. Drug Targets 2010, 9, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Pothineni, V.R.; Wagh, D.; Inayathullah, M.; Liu, S.; Kim, K.-M.; Rajadas, J. Borreliacidal activity of Borrelia metal transporter A (BmtA) binding small molecules by manganese transport inhibition. Drug Des. Devel. Ther. 2015, 9, 805. [Google Scholar] [CrossRef] [PubMed]
- Pothineni, V.R.; Potula, H.H.S.K.; Ambati, A.; Mallajosyula, V.V.A.; Sridharan, B.; Inayathullah, M.; Ahmed, M.S.; Rajadas, J. Azlocillin can be the potential drug candidate against drug-tolerant Borrelia burgdorferi sensu stricto JLB31. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Barthold, S.W.; Beck, D.S.; Hansen, G.M.; Terwilliger, G.A.; Moody, K.D. Lyme Borreliosis in Selected Strains and Ages of Laboratory Mice. J. Infect. Dis. 1990, 162, 133–138. [Google Scholar] [CrossRef]
- Armstrong, A.L.; Barthold, S.W.; Persing, D.H.; Beck, D.S. Carditis in Lyme Disease Susceptible and Resistant Strains of Laboratory Mice Infected with Borrelia Burgdorferj. Am. J. Trop. Med. Hyg. 1992, 47, 249–258. [Google Scholar] [CrossRef]
- Sigal, L.H. Lyme Disease: A Review of Aspects of Its Immunology and Immunopathogenesis. Annu. Rev. Immunol. 1997, 15, 63–92. [Google Scholar] [CrossRef]
- Tunev, S.S.; Hastey, C.J.; Hodzic, E.; Feng, S.; Barthold, S.W.; Baumgarth, N. Lymphoadenopathy during Lyme Borreliosis Is Caused by Spirochete Migration-Induced Specific B Cell Activation. PLoS Pathog. 2011, 7, e1002066. [Google Scholar] [CrossRef]
- Gourley, T.S.; Wherry, E.J.; Masopust, D.; Ahmed, R. Generation and maintenance of immunological memory. Semin. Immunol. 2004, 16, 323–333. [Google Scholar] [CrossRef]
- Pothineni, V.R.; Wagh, D.; Babar, M.M.; Inayathullah, M.; Watts, R.E.; Kim, K.-M.; Parekh, M.B.; Gurjarpadhye, A.A.; Solow-Cordero, D.; Tayebi, L.; et al. Screening of NCI-DTP library to identify new drug candidates for Borrelia burgdorferi. J. Antibiot. 2017, 70, 308–312. [Google Scholar] [CrossRef]
- Sapi, E.; Kaur, N.; Anyanwu, S.; Luecke, D.F.; Datar, A.; Patel, S.; Rossi, M.; Stricker, R.B. Evaluation of in-vitro antibiotic susceptibility of different morphological forms of Borrelia burgdorferi. Infect. Drug Resist. 2011, 4, 97–113. [Google Scholar] [CrossRef]
- Feng, J.; Wang, T.; Zhang, S.; Shi, W.; Zhang, Y. An Optimized SYBR Green I/PI Assay for Rapid Viability Assessment and Antibiotic Susceptibility Testing for Borrelia burgdorferi. PLoS ONE 2014, 9, 111809. [Google Scholar] [CrossRef]
- Frazier, K.R.; Moore, J.A.; Long, T.E. Antibacterial activity of disulfiram and its metabolites. J. Appl. Microbiol. 2019, 126, 79–86. [Google Scholar] [CrossRef]
- Borlinghaus, J.; Albrecht, F.; Gruhlke, M.; Nwachukwu, I.; Slusarenko, A.; Borlinghaus, J.; Albrecht, F.; Gruhlke, M.C.H.; Nwachukwu, I.D.; Slusarenko, A.J. Allicin: Chemistry and Biological Properties. Molecules 2014, 19, 12591–12618. [Google Scholar] [CrossRef] [PubMed]
- Owen, S.C.; Doak, A.K.; Wassam, P.; Shoichet, M.S.; Shoichet, B.K. Colloidal aggregation affects the efficacy of anticancer drugs in cell culture. ACS Chem. Biol. 2012, 7, 1429–1435. [Google Scholar] [CrossRef]
- Owen, S.C.; Doak, A.K.; Ganesh, A.N.; Nedyalkova, L.; McLaughlin, C.K.; Shoichet, B.K.; Shoichet, M.S. Colloidal drug formulations can explain “bell-shaped” concentration-response curves. ACS Chem. Biol. 2014, 9, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Yip, N.C.; Fombon, I.S.; Liu, P.; Brown, S.; Kannappan, V.; Armesilla, A.L.; Xu, B.; Cassidy, J.; Darling, J.L.; Wang, W. Disulfiram modulated ROS-MAPK and NFB pathways and targeted breast cancer cells with cancer stem cell-like properties. Br. J. Cancer 2011, 104, 1564–1574. [Google Scholar] [CrossRef] [PubMed]
- Wickström, M.; Danielsson, K.; Rickardson, L.; Gullbo, J.; Nygren, P.; Isaksson, A.; Larsson, R.; Lövborg, H. Pharmacological profiling of disulfiram using human tumor cell lines and human tumor cells from patients. Biochem. Pharmacol. 2007, 73, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, S.; Stoye, N.; Luderer, M.; Kiefer, F.; Schmitt, U.; Lieb, K.; Endres, K. Identification of disulfiram as a secretase-modulating compound with beneficial effects on Alzheimer’s disease hallmarks. Sci. Rep. 2018, 8, 1329. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.; Moore, R.D. Disulfiram treatment of alcoholism. Am. J. Med. 1990, 88, 647–655. [Google Scholar] [CrossRef]
- Johansson, B. A review of the pharmacokinetics and pharmacodynamics of disulfiram and its metabolites. Acta Psychiatr. Scand. Suppl. 1992, 369, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.M.; Peter, K. Gessner and Teresa Gessner Disulfiram and Its Metabolite Diethyldithiocarbamate: Pharmacology and Status in the Treatment of Alcoholism, HIV Infection, AIDS and Heavy Metal Toxicity; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Galvao, J.; Davis, B.; Tilley, M.; Normando, E.; Duchen, M.R.; Cordeiro, M.F. Unexpected low-dose toxicity of the universal solvent DMSO. FASEB J. 2014, 28, 1317–1330. [Google Scholar] [CrossRef]
- De Schaepdrijver, L.; Mariën, D.; Rhimi, C.; Voets, M.; van Heerden, M.; Lammens, L. Juvenile animal testing of hydroxypropyl-β-cyclodextrin in support of pediatric drug development. Reprod. Toxicol. 2015, 56, 87–96. [Google Scholar] [CrossRef]
- Li, P.; Song, J.; Ni, X.; Guo, Q.; Wen, H.; Zhou, Q.; Shen, Y.; Huang, Y.; Qiu, P.; Lin, S.; et al. Comparison in toxicity and solubilizing capacity of hydroxypropyl-β-cyclodextrin with different degree of substitution. Int. J. Pharm. 2016, 513, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.K.; Telford Iii, S.R.; Criscione, L.; Lin, S.R.; Spielman, A.; Gravallese, E.M. Cytokines in Murine Lyme Carditis: Th1 Cytokine Expression Follows Expression of Proinflammatory Cytokines in a Susceptible Mouse Strain. J. Infect. Dis. 1998, 177, 242–246. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lasky, C.E.; Olson, R.M.; Brown, C.R. Macrophage Polarization during Murine Lyme Borreliosis. Infect. Immun. 2015, 83, 2627–2635. [Google Scholar] [CrossRef]
- Zhi, H.; Xie, J.; Skare, J.T. The Classical Complement Pathway Is Required to Control Borrelia burgdorferi Levels During Experimental Infection. Front. Immunol. 2018, 9, 959. [Google Scholar] [CrossRef]
- Sprenger, H.; Krause, A.; Kaufmann, A.; Priem, S.; Fabian, D.; Burmester, G.R.; Gemsa, D.; Rittig, M.G. Borrelia burgdorferi Induces Chemokines in Human Monocytes. Infect. Immun. 1997, 65, 4384–4388. [Google Scholar] [CrossRef]
- Davis, M.J.; Tsang, T.M.; Qiu, Y.; Dayrit, J.K.; Freij, J.B.; Huffnagle, G.B.; Olszewski, M.A. Macrophage M1/M2 polarization dynamically adapts to changes in cytokine microenvironments in Cryptococcus neoformans infection. MBio 2013, 4, e00264-13. [Google Scholar] [CrossRef]
- LaRocca, T.J.; Holthausen, D.J.; Hsieh, C.; Renken, C.; Mannella, C.A.; Benach, J.L. The bactericidal effect of a complement-independent antibody is osmolytic and specific to Borrelia. Proc. Natl. Acad. Sci. USA 2009, 106, 10752–10757. [Google Scholar] [CrossRef] [PubMed]
- Katona, L.I.; Ayalew, S.; Coleman, J.L.; Benach, J.L. A bactericidal monoclonal antibody elicits a change in its antigen, OspB of Borrelia burgdorferi, that can be detected by limited proteolysis. J. Immunol. 2000, 164, 1425–3141. [Google Scholar] [CrossRef] [PubMed]
- Hastey, C.J.; Elsner, R.A.; Barthold, S.W.; Baumgarth, N. Delays and diversions mark the development of B cell responses to Borrelia burgdorferi infection. J. Immunol. 2012, 188, 5612–5622. [Google Scholar] [CrossRef]
- Sörman, A.; Zhang, L.; Ding, Z.; Heyman, B. How antibodies use complement to regulate antibody responses. Mol. Immunol. 2014, 61, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Deenick, E.K.; Ma, C.S. The regulation and role of T follicular helper cells in immunity. Immunology 2011, 134, 361–367. [Google Scholar] [CrossRef]
- Dever, L.L.; Jorgensen, J.H.; Barbour, A.G. In Vitro Antimicrobial Susceptibility Testing of Borrelia burgdorferi: A Microdilution MIC Method and Time-Kill Studies. J. Clin. Microbiol. 1992, 30, 2692–2697. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, S.; Shi, W.; Zhang, Y.; Feng, J.; Zhang, S.; Shi, W.; Zhang, Y. Activity of Sulfa Drugs and Their Combinations against Stationary Phase, B. burgdorferi In Vitro. Antibiotics 2017, 6, 10. [Google Scholar] [CrossRef]
- Hyde, J.A.; Weening, E.H.; Skare, J.T. Genetic transformation of Borrelia burgdorferi. Curr. Protoc. Microbiol. 2011, 20, 12C-4. [Google Scholar] [CrossRef]
- Potula, H.-H.; Richer, L.; Werts, C.; Gomes-Solecki, M. Pre-treatment with Lactobacillus plantarum prevents severe pathogenesis in mice infected with Leptospira interrogans and may be associated with recruitment of myeloid cells. PLoS Negl. Trop. Dis. 2017, 11, e0005870. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.R.; Booth, C.J.; Wang, X.; Blaho, V.A.; Malawista, S.E.; Brown, C.R. Recruitment of macrophages and polymorphonuclear leukocytes in Lyme carditis. Infect. Immun. 2007, 75, 613–620. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Mice Infected | Drug Name | Post 21 Days Ear Culture in BSK Medium | No. of fla-b DNA Copies/Ear | No. of fla-b DNA Copies/Bladder | No. of fla-b DNA Copies/Heart |
|---|---|---|---|---|---|
| 1 | Saline (No drug) | + | 847 | 1761 | 15893 |
| 2 | + | 791 | 1422 | 28683 | |
| 3 | + | 7851 | 1335 | 79512 | |
| 4 | + | 1275 | 1136 | 16256 | |
| 5 | + | 1447 | 1933 | 28125 | |
| 1 | Doxycycline | - | 0 | 0 | 842 |
| 2 | - | 4 | 0 | 12 | |
| 3 | - | 0 | 0 | 0 | |
| 4 | - | 56 | 0 | 1325 | |
| 5 | - | 10 | 0 | 0 | |
| 1 | Disulfiram | - | 73 | 0 | 935 |
| 2 | - | 0 | 1439 | 0 | |
| 3 | - | 0 | 0 | 1044 | |
| 4 | - | 20 | 307 | 1297 | |
| 5 | - | 20 | 89 | 0 |
| No. of Mice Infected | Drug Name | Post 21 Days Ear Culture in BSK Medium | No. of fla-b DNA Copies/Ear | No. of fla-b DNA Copies/Bladder | No. of fla-b DNA Copies/Heart |
|---|---|---|---|---|---|
| 1 | Saline (No drug) | + | 477 | 625 | 447 |
| 2 | + | 509 | 469 | 663 | |
| 3 | + | 412 | 837 | 534 | |
| 4 | + | 835 | 448 | 745 | |
| 5 | + | 753 | 331 | 556 | |
| 1 | Doxycycline | - | 28 | 0 | 0 |
| 2 | - | 5 | 0 | 0 | |
| 3 | - | 14 | 6 | 0 | |
| 4 | - | 15 | 0 | 0 | |
| 1 | Disulfiram | - | 0 | 0 | 0 |
| 2 | - | 21 | 0 | 0 | |
| 3 | - | 0 | 0 | 0 | |
| 4 | - | 51 | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potula, H.-H.S.K.; Shahryari, J.; Inayathullah, M.; Malkovskiy, A.V.; Kim, K.-M.; Rajadas, J. Repurposing Disulfiram (Tetraethylthiuram Disulfide) as a Potential Drug Candidate against Borrelia burgdorferi In Vitro and In Vivo. Antibiotics 2020, 9, 633. https://doi.org/10.3390/antibiotics9090633
Potula H-HSK, Shahryari J, Inayathullah M, Malkovskiy AV, Kim K-M, Rajadas J. Repurposing Disulfiram (Tetraethylthiuram Disulfide) as a Potential Drug Candidate against Borrelia burgdorferi In Vitro and In Vivo. Antibiotics. 2020; 9(9):633. https://doi.org/10.3390/antibiotics9090633
Chicago/Turabian StylePotula, Hari-Hara S. K., Jahanbanoo Shahryari, Mohammed Inayathullah, Andrey Victorovich Malkovskiy, Kwang-Min Kim, and Jayakumar Rajadas. 2020. "Repurposing Disulfiram (Tetraethylthiuram Disulfide) as a Potential Drug Candidate against Borrelia burgdorferi In Vitro and In Vivo" Antibiotics 9, no. 9: 633. https://doi.org/10.3390/antibiotics9090633
APA StylePotula, H.-H. S. K., Shahryari, J., Inayathullah, M., Malkovskiy, A. V., Kim, K.-M., & Rajadas, J. (2020). Repurposing Disulfiram (Tetraethylthiuram Disulfide) as a Potential Drug Candidate against Borrelia burgdorferi In Vitro and In Vivo. Antibiotics, 9(9), 633. https://doi.org/10.3390/antibiotics9090633