
The Effect of Different Feeding Applications on the Swimming Behaviour of Siberian Sturgeon: A Method for Improving Restocking Programmes

 , , , ,
, , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Acquisition and Husbandry
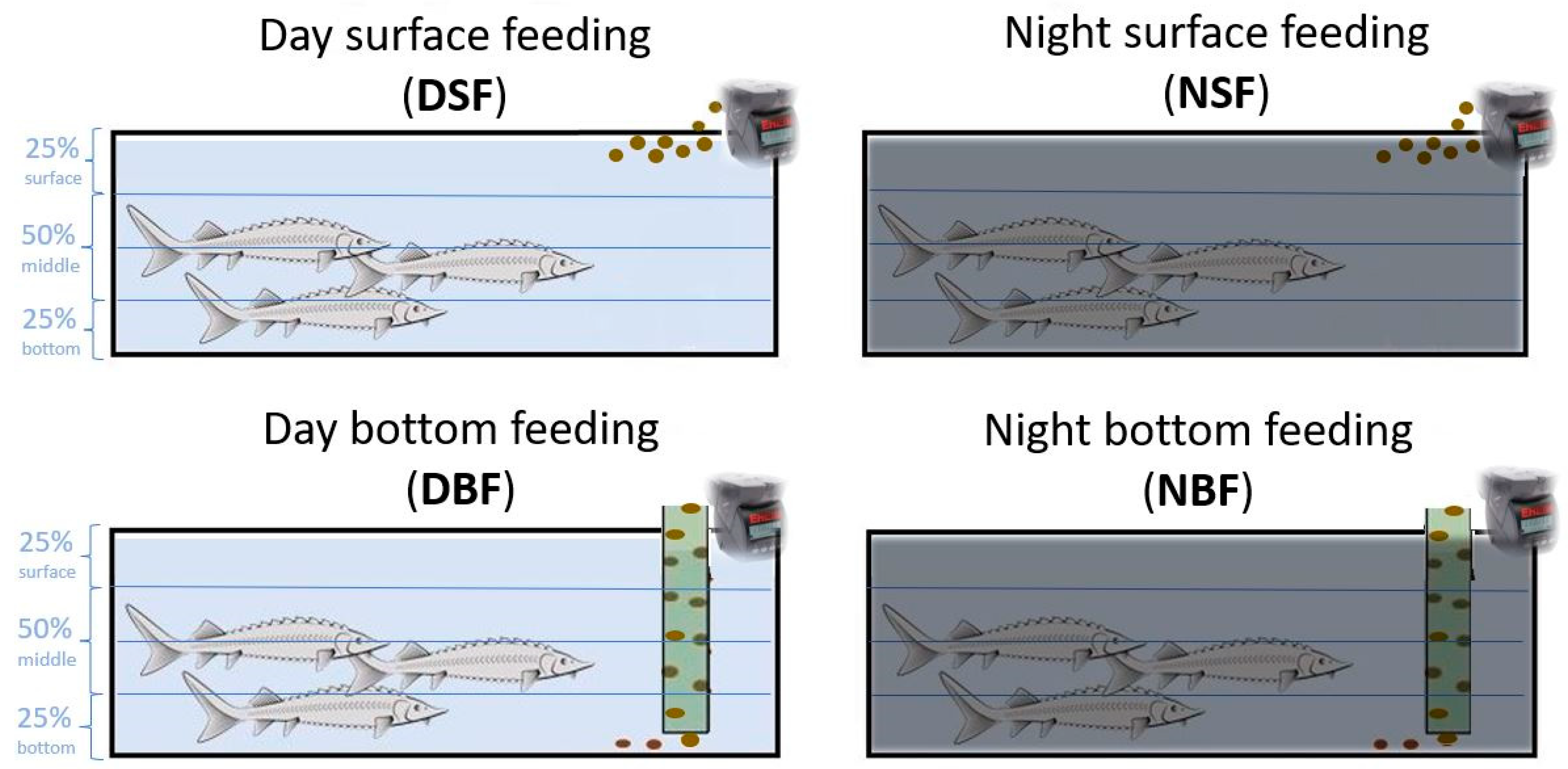
2.2. Feeding Regimes Adaptation
2.3. Video Recording of Swimming Behaviour
2.4. Effect of Starvation on Swimming Behaviour after Different Feeding Applications
2.5. Statistical Analysis
3. Results
3.1. Effect of Surface- and Bottom-Feeding Applications with Day and Night Regimes on Sturgeons Swimming Behaviour
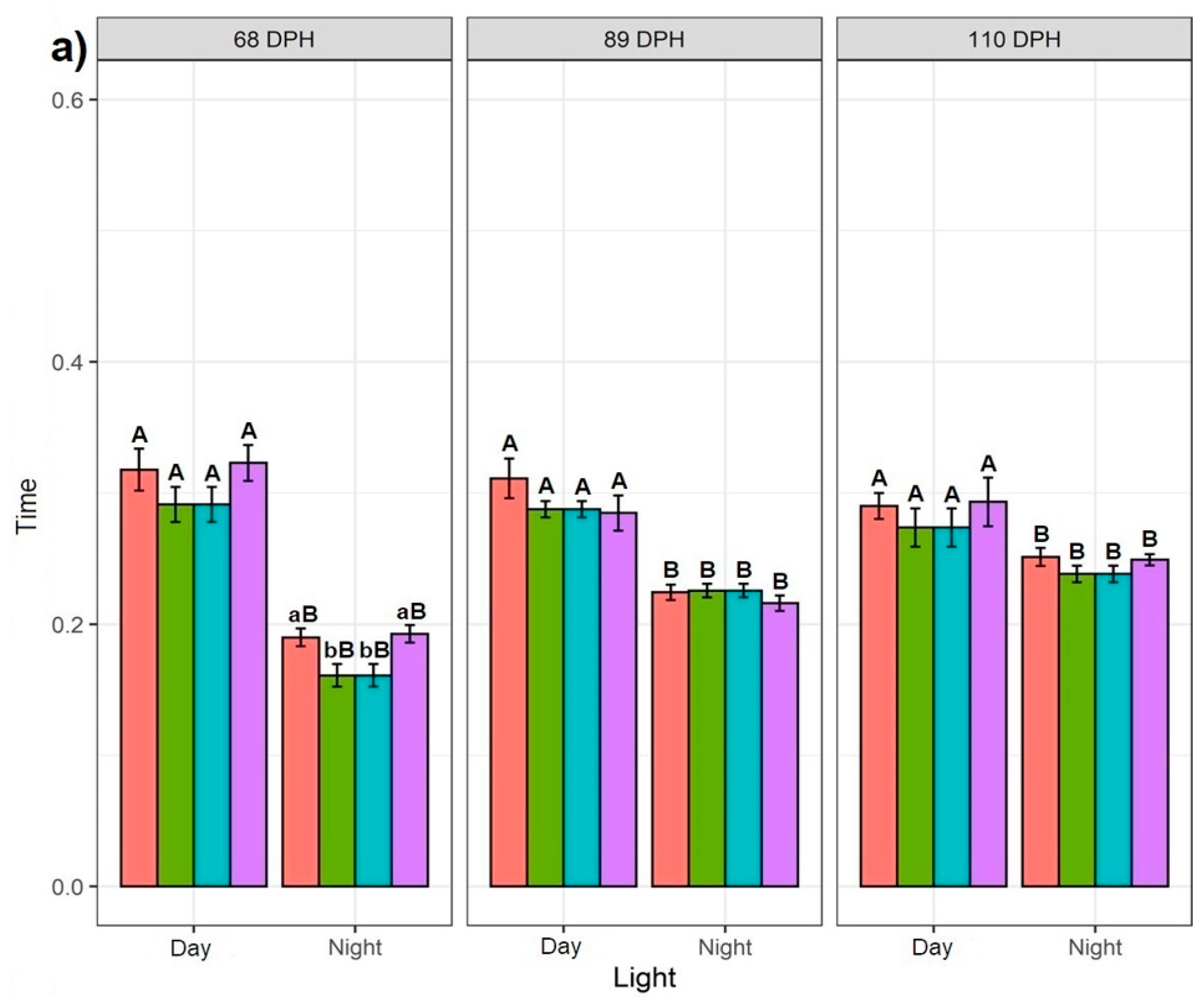
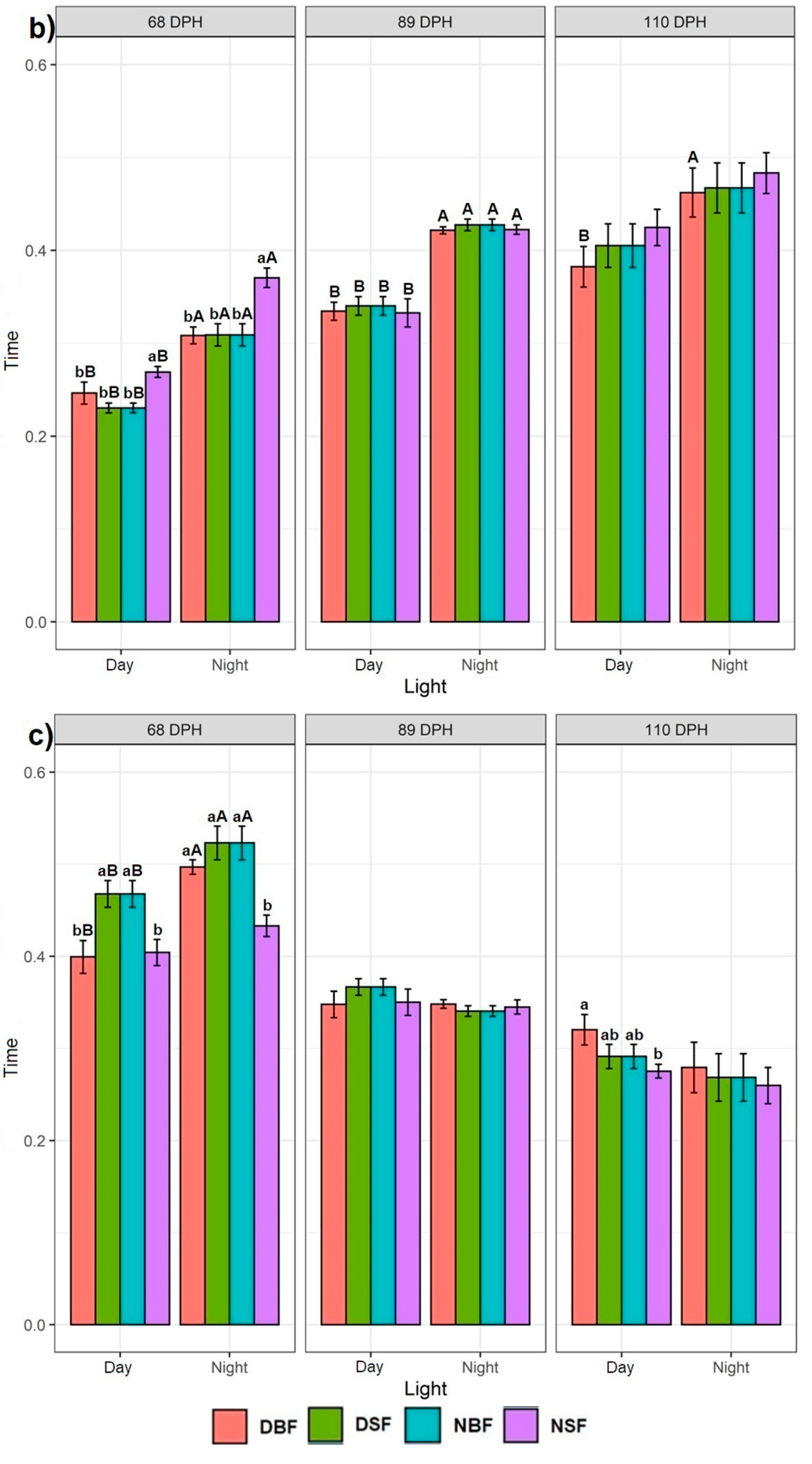
3.1.1. Effect of Daylight
3.1.2. Effect of Feeding Application
3.2. Effect of Timepoints Recorded at 68, 89, and 110 dph on Sturgeons Swimming Behaviour
3.2.1. Swimming Behaviour at One Particular Time Point
3.2.2. Effect of Consecutive Time Points
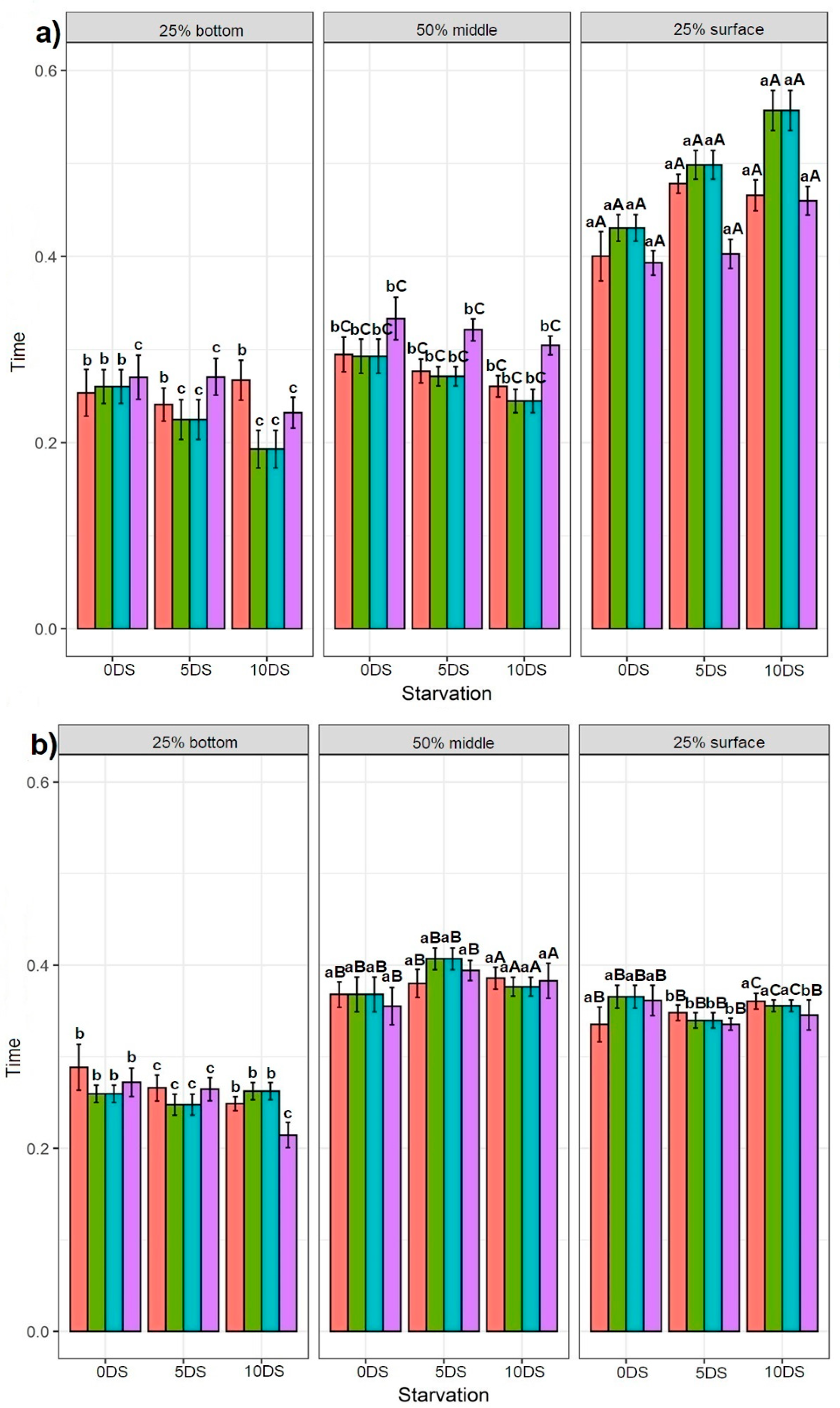
3.3. Effect of Feeding Application and 5 and 10 Days of Starvation on Sturgeons Swimming Behaviour
3.3.1. Effect of Feeding Application on the Starved Fish
3.3.2. Effect of 5 and 10 Days of Starvation on Sturgeon Behaviour
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Merino, G.; Barange, M.; Blanchard, J.L.; Harle, J.; Holmes, R.; Allen, I.; Allison, E.H.; Badjeck, M.C.; Dulvy, N.K.; Holt, J. Can marine fisheries and aquaculture meet fish demand from a growing human population in a changing climate? Glob. Environ. Chang. 2012, 22, 795–806. [Google Scholar] [CrossRef]
- Duarte, R.; Pinilla, V.; Serrano, A. Looking backward to look forward: Water use and economic growth from a long-term perspective. Appl. Econ. 2014, 46, 212–224. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations. The Future of Food and Agriculture—Trends and Challenges 2017; The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2017; ISBN 978-92-5-109551-5. [Google Scholar]
- Shiklomanov, I.A. Appraisal and assessment of world water resources. Water Int. 2000, 25, 11–32. [Google Scholar] [CrossRef]
- Dudgeon, D. Large-scale hydrological changes in tropical Asia: Prospects for riverine biodiversity: The construction of large dams will have an impact on the biodiversity of tropical Asian rivers and their associated wetlands. Bioscience 2000, 50, 793–806. [Google Scholar] [CrossRef]
- Nilsson, C.; Berggren, K. Alterations of riparian ecosystems caused by river regulation: Dam operations have caused global-scale ecological changes in riparian ecosystems. How to protect river environments and human needs of rivers remains one of the most important questions of our time. Bioscience 2000, 50, 783–792. [Google Scholar]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef]
- Copp, G.H. Effect of regulation on 0+ fish recruitment in the Great Ouse, a lowland river. Regul. Rivers Res. Manag. 1990, 5, 251–263. [Google Scholar] [CrossRef]
- Jurajda, P. Effect of channelization and regulation on fish recruitment in a flood plain river. Regul. Rivers Res. Manag. 1995, 10, 207–215. [Google Scholar] [CrossRef]
- Haro, A.; Richkus, W.; Whalen, K.; Hoar, A.; Busch, W.-D.; Lary, S.; Brush, T.; Dixon, D. Population decline of the American eel: Implications for research and management. Fisheries 2000, 25, 7–16. [Google Scholar] [CrossRef]
- Aarts, B.G.; Van Den Brink, F.W.; Nienhuis, P.H. Habitat loss as the main cause of the slow recovery of fish faunas of regulated large rivers in Europe: The transversal floodplain gradient. River Res. Appl. 2004, 20, 3–23. [Google Scholar] [CrossRef]
- Gao, X.; Brosse, S.; Chen, Y.; Lek, S.; Chang, J. Effects of damming on population sustainability of Chinese sturgeon, Acipenser sinensis: Evaluation of optimal conservation measures. Environ. Biol. Fishes 2009, 86, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wei, Q.; Du, H.; Li, L. Present status and risk for extinction of the Dabry’s sturgeon (Acipenser dabryanus) in the Yangtze River watershed: A concern for intensified rehabilitation needs. J. Appl. Ichthyol. 2011, 27, 181–185. [Google Scholar] [CrossRef]
- Haxton, T.J.; Cano, T.M. A global perspective of fragmentation on a declining taxon the sturgeon (Acipenseriformes). Endanger. Species Res. 2016, 31, 203–210. [Google Scholar] [CrossRef]
- Lenhardt, M.0; Jaric, I.; Kalauzi, A.; Cvijanovic, G. Assessment of extinction risk and reasons for decline in sturgeon. Biodivers. Conserv. 2006, 15, 1967–1976. [Google Scholar] [CrossRef]
- Bronzi, P.; Rosenthal, H. Present and future sturgeon and caviar production and marketing: A global market overview. J. Appl. Ichthyol. 2014, 30, 1536–1546. [Google Scholar] [CrossRef]
- Bronzi, P.; Rosenthal, H.; Gessner, J. Global sturgeon aquaculture production: An overview. J. Appl. Ichthyol. 2011, 27, 169–175. [Google Scholar] [CrossRef]
- Arndt, G.; Gessner, J.; Raymakers, C. Trends in farming, trade and occurrence of native and exotic sturgeons in natural habitats in Central and Western Europe. J. Appl. Ichthyol. 2002, 18, 444–448. [Google Scholar] [CrossRef]
- Knapp, A.; Kitschke, C.; Von Meibom, S. International Sturgeon Enforcement Workshop to Combat Illegal Trade in Caviar. In Proceeding of International Sturgeon Enforcement Workshop to Combat Illegal Trade in Caviar, Brussels, Belgium, 27–29 June 2006; Knapp, A., Kitschke, C., Von Meibom, S., Eds.; Traffic the Wildlife Trade Monitoring Network: Cambridge, UK, 2006; 155p. [Google Scholar]
- Navodaru, I.; Staraş, M. Conservation of fish stocks in the Danube Delta, Romania: Present status, constraints, and recommendation. Ital. J. Zool. 1998, 65, 369–371. [Google Scholar] [CrossRef]
- Williot, P.; Sabeau, L.; Gessner, J.; Arlati, G.; Bronzi, P.; Gulyas, T.; Berni, P. Sturgeon farming in Western Europe: Recent developments and perspectives. Aquat. Living Resour. 2001, 14, 367–374. [Google Scholar] [CrossRef]
- Wei, Q.; Zou, Y.; Li, P.; Li, L. Sturgeon aquaculture in China: Progress, strategies and prospects assessed on the basis of nation-wide surveys (2007–2009). J. Appl. Ichthyol. 2011, 27, 162–168. [Google Scholar] [CrossRef]
- Shen, L.; Shi, Y.; Zou, Y.; Zhou, X.; Wei, Q. Sturgeon Aquaculture in China: Status, challenge and proposals based on nation-wide surveys of 2010–2012. J. Appl. Ichthyol. 2014, 30, 1547–1551. [Google Scholar] [CrossRef]
- Brown, C.; Day, R.L. The future of stock enhancements: Lessons for hatchery practice from conservation biology. Fish Fish. 2002, 3, 79–94. [Google Scholar] [CrossRef] [Green Version]
- Ireland, S.C.; Beamesderfer, R.; Paragamian, V.; Wakkinen, V.; Siple, J. Success of hatchery-reared juvenile white sturgeon (Acipenser transmontanus) following release in the Kootenai River, Idaho, USA. J. Appl. Ichthyol. 2002, 18, 642–650. [Google Scholar] [CrossRef] [Green Version]
- Crossman, J.; Forsythe, P.; Scribner, K.; Baker, E. Hatchery rearing environment and age affect survival and movements of stocked juvenile lake sturgeon. Fish. Manag. Ecol. 2011, 18, 132–144. [Google Scholar] [CrossRef]
- Secor, D.; Houde, E. Use of larval stocking in restoration of Chesapeake Bay striped bass. ICES J. Mar. Sci. 1998, 55, 228–239. [Google Scholar] [CrossRef] [Green Version]
- Svåsand, T.; Kristiansen, T.S.; Pedersen, T.; Salvanes, A.V.; Engelsen, R.; Naevdal, G.; Nødtvedt, M. The enhancement of cod stocks. Fish Fish. 2000, 1, 173–205. [Google Scholar] [CrossRef]
- Myers, R.A.; Levin, S.A.; Lande, R.; James, F.C.; Murdoch, W.W.; Paine, R.T. Hatcheries and endangered salmon. Science 2004, 303, 1980. [Google Scholar] [CrossRef]
- Baer, J.; Blasel, K.; Diekmann, M. Benefits of repeated stocking with adult, hatchery-reared brown trout, Salmo trutta, to recreational fisheries? Fish. Manag. Ecol. 2007, 14, 51–59. [Google Scholar] [CrossRef]
- Bell, J.D.; Bartley, D.M.; Lorenzen, K.; Loneragan, N.R. Restocking and stock enhancement of coastal fisheries: Potential, problems and progress. Fish. Res. 2006, 80, 1–8. [Google Scholar] [CrossRef]
- Saloniemi, I.; Jokikokko, E.; Kallio-Nyberg, I.; Jutila, E.; Pasanen, P. Survival of reared and wild Atlantic salmon smolts: Size matters more in bad years. ICES J. Mar. Sci. 2004, 61, 782–787. [Google Scholar] [CrossRef]
- Thorstad, E.B.; Uglem, I.; Arechavala-Lopez, P.; Økland, F.; Finstad, B. Low survival of hatchery-released Atlantic salmon smolts during initial river and fjord migration. Boreal Environ. Res. 2011, 16, 115–120. [Google Scholar]
- Secor, D.; Arefjev, V.; Nikolaev, A.; Sharov, A. Restoration of sturgeons: Lessons from the Caspian Sea sturgeon ranching programme. Fish Fish. 2000, 1, 215–230. [Google Scholar] [CrossRef]
- Chebanov, M.; Rosenthal, H.; Gessner, J.; van Anrooy, R.; Doukakis, P.; Pourkazemi, M.; Williot, P. Sturgeon Hatchery Practices and Management for Release Guidelines; FAO Fisheries and Aquaculture Technical Paper No 570; FAO: Ankara, Turkey, 2011; pp. 2–110. [Google Scholar]
- McNeil, W.J. Expansion of cultured Pacific salmon into marine ecosystems. Aquaculture 1991, 98, 173–183. [Google Scholar] [CrossRef]
- Salvanes, A. 2001. Ocean ranching. In Marine Policy and Economics, 2nd ed.; Steele, J.H., Thorpe, S.A., Turekian, K.K., Eds.; Elsevier Ltd.: London, UK, 2010; pp. 226–235. [Google Scholar]
- Blaxter, J. The enhancement of marine fish stocks. Adv. Mar. Biol. 2000, 38, 2–54. [Google Scholar]
- Tanaka, Y.; Yamaguchi, H.; Gwak, W.-S.; Tominaga, O.; Tsusaki, T.; Tanaka, M. Influence of mass release of hatchery-reared Japanese flounder on the feeding and growth of wild juveniles in a nursery ground in the Japan Sea. J. Exp. Mar. Biol. Ecol. 2005, 314, 137–147. [Google Scholar] [CrossRef]
- Furuta, S. Releasing techniques and fry quality. In Healthy Fry for Release, and Their Production Techniques; Kitajima, C., Ed.; Koseisha-Koseikaku: Tokyo, Japan, 1993; pp. 94–101. (In Japanese) [Google Scholar]
- Miyazaki, T.; Masuda, R.; Furuta, S.; Tsukamoto, K. Feeding behaviour of hatchery-reared juveniles of the Japanese flounder following a period of starvation. Aquaculture 2000, 190, 129–138. [Google Scholar] [CrossRef]
- Wishingrad, V.; Ferrari, M.C.; Chivers, D.P. Behavioural and morphological defences in a fish with a complex antipredator phenotype. Anim. Behav. 2014, 95, 137–143. [Google Scholar] [CrossRef]
- Nguyen, R.M.; Crocker, C.E. The effects of substrate composition on foraging behavior and growth rate of larval green sturgeon, Acipenser medirostris. Environ. Biol. Fishes 2006, 76, 129–138. [Google Scholar] [CrossRef]
- Ross, R.; Bennett, R. Comparative behaviour and dietary effects in early life phases of American sturgeons. Fish. Manag. Ecol. 1997, 4, 17–30. [Google Scholar] [CrossRef]
- Chai, Y.; Tan, F.X.; Li, L.X.; Wei, Q.W. Effects of delayed initial feeding on growth and survival of Chinese sturgeon (Acipenser sinensis Gray, 1835) larvae. J. Appl. Ichthyol. 2014, 30, 1590–1595. [Google Scholar] [CrossRef]
- Hardy, R.S.; Zadmajid, V.; Butts, I.A.; Litvak, M.K. Growth, survivorship, and predator avoidance capability of larval shortnose sturgeon (Acipenser brevirostrum) in response to delayed feeding. PLoS ONE 2021, 16, e0247768. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H.; Chang, W.; Wickham, M.H. Package ‘ggplot2’. In Create Elegant Data Visualisations Using the Grammar of Graphics, 2nd ed.; Springer: New York, NY, USA, 2016; pp. 1–189. [Google Scholar]
- Jokikokko, E.; Kallio-Nyberg, I.; Saloniemi, I.; Jutila, E. The survival of semi-wild, wild and hatchery-reared Atlantic salmon smolts of the Simojoki River in the Baltic Sea. J. Fish. Biol. 2006, 68, 430–442. [Google Scholar] [CrossRef]
- Czerniawski, R.; Pilecka-Rapacz, M.; Domagala, J. Stocking experiment with Atlantic salmon and sea trout parr reared on either live prey or a pellet diet. J. Appl. Ichthyol. 2011, 27, 984–989. [Google Scholar] [CrossRef]
- Tomiyama, T.; Watanabe, M.; Kawata, G.; Ebe, K. Post-release feeding and growth of hatchery-reared Japanese flounder Paralichthys olivaceus: Relevance to stocking effectiveness. J. Fish. Biol. 2011, 78, 1423–1436. [Google Scholar] [CrossRef] [PubMed]
- Krepski, T.; Czerniawski, R. Can we teach a fish how to eat? The impact of bottom and surface feeding on survival and growth of hatchery-reared sea trout parr (Salmo trutta trutta L.) in the wild. PLoS ONE 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kellison, G.; Eggleston, D.; Burke, J. Comparative behaviour and survival of hatchery-reared versus wild summer flounder (Paralichthys dentatus). Can. J. Fish. Aquat. 2000, 57, 1870–1877. [Google Scholar] [CrossRef]
- Moberg, O.; Braithwaite, V.A.; Jensen, K.H.; Salvanes, A.G.V. Effects of habitat enrichment and food availability on the foraging behaviour of juvenile Atlantic Cod (Gadus morhua L.). Environ. Biol. Fishes. 2011, 91, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Gisbert, E. Early development and allometric growth patterns in Siberian sturgeon and their ecological significance. J. Fish. Biol. 1999, 54, 852–862. [Google Scholar] [CrossRef]
- Gisbert, E.; Williot, P.; Castelló-Orvay, F. Behavioural modifications in the early life stages of Siberian sturgeon (Acipenser baerii, Brandt). J. Appl. Ichthyol. 1999, 15, 237–242. [Google Scholar] [CrossRef]
- Takahashi, K.; Masuda, R.; Yamashita, Y. Bottom feeding and net chasing improve foraging behavior in hatchery-reared Japanese flounder Paralichthys olivaceus juveniles for stocking. Fish. Sci. 2013, 79, 55–60. [Google Scholar] [CrossRef]
- Steel, A.; Hansen, M.; Cocherell, D.; Fangue, N. Behavioral responses of juvenile white sturgeon (Acipenser transmontanus) to manipulations of nutritional state and predation risk. Environ. Biol. Fishes. 2019, 102, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Kasumyan, A.; Kazhlayev, A. Formation of Searching Behavioral Reaction and Olfactory Sensitivity to Food Chemical Signals during Onlogeny of Sturgeons (Acipenseridae). J. Ichthyol. 1993, 33, 51–65. [Google Scholar]
- Tanaka, M.; Goto, T.; Tomiyama, M.; Sudo, H. Immigration, settlement and mortality of flounder (Paralichthys olivaceus) larvae and juveniles in a nursery ground, Shijiki Bay, Japan. Neth. J. Sea Res. 1989, 24, 57–67. [Google Scholar] [CrossRef]
- Furuta, S. Comparison of feeding behavior of wild and hatchery-reared Japanese flounder, Paralichthys olivaceus, juveniles by laboratory experiments. Nippon Suisan Gakkaishi 1998, 64, 393–397. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, Y.; Yamamoto, K.; Nagahora, S.; Igarashi, K.; Ishikawa, Y.; Sakuma, O.; Yamada, H.; Nakamoto, Y. Predation by fishes on hatchery-raised Japanese flounder, Paralichthys olivaceus, fry in the coastal waters of Iwate prefecture, northeastern Japan. Aquac. Sci. 1993, 41, 497–505. [Google Scholar]
- Richmond, A.M.; Kynard, B. Ontogenetic behavior of shortnose sturgeon, Acipenser brevirostrum. Copeia 1995, 1995, 172–182. [Google Scholar] [CrossRef]
- Wishingrad, V.; Musgrove, A.B.; Chivers, D.P.; Ferrari, M.C. Risk in a changing world: Environmental cues drive anti-predator behaviour in lake sturgeon (Acipenser fulvescens) in the absence of predators. Behaviour 2015, 152, 635–652. [Google Scholar] [CrossRef] [Green Version]
- Sundström, L.F.; Petersson, E.; Höjesjö, J.; Johnsson, J.I.; Järvi, T. Hatchery selection promotes boldness in newly hatched brown trout (Salmo trutta): Implications for dominance. Behav. Ecol. 2004, 15, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Fernö, A.; Huse, G.; Jakobsen, P.J.; Kristiansen, T.S.; Nilsson, J. Fish behaviour, learning, aquaculture and fisheries. In Fish Cognition and Behavior, 2nd ed.; Brown, C., Laland, K., Krause, J., Eds.; Wiley-Blackwell: Oxford, UK, 2011; pp. 359–404. [Google Scholar]
- Jackson, C.D.; Brown, G.E. Differences in antipredator behaviour between wild and hatchery-reared juvenile Atlantic salmon (Salmo salar) under seminatural conditions. Can. J. Fish. Aquat. 2011, 68, 2157–2166. [Google Scholar] [CrossRef]
- Alvarez, D.; Nicieza, A. Predator avoidance behaviour in wild and hatchery-reared brown trout: The role of experience and domestication. J. Fish. Biol. 2003, 63, 1565–1577. [Google Scholar] [CrossRef]
- Kawabata, Y.; Asami, K.; Kobayashi, M.; Sato, T.; Okuzawa, K.; Yamada, H.; Yoseda, K.; Arai, N. Effect of shelter acclimation on the post-release movement and putative predation mortality of hatchery-reared black-spot tuskfish Choerodon schoenleinii, determined by acoustic telemetry. Fish. Sci. 2011, 77, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Mirza, R.S.; Chivers, D.P. Predator-recognition training enhances survival of brook trout: Evidence from laboratory and field-enclosure studies. Can. J. Zool. 2000, 78, 2198–2208. [Google Scholar] [CrossRef]
- Cámara Ruiz, M.; Espírito Santo, C.; Mai, A.; Gessner, J.; Wuertz, S. Can juvenile Baltic sturgeon (Acipenser oxyrinchus) smell the enemy? J. Appl. Ichthyol. 2019, 35, 835–842. [Google Scholar] [CrossRef]
- Brown, G.E.; Ferrari, M.C.; Chivers, D.P. Learning about danger: Chemical alarm cues and threat-sensitive assessment of predation risk by fishes. In Fish Cognition and Behavior, 2nd ed.; Brown, C., Laland, K., Krause, J., Eds.; Wiley-Blackwell: Oxford, UK, 2011; pp. 59–80. [Google Scholar]
- Lappalainen, J.; Dörner, H.; Wysujack, K. Reproduction biology of pikeperch (Sander lucioperca (L.))—A review. Ecol. Freshw. Fish 2003, 12, 95–106. [Google Scholar] [CrossRef]
- Komsari, M.S.; Bani, A.; Khara, H. Growth and population structure of the European perch, Perca fluviatilis Linnaeus, 1758 (Osteichthyes: Percidae) in the Anzali Wetland south-west Caspian Sea. Indian J. Fish. 2015, 62, 6–11. [Google Scholar]
- Khara, H.; Sattari, M. Occurrence and intensity of parasites in Wels catfish, Silurus glanis L. 1758 from Amirkelayeh wetland, southwest of the Caspian Sea. J. Parasit. Dis. 2016, 40, 848–852. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimzadeh Kouchesfahani, N.; Forouhar Vajargah, M. A short review on the biological characteristics of the species Esox lucius, Linnaeus, 1758 in Caspian Sea Basin (Iran). Transylv. Rev. Syst. Ecol. Res. 2021, 1, 1–23. [Google Scholar]
- Templeton, C.N.; Shriner, W.M. Multiple selection pressures influence Trinidadian guppy (Poecilia reticulata) antipredator behavior. Behav. Ecol. 2004, 15, 673–678. [Google Scholar] [CrossRef]
- Darwish, T.L.; Mirza, R.S.; Leduc, A.O.; Brown, G.E. Acquired recognition of novel predator odour cocktails by juvenile glowlight tetras. Anim. Behav. 2005, 70, 83–89. [Google Scholar] [CrossRef]
- Hintz, W.; Grimes, G.; Garvey, J. Shovelnose sturgeon exhibit predator avoidance behaviour in the presence of a hungry predator. J. Appl. Ichthyol. 2013, 29, 2–5. [Google Scholar] [CrossRef]
- Hawkins, L.A.; Magurran, A.E.; Armstrong, J.D. Ontogenetic learning of predator recognition in hatchery-reared Atlantic salmon, Salmo salar. Anim. Behav. 2008, 75, 1663–1671. [Google Scholar] [CrossRef]
- Sloychuk, J.R.; Chivers, D.P.; Ferrari, M.C. Juvenile lake sturgeon go to school: Life-skills training for hatchery fish. Trans. Am. Fish. Soc. 2016, 145, 287–294. [Google Scholar] [CrossRef]
- Wishingrad, V.; Sloychuk, J.; Ferrari, M.; Chivers, D. Alarm cues in Lake Sturgeon Acipenser fulvescens Rafinesque, 1817: Potential implications for life-skills training. J. Appl. Ichthyol. 2014, 30, 1441–1444. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebauer, T.; Gebauer, R.; Císař, P.; Tran, H.Q.; Tomášek, O.; Podhorec, P.; Prokešová, M.; Rebl, A.; Stejskal, V. The Effect of Different Feeding Applications on the Swimming Behaviour of Siberian Sturgeon: A Method for Improving Restocking Programmes. Biology 2021, 10, 1162. https://doi.org/10.3390/biology10111162
Gebauer T, Gebauer R, Císař P, Tran HQ, Tomášek O, Podhorec P, Prokešová M, Rebl A, Stejskal V. The Effect of Different Feeding Applications on the Swimming Behaviour of Siberian Sturgeon: A Method for Improving Restocking Programmes. Biology. 2021; 10(11):1162. https://doi.org/10.3390/biology10111162
Chicago/Turabian StyleGebauer, Tatyana, Radek Gebauer, Petr Císař, Hung Quang Tran, Ondřej Tomášek, Peter Podhorec, Markéta Prokešová, Alexander Rebl, and Vlastimil Stejskal. 2021. "The Effect of Different Feeding Applications on the Swimming Behaviour of Siberian Sturgeon: A Method for Improving Restocking Programmes" Biology 10, no. 11: 1162. https://doi.org/10.3390/biology10111162
APA StyleGebauer, T., Gebauer, R., Císař, P., Tran, H. Q., Tomášek, O., Podhorec, P., Prokešová, M., Rebl, A., & Stejskal, V. (2021). The Effect of Different Feeding Applications on the Swimming Behaviour of Siberian Sturgeon: A Method for Improving Restocking Programmes. Biology, 10(11), 1162. https://doi.org/10.3390/biology10111162