
Rhizoplane and Rhizosphere Fungal Communities of Geographically Isolated Korean Bellflower (Campanula takesimana Nakai)

 ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling of the Host Plant
2.2. DNA Extraction and Polymerase Chain Reaction Amplification
2.3. Sequence Processing and Data Analyses
3. Results and Discussion
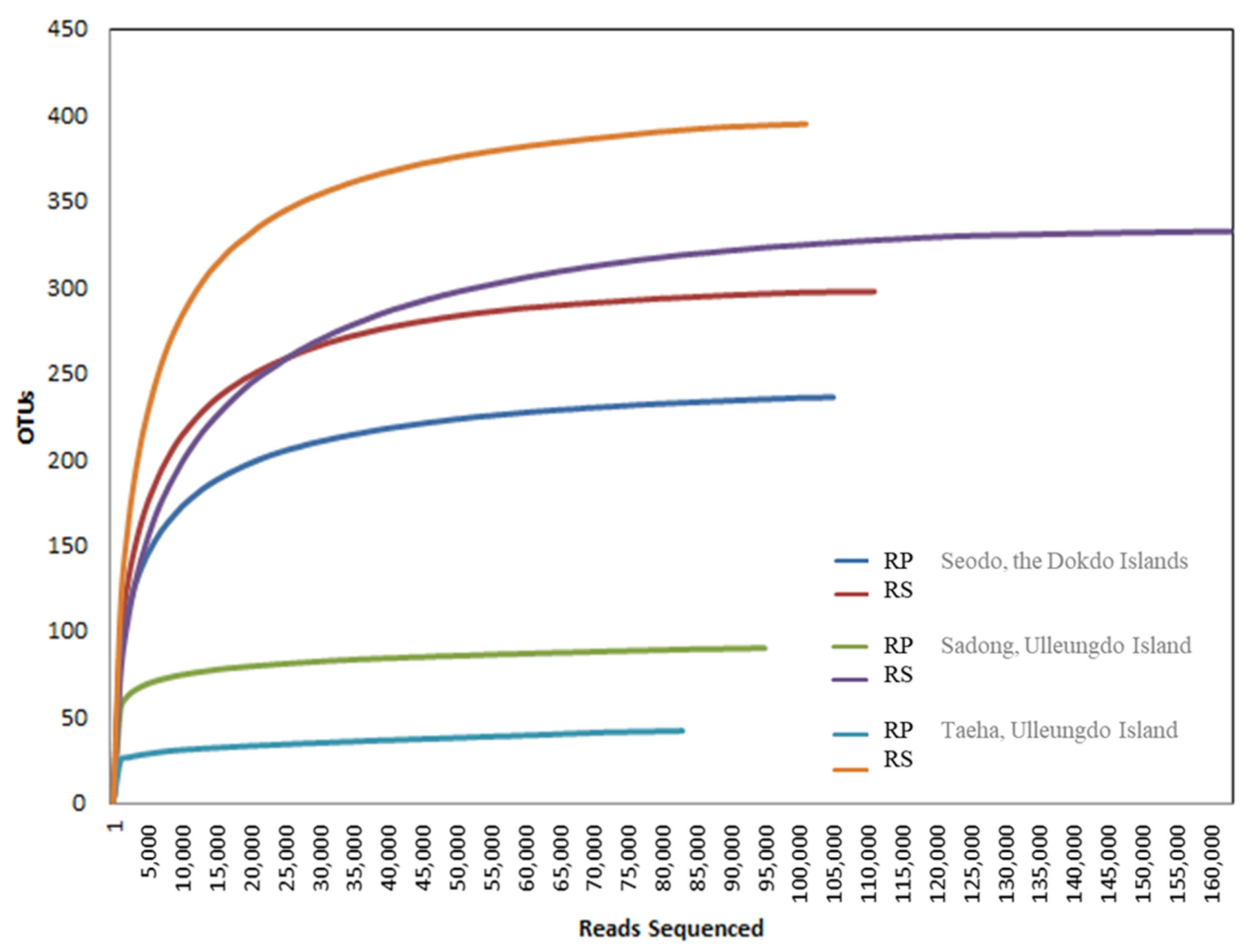
3.1. Illumina MiSeq Sequencing Results
3.2. Variation of the Fungal Phyla Associated with Geographically Isolated Hosts
3.3. Variation of Fungal Diverstity
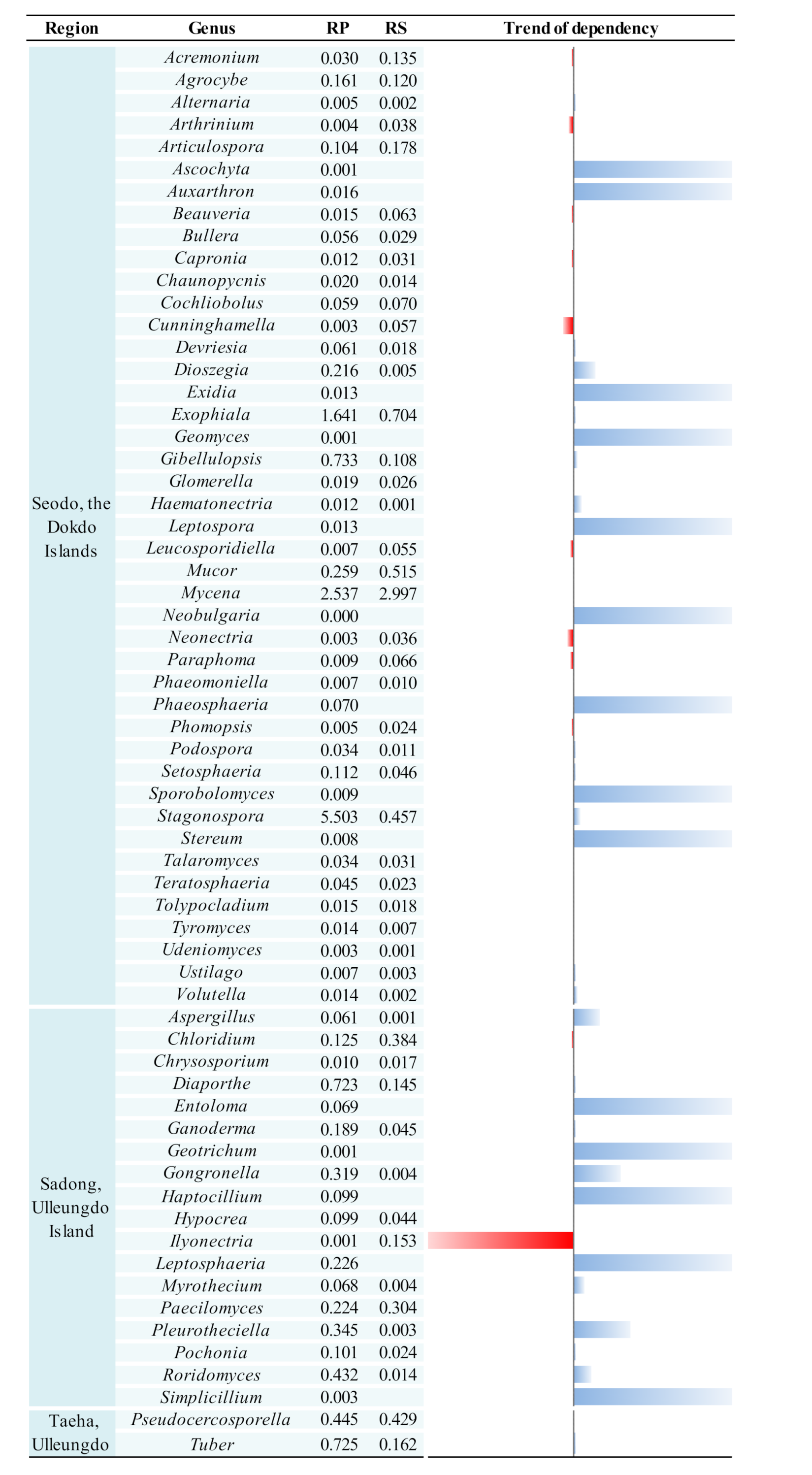
3.4. Variation of Fungal Genera
3.5. Host Dependency and Selective Pressure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiersema, J.H. GRIN Taxonomy. US National Plant Germplasm System. Checklist Dataset. 2019. Available online: https://doi.org/10.15468/ao14pp (accessed on 4 April 2020).
- Brandt, K.; Kondo, T.; Aoki, H.; Goto, T. Structure and biosynthesis of anthocyanins in flowers of Campanula. Phytochemistry 1993, 33, 209–212. [Google Scholar] [CrossRef]
- Stuchlíková, L.; Jirásko, R.; Skálová, L.; Pavlík, F.; Szotáková, B.; Holčapek, M.; Vaněk, T.; Podlipná, R. Metabolic pathways of benzimidazole anthelmintics in harebell (Campanula rotundifolia). Chemosphere 2016, 157, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Folquitto, D.G.; Swiech, J.N.D.; Pereira, C.B.; Bobek, V.B.; Halila Possagno, G.C.; Farago, P.V.; Miguel, M.D.; Duarte, J.L.; Miguel, O.G. Biological activity, phytochemistry and traditional uses of genus Lobelia (Campanulaceae): A systematic review. Fitoterapia 2019, 134, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.Y.; Bo, A.; Yang, M.; Xu, J.F.; Jiang, L.L.; Zhou, B.C.; Li, M.H. The pharmacological effects and health benefits of Platycodon grandiflorus-A medicine food homology species. Foods 2020, 9, 142. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.; Wang, P.; Wang, X.; Zhou, X.; Hu, X.; Chen, F. Dietary Platycodon grandiflorus attenuates hepatic insulin resistance and oxidative stress in high-fat-diet induced non-alcoholic fatty liver disease. Nutrients 2020, 12, 480. [Google Scholar] [CrossRef] [PubMed]
- Cultural Heritage Administration (CHA) of Korea. Natural Heritage of Korea, Dokdo. Available online: http://116.67.83.213/NEW_PDF/EM036763_01.pdf (accessed on 17 October 2020).
- Park, J.M.; Hong, J.W.; Son, J.S.; Hwang, Y.J.; Cho, H.M.; You, Y.H.; Ghim, S.Y. A strategy for securing unique microbial resources–focusing on Dokdo islands-derived microbial resources. Isr. J. Ecol. Evol. 2018, 64, 1–15. [Google Scholar] [CrossRef]
- Park, K.R.; Jung, H.J. Isozyme and morphological variation in Campanula punctata and C. takesimana (Campanulaceae). Korean J. Plant Taxon. 2000, 30, 1–16. [Google Scholar] [CrossRef]
- Shulkina, T.V.; Gaskin, J.F.; Eddie, W.M.M. Morphological studies toward an improved classification of Campanulaceae s. str. Ann. MO. Bot. Gard. 2003, 90, 576–591. [Google Scholar] [CrossRef]
- Ahn, Y.H.; Jo, D.K.; Kim, K.S.; Lee, J.H.; Shin, C.H.; Lee, S.J.; Kang, K.H. Ecological characteristics of native Campanula takesimana habitats in Ulleung-do Island. J. Plant Environ. 2005, 1, 19–24. [Google Scholar]
- Thorpe, R.S.; Surget-Groba, Y.; Johansson, H. Genetic tests for ecological and allopatric speciation in anoles on an island archipelago. PLoS Genet. 2010, 6, e1000929. [Google Scholar] [CrossRef] [PubMed]
- Aseel, D.G.; Rashad, Y.M.; Hammad, S.M. Arbuscular mycorrhizal fungi trigger transcriptional expression of flavonoid and chlorogenic acid biosynthetic pathways genes in tomato against Tomato Mosaic Virus. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Haruma, T.; Yamaji, K.; Ogawa, K.; Masuya, H.; Sekine, Y.; Kozai, N. Root-endophytic Chaetomium cupreum chemically enhances aluminium tolerance in Miscanthus sinensis via increasing the aluminium detoxicants, chlorogenic acid and oosporein. PLoS ONE 2019, 14, e0212644. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, C.B.; Ma, X.G.; Liang, Q.L.; He, X.J. Phylogeographic analysis of a temperate-deciduous forest restricted plant (Bupleurum longiradiatum Turcz.) reveals two refuge areas in China with subsequent refugial isolation promoting speciation. Mol. Phylogenet. Evol. 2013, 68, 628–643. [Google Scholar] [CrossRef] [PubMed]
- Baez-Rogelio, A.; Morales-García, Y.E.; Quintero-Hernández, V.; Muñoz-Rojas, J. Next generation of microbial inoculants for agriculture and bioremediation. Microb. Biotechnol. 2017, 10, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Perlińska-Lenart, U.; Piłsyk, S.; Gryz, E.; Turło, J.; Hilszczańska, D.; Kruszewska, J.S. Identification of bacteria and fungi inhabiting fruiting bodies of Burgundy truffle (Tuber aestivum Vittad.). Arch. Microbiol. 2020, 202, 2727–2738. [Google Scholar] [CrossRef] [PubMed]
- National Geographic Information Institute (NGII) of Korea. Geography of Dokdo. 2015. Available online: https://map.ngii.go.kr/ms/pblictn/dokdoGrphHist.do (accessed on 17 October 2020).
- You, Y.H.; Park, J.M.; Park, J.H.; Kim, J.G. Specific rhizobacterial resources: Characterization and comparative analysis from contrasting coastal environments of Korea. J. Basic Microbiol. 2016, 56, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.S.; Watts, P. The Plant Geography of Korea with an Emphasis on the Alpine Zones; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Vega, N.W.O. A review on beneficial effects of rhizosphere bacterial on soil nutrient availability and plant nutrient uptake. Rev. Environ. Contam. Toxicol. 2007, 223, 33–52. [Google Scholar]
- Flores-Vargas, R.D.; O’Hara, G.W. Isolation and characterization of rhizosphere bacterial with potential for biological control of weed in vineyards. J. Appl. Microbiol. 2006, 100, 946–954. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sánchez-García, M.; Ebersberger, I.; de Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Hoiland, K.; Kjoller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi-recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Huttley, G.A. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Heck, K.L.; van Belle, G.; Simberloff, D. Explicit calculation of the rarefaction diversity measurement and the determination of sufficient sample size. Ecology 1975, 56, 1459–1461. [Google Scholar] [CrossRef]
- Lambshead, P.J.D.; Platt, H.M.; Shaw, K.M. The detection of differences among assemblages of marine benthic species based on an assessment of dominance and diversity. J. Nat. Hist. 1983, 17, 859–874. [Google Scholar] [CrossRef]
- Chao, A.; Shen, T.J. Program SPADE (Species Prediction and Diversity Estimation) Program and User’s Guide; National Tsing Hua University: Hsinchu, Taiwan, 2010; Available online: http://chao.stat.nthu.edu.tw/wordpress/ (accessed on 17 October 2020).
- Chapman, V.J. Salt Marshes and Salt Deserts of the World. In Ecology of Halophytes; Reimold, R.J., Queen, W.H., Eds.; Academic Press: New York, NY, USA, 1974; pp. 3–19. [Google Scholar]
- Von Caemmerer, S.; Edwards, G.E.; Koteyeva, N.; Cousins, A.B. Single-cell C4 photosynthesis in aquatic and terrestrial plants: A gas exchange perspective. Aquat. Bot. 2014, 118, 71–80. [Google Scholar] [CrossRef]
- Sage, R.F. A portrait of the C4 photosynthetic family on the 50th anniversary of its discovery: Species number, evolutionary lineages, and Hall of Fame. J. Exp. Bot. 2017, 68, e11–e28. [Google Scholar] [CrossRef]
- Barton, L.L.; Northup, D.E. Microbial Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Deacon, J.W. Fungal Biology, 4th ed.; Wiley-Blackwell: Malden, MA, USA, 2005. [Google Scholar]
- Webster, J.; Weber, R.W.S. Introduction to Fungi; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Min, Y.J.; Park, M.S.; Fong, J.J.; Quan, Y.; Jung, S.; Lim, Y.W. Diversity and saline resistance of endophytic fungi associated with Pinus thunbergii in coastal shelterbelts of Korea. J. Microbiol. Biotechnol. 2014, 24, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F.; Kubota, M.; Koyama, H.; Hyakumachi, M. The plant growth promoting fungus Penicillium simplicissimum GP17-2 induces resistance in Arabidopsis thaliana by activation of multiple defense signals. Plant Cell Physiol. 2007, 48, 1724–1736. [Google Scholar] [CrossRef]
- Khan, S.A.; Hamayun, M.; Yoon, H.; Kim, H.Y.; Suh, S.J.; Hwang, S.K.; Kim, J.M.; Lee, I.J.; Choo, Y.S.; Yoon, U.H.; et al. Plant growth promotion and Penicillium citrinum. BMC Microbiol. 2008, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Del Pilar Martínez-Diz, M.; Andrés-Sodupe, M.; Berbegal, M.; Bujanda, R.; Díaz-Losada, E.; Gramaje, D. Droplet digital PCR technology for detection of Ilyonectria liriodendri from grapevine environmental samples. Plant Dis. 2020, 104, 1144–1150. [Google Scholar] [CrossRef]
- Farh, M.E.; Kim, Y.J.; Singh, P.; Yang, D.C. Cross interaction between Ilyonectria mors-panacis isolates infecting Korean ginseng and ginseng saponins in correlation with their pathogenicity. Phytopathology 2017, 107, 561–569. [Google Scholar] [CrossRef]
- Farh, M.E.; Kim, Y.J.; Abbai, R.; Singh, P.; Jung, K.H.; Kim, Y.J.; Yang, D.C. Pathogenesis strategies and regulation of ginsenosides by two species of Ilyonectria in Panax ginseng: Power of speciation. J. Ginseng Res. 2020, 44, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Chen, M.; Ma, Y.; Du, Z.; Yuan, N.; Li, Y.; Xiao, J.; Zhang, Y. Whole-genome and time-course dual RNA-Seq analyses reveal chronic pathogenicity-related gene dynamics in the ginseng rusty root rot pathogen Ilyonectria robusta. Sci. Rep. 2020, 10, 1586. [Google Scholar] [CrossRef]
- Liao, H.L.; Bonito, G.; Rojas, J.A.; Hameed, K.; Wu, S.; Schadt, C.W.; Labbé, J.; Tuskan, G.A.; Martin, F.; Grigoriev, I.V.; et al. Fungal endophytes of Populus trichocarpa alter host phenotype, gene expression, and rhizobiome composition. Mol. Plant Microbe Interact. 2019, 32, 853–864. [Google Scholar] [CrossRef]
- Deng, J.X.; Paul, N.C.; Li, M.J.; Cho, H.S.; Lee, H.B.; Yu, S.H. Stemphylium platycodontis sp. nov., isolated from Platycodon grandiflorus in Korea. Mycol. Prog. 2014, 13, 477–482. [Google Scholar] [CrossRef]
- Olsen, K.J.K.; Rossman, A.; Andersen, B. Metabolite production by species of Stemphylium. Fungal Biol. 2018, 122, 172–181. [Google Scholar] [CrossRef]
- Johnson, N.C.; Graham, J.H.; Smith, F.A. Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytol. 1997, 135, 575–585. [Google Scholar] [CrossRef]
- Bäumler, A.; Fang, F.C. Host specificity of bacterial pathogens. Cold Spring Harb. Perspect. Med. 2013, 3, a010041. [Google Scholar]
- Morris, C.E.; Moury, B. Revisiting the concept of host range of plant pathogens. Annu. Rev. Phytopathol. 2019, 57, 63–90. [Google Scholar] [CrossRef]
- Kutcher, H.R.; Johnston, A.M.; Bailey, K.L.; Malhi, S.S. Managing crop losses from plant diseases with foliar fungicides, rotation and tillage on a Black Chernozem in Saskatchewan, Canada. Field Crops Res. 2011, 124, 205–212. [Google Scholar] [CrossRef]
- Atiphasaworn, P.; Monggoot, S.; Gentekaki, E.; Brooks, S.; Pripdeevech, P. Antibacterial and antioxidant constituents of extracts of endophytic fungi isolated from Ocimum basilicum var. thyrsiflora leaves. Curr. Microbiol. 2017, 74, 1185–1193. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; David, J.C.; Stalpers, J.A. Ainsworth and Bisby’s Dictionary of the Fungi, 9th ed.; CABI Publishing: Hague, The Netherlands, 2001. [Google Scholar]
- Blehert, D.S.; Hicks, A.C.; Behr, M.; Meteyer, C.U.; Berlowski-Zier, B.M.; Buckles, E.L.; Coleman, J.T.H.; Darling, S.R.; Gargas, A.; Niver, R.; et al. Bat white-nose syndrome: An emerging fungal pathogen? Science 2009, 323, 227. [Google Scholar] [CrossRef] [PubMed]
- Kryvomaz, T.I.; Michaud, A.; Minter, D.W. IMI Descriptions of Fungi and Bacteria; CABI Publication: Haue, The Netherlands, 2019. [Google Scholar]
- Li, C.J.; Gao, J.H.; Nan, Z.B. Interactions of Neotyphodium gansuense, Achnatherum inebrians, and plant-pathogenic fungi. Mycol. Res. 2007, 111, 1220–1227. [Google Scholar] [CrossRef]
- Ozimek, E.; Jaroszuk-Ściseł, J.; Bohacz, J.; Korniłłowicz-Kowalska, T.; Tyśkiewicz, R.; Słomka, A.; Nowak, A.; Hanaka, A. Synthesis of indoleacetic acid, gibberellic acid and ACC-deaminase by Mortierella strains promote winter wheat seedlings growth under different conditions. Int. J. Mol. Sci. 2018, 19, 3218. [Google Scholar] [CrossRef]
- Davies, J.L.; Wobeser, G.A. Systemic infection with Mortierella wolfii following abortion in a cow. Can. Vet. J. 2010, 51, 1391. [Google Scholar]
- Zhang, H.; Wu, X.; Li, G.; Qin, P. Interactions between arbuscular mycorrhizal fungi and phosphate-solubilizing fungus (Mortierella sp.) and their effects on Kostelelzkya virginica growth and enzyme activities of rhizosphere and bulk soils at different salinities. Biol. Fertil. Soils 2011, 47, 543. [Google Scholar] [CrossRef]
- Toju, H.; Sato, H. Root-associated fungi shared between arbuscular mycorrhizal and ectomycorrhizal conifers in a temperate forest. Front. Microbiol. 2018, 9, 433. [Google Scholar] [CrossRef] [PubMed]
- Wani, Z.A.; Kumar, A.; Sultan, P.; Bindu, K.; Riyaz-Ul-Hassan, S.; Ashraf, N. Mortierella alpina CS10E4, an oleaginous fungal endophyte of Crocus sativus L. enhances apocarotenoid biosynthesis and stress tolerance in the host plant. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.S.; Qin, F.F.; Qin, P.; Pan, S.M. Evidence that arbuscular mycorrhizal and phosphate-solubilizing fungi alleviate NaCl stress in the halophyte Kosteletzkya virginica: Nutrient uptake and ion distribution within root tissues. Mycorrhiza 2014, 24, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Yuste, J.C.; Fernandez-Gonzalez, A.J.; Fernandez-Lopez, M.; Ogaya, R.; Penuelas, J.; Sardans, J.; Lloret, F. Strong functional stability of soil microbial communities under semiarid Mediterranean conditions and subjected to long-term shifts in baseline precipitation. Soil Biol. Biochem. 2014, 69, 223–233. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Coordinates (Latitude, Longitude) | Avg. Altitude (m) | Angle of Slope (°) | Geographical Feature | Avg. Height (cm) | Avg. Density (Individual/m2) | Surrounding Vegetation | Avg. Area Occupied by a Colony (m2) |
|---|---|---|---|---|---|---|---|---|
| Seodo, the Dokdo Islands | 37°14′31.2″ N, 131°51′53.0″ E | 87 | 45 | Small oceanic volcanic island | 5 | 20 | Herbaceous plants | 1 |
| Sadong, Ulleungdo Island | 37°27′23.6″ N, 130°52′30.0″ E | 10 | 20 | Coastline of large volcanic island | 25 | 40 | Herbaceous plants | 9 |
| Taeha, Ulleungdo Island | 37°29′40.8″ N, 130°49′39.4″ E | 425 | 5 | Inland area of large volcanic island | 15 | 80 | Herbaceous plants | 6 |
| Sampling Site | Seodo, the Dokdo Islands | Sadong, Ulleungdo Island | Taeha, Ulleungdo Island | |||
|---|---|---|---|---|---|---|
| Sampling Point | RP | RS | RP | RS | RP | RS |
| Total read counts | 240,956 | 247,847 | 194,918 | 285,563 | 160,662 | 238,724 |
| Total bases | 87,920,927 | 92,067,817 | 76,300,872 | 105,684,291 | 63,478,745 | 88,850,527 |
| Number of valid sequences | 103,130 | 105,988 | 93,402 | 142,825 | 78,860 | 95,022 |
| GC (%) | 49.7 | 49.6 | 51.8 | 52.9 | 47.8 | 49.0 |
| OTUs | 236 | 298 | 90 | 333 | 42 | 395 |
| Fungal phylum | 5 | 4 | 3 | 5 | 3 | 5 |
| Fungal genera | 58 | 71 | 34 | 85 | 15 | 101 |
| Genus | Seodo, the Dokdo Islands | Sadong, Ulleungdo Island | Taeha, Ulleungdo Island | |||
|---|---|---|---|---|---|---|
| RP (%) | RS (%) | RP (%) | RS (%) | RP (%) | RS (%) | |
| Fusarium | 1.225 | 1.154 | 0.541 | 0.096 | 0.001 | 0.049 |
| Mortierella | 7.780 | 13.064 | 3.541 | 1.732 | 21.260 | 23.204 |
| Penicillium | 1.063 | 1.020 | 0.246 | 0.267 | 0.001 | 0.363 |
| Trichoderma | 0.778 | 0.104 | 0.386 | 0.320 | 2.205 | 0.155 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.M.; Kim, B.; Cho, Y.-C.; Lee, B.-H.; Hong, J.W.; You, Y.-H. Rhizoplane and Rhizosphere Fungal Communities of Geographically Isolated Korean Bellflower (Campanula takesimana Nakai). Biology 2021, 10, 138. https://doi.org/10.3390/biology10020138
Park JM, Kim B, Cho Y-C, Lee B-H, Hong JW, You Y-H. Rhizoplane and Rhizosphere Fungal Communities of Geographically Isolated Korean Bellflower (Campanula takesimana Nakai). Biology. 2021; 10(2):138. https://doi.org/10.3390/biology10020138
Chicago/Turabian StylePark, Jong Myong, Bomi Kim, Young-Chang Cho, Byoung-Hee Lee, Ji Won Hong, and Young-Hyun You. 2021. "Rhizoplane and Rhizosphere Fungal Communities of Geographically Isolated Korean Bellflower (Campanula takesimana Nakai)" Biology 10, no. 2: 138. https://doi.org/10.3390/biology10020138
APA StylePark, J. M., Kim, B., Cho, Y.-C., Lee, B.-H., Hong, J. W., & You, Y.-H. (2021). Rhizoplane and Rhizosphere Fungal Communities of Geographically Isolated Korean Bellflower (Campanula takesimana Nakai). Biology, 10(2), 138. https://doi.org/10.3390/biology10020138