Phylogenetic Relatedness within the Internally Brooding Sea Anemones from the Arctic-Boreal Region

 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Molecular Data Analysis
2.2. Ecological and Morphological Data Analysis
3. Results
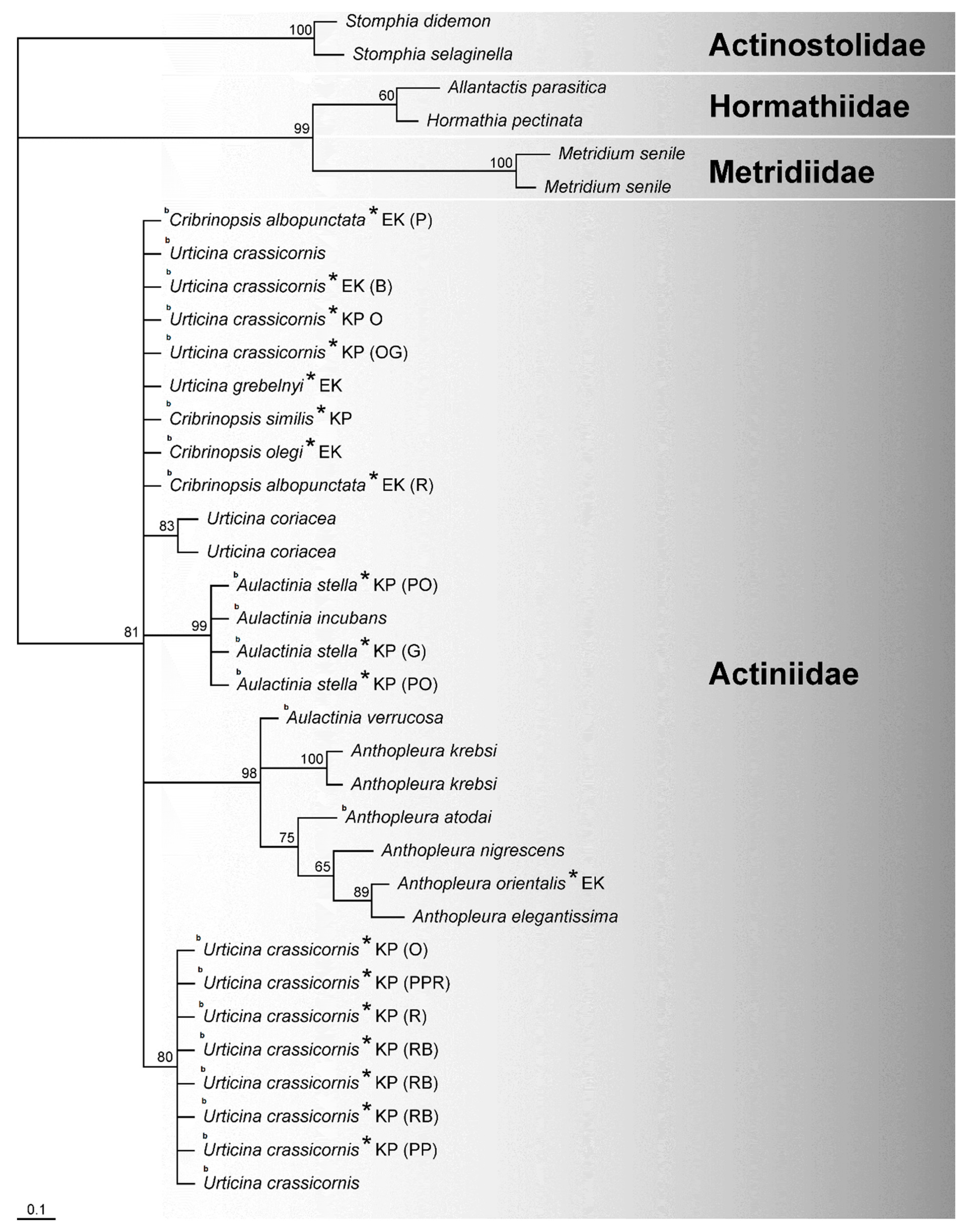
3.1. Alignments and Trees Based on Molecular Characteristics
3.2. Trees Based on Morphological and Ecological Characters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 16S rDNA | the sequence of mitochondrial small subunit of a ribosome |
| 28S rDNA | the sequence of nuclear subunit of a ribosome |
| B | color of sea anemones, beige |
| CP | Central Poland |
| EK | East Kamchatka |
| FJL | Franz Josef Land |
| G | color of sea anemones, grey |
| KP | Kola Peninsula |
| O | color of sea anemones, orange |
| OG | color of sea anemones, orange with grey tentacles |
| PCR | polymerase chain reaction |
| PO | color of sea anemones, pale orange |
| PP | color of sea anemones, pale pink |
| PPR | color of sea anemones, pale pink with red stripes |
| R | color of sea anemones, red |
| RB | color of sea anemones, red–blue blotched. |
Appendix A. Ecological and Morphological Characters Used in the Phylogenetic Analysis
- Habitat preference: rocks (0), sand (1).
- Occurrence on shells: absent (0), present (1).
- Buried in sand: no (0), yes (1).
- Intertidal zone: not inhabited (0), inhabited (1).
- Hermaphroditism: not observed (0), observed (1).
- Dioecy: not observed (0), observed (1).
- Parthenogenesis: not observed (0), observed (1).
- Asexual reproduction (budding, fission, pedal laceration): not observed (0), observed (1).
- In Arctic waters: absent (0), present (1)
- Brooding: not observed (0), observed (1).
- Symbionts: absent (0), present (1).
- Column diameter: small size <1.0 cm (0), medium size 1.0–5.0 cm (1), large size >5.0 cm (2).
- Column high: <3.0 cm (0), 3.0–8.0 cm (1), >8.0 cm (2).
- Number of tentacles: ≤ 96 (0), >96 (1).
- Spirocysts in tentacles: <50 µm (0), ≥ 50 µm (1).
- Basitrichs in tentacles: <30 µm (0), ≥ 30 µm (1).
- p-mastigophores in tentacles: absent (0), present (1).
- b-mastigophores in tentacles: absent (0), present (1).
- Basitrichs in actinopharynx: <60 µm (0), ≥ 60 µm (1).
- p-mastigophores in actinopharynx: absent (0), present (1).
- p-mastigophores in actinopharynx: <30 µm (0), ≥ 30 µm (1).
- Differences in cnidae between feeding-tentacles and catch-tentacles: not observed (0), observed (1).
- Dense aggregations: not formed (0), formed (1).
- Mutually relationships with gastropods: absent (0), present (1).
- Verrucae: absent (0), nonadhesive (1), adhesive (2).
- Acrorhagi: absent (0), present (1).
- Aggressive behavior: not observed (0), observed (1).
- Color of oral disc: similar to the column (0), different from the column (1).
- The pedal disc: similar in diameter to the column (0), wider than the column (1).
- The tentacles are: the same color as the disc (0), different in color from the disc (1).
- Inhabitance of the littoral zone: absent (0), present (1).
- Tropical seas: not occupied (0), occupied (1).
- Body wall: without adhered particles (0), with adhered particles (sand, gravel, shell) (1).
- Mutual relationships with shrimps: absent (0), present (1).
- Color of the column: pale (0), colorful without stripes or spots (1), colorful with stripes or spots (2).
- Color of the tentacles: pale (0), colorful (1), striped or spotted (2).
- Arrangement of tentacles: scattered (0), one cycle (1), more than two cycles (2).
- Shape of tentacles: conical (0), spherical (1), slender (2).
- In Antarctic waters: absent (0), present (1)
- Color of the oral disc: pale (0), colorful without stripes or spots (1), colorful with stripes or spots (2)
References
- Rodríguez, E.; Barbeitos, M.; Daly, M.; Gusmão, L.C.; Häussermann, V. Toward a natural classification: Phylogeny of acontiate sea anemones (Cnidaria, Anthozoa, Actiniaria). Cladistics 2012, 1, 1–18. [Google Scholar] [CrossRef]
- Geller, J.B.; Walton, E.D. Breaking up and getting together: Evolution of symbiosis and cloning by fission in sea anemones (Genus Anthopleura). Evolution 2001, 55, 1781–1794. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.; Chaudhuri, A.; Gusmão, L.; Rodríguez, E. Phylogenetic relationships among sea anemones (Cnidaria: Anthozoa: Actiniaria). Mol. Phylogenetics Evol. 2008, 48, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Carlgren, O. A survey of the Ptychodactiaria, Corallimorpharia and Actiniaria. Kungl. Sven. Vetensk. Handl. 1949, 1, 1–122. [Google Scholar]
- Schmidt, H. On evolution in the Anthozoa. Proc. Int. Coral Reef. Symp. 1974, 1, 533–560. [Google Scholar]
- Rodríguez, E.; Daly, M. Phylogenetic Relationships among Deep-Sea and Chemosynthetic Sea Anemones: Actinoscyphiidae and Actinostolidae (Actiniaria: Mesomyaria). PLoS ONE 2010, 5, e10958. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.B. Measurements of cnidae from sea anemones (Cnidaria: Actiniaria), II: Further studies of differences amongst sample means and their taxonomic relevance. Sci. Mar. 1998, 62, 361–372. [Google Scholar] [CrossRef]
- Schmidt, H. Die Nesselkapseln der Aktinien und ihre differentialdiagnostische Bedeutung. Helgoländer Wiss. Meeresunters. 1969, 19, 284–317. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, E.; Daly, M.; Fautin, D.G. Order Actiniaria. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa 2007, 1668, 131–138. [Google Scholar]
- Kaliszewicz, A.; Panteleeva, N.; Oleiniczak, I.; Boniecki, P.; Sawicki, M. Internal brooding affects the spatial structure of intertidal sea anemones in the Arctic-boreal region. Polar Biol. 2012, 35, 1911–1919. [Google Scholar] [CrossRef]
- Sanamyan, N.P.; Sanamyan, K.E. The genera Urticina and Cribinopsis (Anthozoa: Actiniaria) from the north-western Pacific. J. Nat. Hist. 2006, 40, 359–393. [Google Scholar] [CrossRef]
- Schama, R.; Solé-Cava, A.M.; Thorpe, J.P. Genetic divergence between east and west Atlantic populations of Actinia spp. sea anemones (Cnidaria: Actiniidae). Mar. Biol. 2005, 146, 435–443. [Google Scholar] [CrossRef]
- Chomsky, O.; Douek, J.; Chadwick, N.E.; Dubinsky, Z.; Rinkevich, B. Biological and Population—Genetic aspects of the sea anemone Actinia equina (Cnidaria: Anthozoa) along the Mediterranean coast of Israel. J. Exp. Mar. Biol. Ecol. 2009, 375, 16–20. [Google Scholar] [CrossRef]
- Cunningham, C.W.; Buss, L.W. Molecular evidence for multiple episodes of paedomorphosis in the family Hydractiniidae. Biochem. Syst. Ecol. 1993, 21, 57–69. [Google Scholar] [CrossRef]
- Littlewood, D.T. Molecular phylogenetics of cupped oysters based on partial 28S rRNA gene sequence. Mol. Phylog. Evol. 1994, 3, 221–229. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl. Acids. Res. 1997, 24, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Carlgren, O. The Danish Ingolf-Expedition; Actiniaria; Bianco Luno: Copenhagen, Denmark, 1921. [Google Scholar]
- Kanaev, I.I. Gidra (in Russian); RAS SSRR: Moscow, Russia, 1952. [Google Scholar]
- Siebert, A.E. A description of the Sea Anemone Stomphia didemon sp. nov. and its development. Pac. Sci. 1973, 27, 363–376. [Google Scholar]
- Fautin, D.G. More Antarctic and Subantarctic sea anemones (Coelenterata: Coralliomorpharia and Actiniaria). Biology of the Antarctic Seas XVI. Antarct. Res. Ser. 1984, 41, 1–42. [Google Scholar]
- Riemann-Zürneck, K. Taxonomy and ecological aspects of the subarctic sea anemones Hormathia digitata, Hormathia nodosa and Allantactic parasitica (Coelenterata, Actiniaria). Ophelia 1994, 39, 197–224. [Google Scholar] [CrossRef]
- Daly, M.; den Hartog, J.C. Taxonomy, circumscription, and usage in Anthopleura (Cnidaria: Anthozoa: Actiniaria) from the Gulf of Mexico and Caribbean. Bull. Mar. Sci. 2004, 74, 401–421. [Google Scholar]
- Yanagi, K.; Daly, M. The hermaphroditic sea anemone Anthopleura atodai n. sp. (Anthozoa: Actiniaria: Actiniidae) from Japan, with a redescription of A. hermaphroditica. Proc. Biol. Soc. Wash. 2004, 117, 408–422. [Google Scholar]
- Acuña, F.H.; Alvarado, J.; Garese, A.; Cortés, J. First record of the sea anemone Anthopleura nigrescens (Cnidaria: Actiniaria: Actiniidae) on the Pacific coast of Central America. Mar. Biodivers. Rec. 2012, 5, e24. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Catalano, S.A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Nixon, K.C. WinClada; Ithaca, NY, USA,, 2002. [Google Scholar]
- Spaulding, J.G. The development and brooding behavior of the sea anemone Cribrinopsis williamsi Carlgren. Am. Zool. 1971, 11, 521. [Google Scholar]
- Larson, P. Brooding sea anemones (Cnidaria: Anthozoa: Actiniaria): Paragons of diversity in mode, morphology, and maternity. Invertebr. Biol. 2017, 136, 92–112. [Google Scholar] [CrossRef]
- Bocharova, E.S. Reproductive biology and genetic diversity of the sea anemone Aulactinia stella (Verrill, 1864). Hydrobiologia 2015, 759, 27–38. [Google Scholar] [CrossRef]
- Daly, M.; Crowley, L.M.; Larson, P.; Rodríguez, E.; Heestand Saucier, E.; Fautin, D.G. Anthopleura and the phylogeny of Actinioidea (Cnidaria: Anthozoa: Actiniaria). Org. Divers. Evol. 2017, 17, 545–564. [Google Scholar] [CrossRef]
- Daly, M.; Gusmão, L.C.; Reft, A.J.; Rodríguez, E. Phylogenetic signal in mitochondrial and nuclear markers in sea anemones (Cnidaria, Actiniaria). Integr. Compar. Biol. 2010, 50, 371–388. [Google Scholar] [CrossRef] [Green Version]
- González-Muñoz, R.; Simões, N.; Mascaró, M.; Tello-Musi, J.L. Morphological and molecular variability of the sea anemone Phymanthus crucifer (Cnidaria, Anthozoa, Actiniaria, Actinoidea). J. Mar. Biol. Assoc. 2015, 95, 69–79. [Google Scholar] [CrossRef]
- Dolan, E.; Tyler, P.A.; Yesson, C.; Rogers, A.D. Phylogeny and systematics of deep-sea sea pens (Anthozoa: Octocorallia: Pennatulacea). Mol. Phylogenet. Evol. 2013, 69, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Shearer, T.L.; Van Oppen, M.J.; Romano, S.L.; Wörheide, G. Slow mitochondrial DNA sequence evolution in the Anthozoa (Cnidaria). Mol. Ecol. 2002, 11, 2475–2487. [Google Scholar] [CrossRef] [PubMed]
- Brugler, M.R.; Opresko, D.M.; France, S. The evolutionary history of the order Antipatharia (Cnidaria: Anthozoa: Hexacorallia) as inferred from mitochondrial and nuclear DNA: Implications for black coral taxonomy and systematics. Zool. J. Linn. Soc. 2013, 169, 312–361. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Family | Species | Locality | Color Pattern | GenBank Accession |
|---|---|---|---|---|---|
| Mitochondrial 16S rDNA | |||||
| Anthozoa | Actiniidae | Urticina grebelnyi | EK | MW491942 * | |
| Urticina coriacea | EU190797 | ||||
| KT852114 | |||||
| Urticina crassicornis | KP | RB | MW491943 * | ||
| KP | OG | MW491944 * | |||
| KP | PPR | MW491945 * | |||
| KP | O | MW491946 * | |||
| KP | RB | MW491947 * | |||
| KP | PP | MW491948 * | |||
| KP | O | MW491949 * | |||
| EK | B | MW491950 * | |||
| KP | RB | MW491951 * | |||
| KP | R | MW491952 * | |||
| KT85205 | |||||
| JQ92744 | |||||
| Cribrinopsis albopunctata | EK | PP | MW491953 * | ||
| EK | R | MW491954 * | |||
| Cribrinopsis olegi | EK | MW491955 * | |||
| Cribrinopsis similis | KP | - | MW491956* | ||
| Aulactinia incubans | KT852080 | ||||
| Aulactinia stella | KP | G | MW491959 * | ||
| EK | - | MW491958 * | |||
| KP | PO | MW491957 * | |||
| Aulactinia verrucosa | EU190766 | ||||
| Anthopleura atodai | KT852055 | ||||
| Anthopleura elegantissima | AEU40292 | ||||
| Anthopleura krebsi | EU190758 | ||||
| KY789339 | |||||
| Anthopleura nigrescens | KY789340 | ||||
| Anthopleura orientalis | EK | MW491960 * | |||
| Actinostolidae | Stomphia didemon | EU190795 | |||
| Stomphia selaginella | GU473298 | ||||
| Hormathiidae | Allantactis parasitica | FJ489420 | |||
| Hormathia pectinata | FJ489430 | ||||
| Metridiidae | Metridium senile | AY345876 | |||
| EU190786 |
| Class | Family | Species | Locality | Color Pattern | GenBank Accession |
|---|---|---|---|---|---|
| Nuclear 28S rDNA | |||||
| Anthozoa | Actiniidae | Urticina grebelnyi | EK | MW491984 * | |
| Urticina coriacea | KT852266 | ||||
| Urticina crassicornis | KP | PPR | MW491985 * | ||
| KP | RB | MW491986 * | |||
| KP | O | MW491987 * | |||
| KP | PPR | MW491988 * | |||
| FJL | - | MW491989 * | |||
| EK | - | MW491990 * | |||
| Cribrinopsis albopunctata | EK | R | MW491991 * | ||
| EK | PP | MW491992 * | |||
| Cribrinopsis similis | KP | R | MW491993 * | ||
| KP | B | MW491994 * | |||
| KP | - | MW491995 * | |||
| Aulactinia incubans | KT852256 | ||||
| Aulactinia stella | EK | - | MW491996 * | ||
| KP | G | MW491997 * | |||
| KP | PO | MW491998 * | |||
| FJL | - | MW491999 * | |||
| Aulactinia verrucosa | EU190812 | ||||
| KJ483084 | |||||
| KT852250 | |||||
| Anthopleura atodai | KT852247 | ||||
| Anthopleura elegantissima | EU190801 | ||||
| KJ483104 | |||||
| Anthopleura krebsi | EU190804 | ||||
| Anthopleura nigrescens | KY789375 | ||||
| Anthopleura orientalis | EK | MW492000 * | |||
| Actinostolidae | Stomphia didemon | EU190837 | |||
| Stomphia selaginella | GU473331 | ||||
| Hormathiidae | Allantactis parasitica | FJL | MW492001 * | ||
| MW492002 * | |||||
| FJL | FJ489454 | ||||
| KJ483056 | |||||
| Hormathia pectinata | FJ489465 | ||||
| Metridiidae | Metridium senile | EU190829 | |||
| JF833000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaliszewicz, A.; Panteleeva, N.; Żmuda-Baranowska, M.; Szawaryn, K.; Olejniczak, I.; Boniecki, P.; Grebelnyi, S.D.; Kabzińska, D.; Romanowski, J.; Maciaszek, R.; et al. Phylogenetic Relatedness within the Internally Brooding Sea Anemones from the Arctic-Boreal Region. Biology 2021, 10, 81. https://doi.org/10.3390/biology10020081
Kaliszewicz A, Panteleeva N, Żmuda-Baranowska M, Szawaryn K, Olejniczak I, Boniecki P, Grebelnyi SD, Kabzińska D, Romanowski J, Maciaszek R, et al. Phylogenetic Relatedness within the Internally Brooding Sea Anemones from the Arctic-Boreal Region. Biology. 2021; 10(2):81. https://doi.org/10.3390/biology10020081
Chicago/Turabian StyleKaliszewicz, Anita, Ninel Panteleeva, Magdalena Żmuda-Baranowska, Karol Szawaryn, Izabella Olejniczak, Paweł Boniecki, Sergey D. Grebelnyi, Dagmara Kabzińska, Jerzy Romanowski, Rafał Maciaszek, and et al. 2021. "Phylogenetic Relatedness within the Internally Brooding Sea Anemones from the Arctic-Boreal Region" Biology 10, no. 2: 81. https://doi.org/10.3390/biology10020081
APA StyleKaliszewicz, A., Panteleeva, N., Żmuda-Baranowska, M., Szawaryn, K., Olejniczak, I., Boniecki, P., Grebelnyi, S. D., Kabzińska, D., Romanowski, J., Maciaszek, R., Górska, E. B., & Zawadzka-Sieradzka, J. (2021). Phylogenetic Relatedness within the Internally Brooding Sea Anemones from the Arctic-Boreal Region. Biology, 10(2), 81. https://doi.org/10.3390/biology10020081