
MLST-Based Analysis and Antimicrobial Resistance of Staphylococcus epidermidis from Cases of Sheep Mastitis in Greece

 , , ,
, , , 
 ,
, 
 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection Regarding S. epidermidis from the MLST Database and Analysis
2.1.1. Data Collection
2.1.2. Statistical Analysis
2.2. Evaluation of S. epidermidis Isolates Recovered in Greece
2.2.1. Bacterial Isolation
2.2.2. MLST and Phylogenetic Analysis
2.2.3. Susceptibility Testing to Antimicrobial Agents
2.2.4. Data Management and Analysis
3. Results
3.1. Details of S. epidermidis in the MLST Database
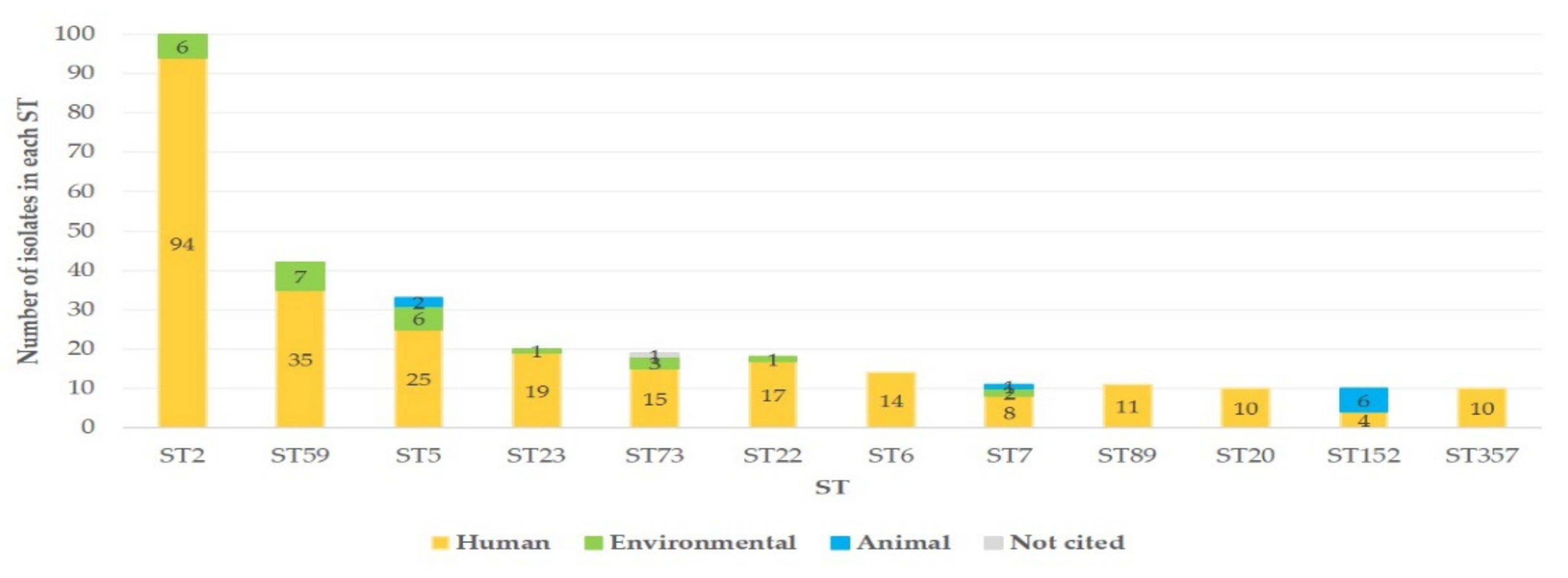
3.1.1. Details of All S. epidermidis Isolates in the Database
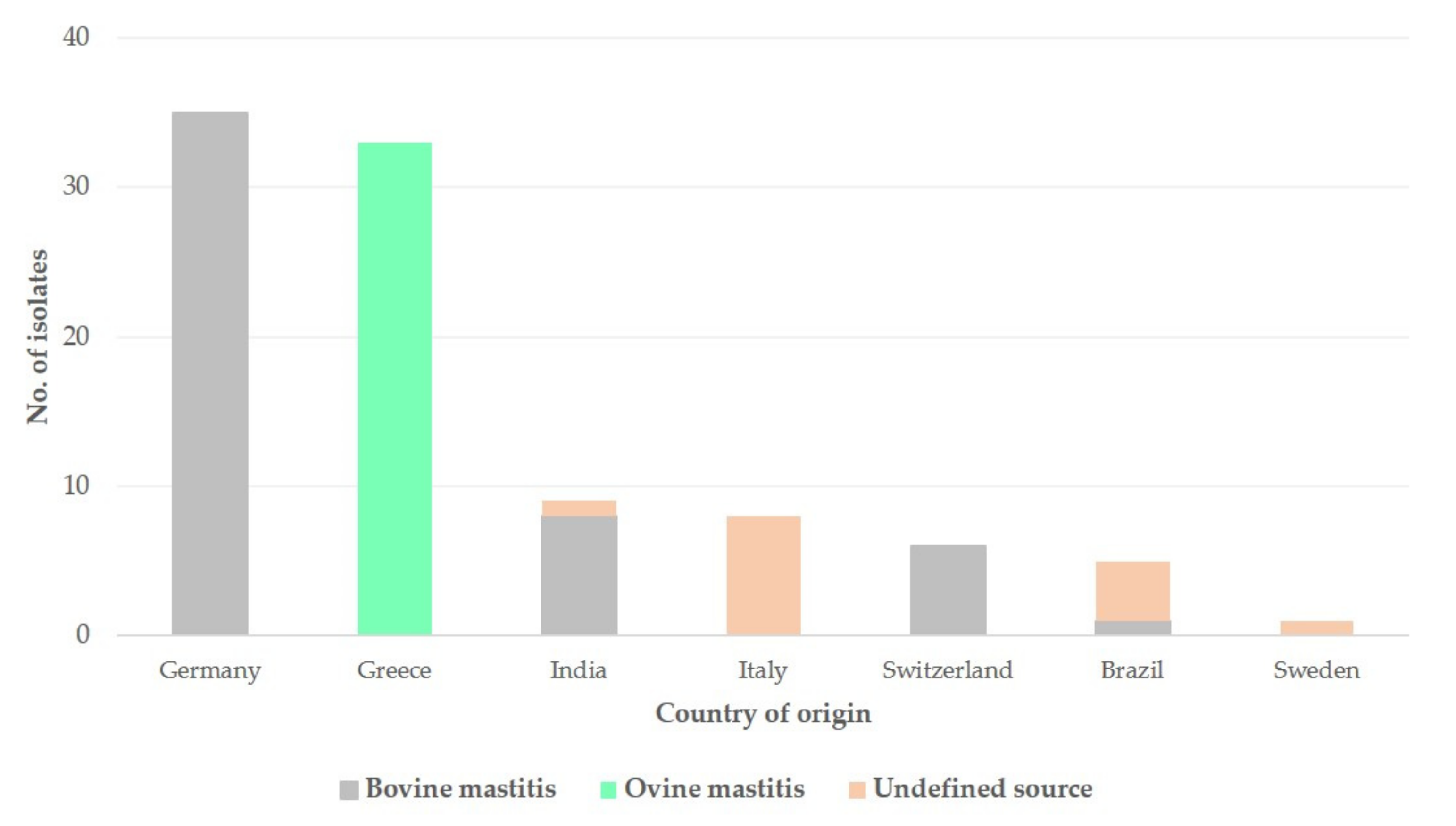
3.1.2. S. epidermidis Isolates Recovered from Mastitis
3.2. S. epidermidis Isolates Recovered from Ovine Mastitis in Greece
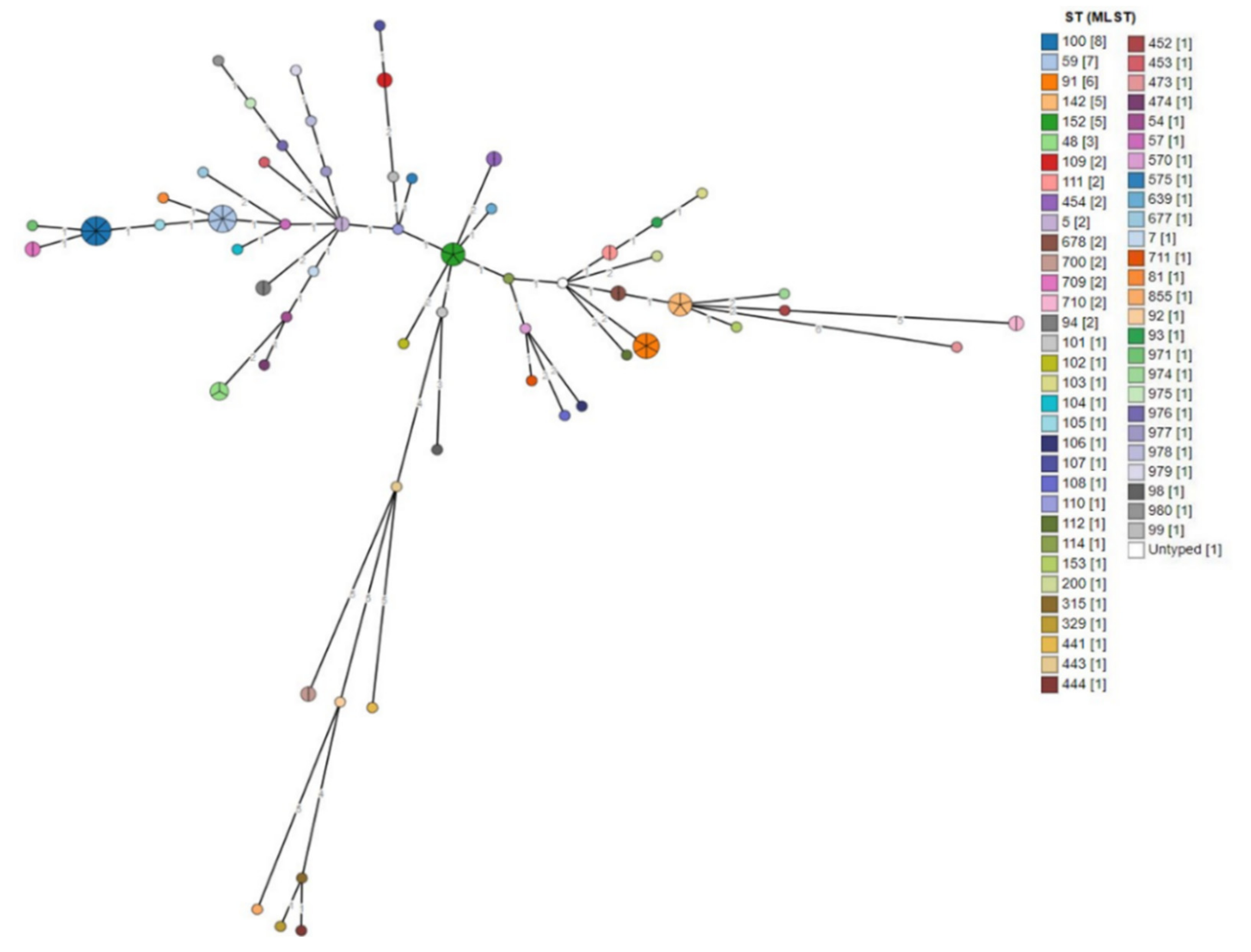
3.2.1. MLST
3.2.2. Resistance to Antimicrobial Agents
3.2.3. Associations between MLST and Resistance to Antimicrobial Agents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maiden, M.C.J.; Bygraves, J.A.; Feil, E.; Morelli, G.; Russell, J.E.; Urwin, R.; Zhang, Q.; Zhou, J.J.; Zurth, K.; Caugant, D.A.; et al. Multilocus sequence typing: A portable approach to the identification of clones within populations of pathogenic microorganisms. Proc. Natl. Acad. Sci. USA 1998, 95, 3140–3145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enright, M.C.; Spratt, B.G. A multilocus sequence typing scheme for Streptococcus pneumoniae: Identification of clones associated with serious invasive disease. Microbiol. UK 1998, 144, 3049–3060. [Google Scholar] [CrossRef] [Green Version]
- Feil, E.J.; Maiden, M.C.J.; Achtman, M.; Spratt, B.G. The relative contributions of recombination and mutation to the divergence of clones of Neisseria meningitides. Molec. Biol. Evol. 1999, 16, 1496–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloos, W.E. Natural populations of the genus Staphylococcus. Ann. Rev. Microbiol. 1980, 34, 559–592. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus epidermidis—The “accidental” pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M. Staphylococcus colonization of the skin and antimicrobial peptides. Exp. Rev. Dermatol. 2010, 5, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Paragioudaki, M.; Stamouli, V.; Kolonitsiou, F.; Anastassiou, E.D.; Dimitracopoulos, G.; Spiliopoulou, I. Intravenous catheter infections associated with bacteraemia: A 2-year study in a University Hospital. Clin. Microbiol. Inf. 2004, 10, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Papadimitriou-Olivgeris, M.; Kolonitsiou, F.; Karamouzos, V.; Tsilipounidaki, K.; Nikolopoulou, A.; Fligou, F.; Marangos, M.; Petinaki, E.; Spiliopoulou, I. Molecular characteristics and predictors of mortality among Gram-positive bacteria isolated from bloodstream infections in critically ill patients during a 5-year period (2012–2016). Eur. J. Clin. Microbiol. Infect. Dis. 2020, 5, 863–869. [Google Scholar] [CrossRef]
- Daskalaki, E.; Koukoulaki, M.; Bakalis, A.; Papastamopoulos, V.; Belesiotou, E.; Perivolioti, E.; Skoutelis, A.; Drakopoulos, S. Blood stream infections in renal transplant recipients: A single-center study. Transpl. Proc. 2014, 46, 3191–3194. [Google Scholar] [CrossRef]
- Giormezis, N.; Kolonitsiou, F.; Foka, F.; Drougka, E.; Liakopoulos, A.; Makri, A.; Papanastasiou, A.D.; Vogiatzi, A.; Dimitriou, G.; Marangos, M.; et al. Coagulase-negative staphylococcal bloodstream and prosthetic-device-associated infections: The role of biofilm formation and distribution of adhesin and toxin genes. J. Med. Microbiol. 2014, 63, 1500–1508. [Google Scholar] [CrossRef]
- Dimitriou, G.; Fouzas, S.; Giormezis, N.; Giannakopoulos, I.; Tzifas, S.; Foka, A.; Anastassiou, D.E.; Spiliopoulou, I.; Mantagos, S. Clinical and microbiological profile of persistent coagulase-negative staphylococcal bacteraemia in neonates. Clin. Microbiol. Inf. 2011, 17, 1684–1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dotis, J.; Printza, N.; Papachristou, F. Peritonitis due to uncommon gram-positive pathogens in children undergoing peritoneal dialysis. Hippokratia 2012, 16, 267–268. [Google Scholar]
- Gkentzi, D.; Kolyva, S.; Spiliopoulou, I.; Marangos, M.; Dimitriou, G. Treatment options for persistent coagulase negative staphylococcal bacteremia in neonates. Curr. Pediatr. Rev. 2016, 12, 199–208. [Google Scholar] [CrossRef]
- Loupa, C.; Mavroidi, N.; Boutsikakis, I.; Paniara, O.; Deligarou, O.; Manoli, H.; Saroglou, G. Infective endocarditis in Greece: A changing profile. Epidemiological, microbiological and therapeutic data. Clin. Microbiol. Infect. 2004, 10, 556–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detorakis, E.T.; Siganos, D.S.; Houlakis, V.M.; Kozobolis, V.P.; Pallikaris, I.G. Microbiological examination of bandage soft contact lenses used in laser refractive surgery. J. Refract. Surg. 1998, 14, 631–635. [Google Scholar]
- Mela, E.K.; Drimtzias, E.G.; Christofidou, M.K.; Filos, K.S.; Anastassiou, E.D.; Gartaganis, S.P. Ocular surface bacterial colonisation in sedated intensive care unit patients. Anaesth. Intensive Care 2010, 38, 190–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmberg, O. Staphylococcus epidermidis isolated from bovine milk. Biochemical properties, phage sensitivity and pathogenicity for the udder. Acta Vet. Scand. 1973, 14, 1–144. [Google Scholar]
- Fthenakis, G.C.; Jones, J.E.T. The effect of inoculation of coagulase-negative staphylococci into the ovine mammary gland. J. Comp. Pathol. 1990, 102, 211–219. [Google Scholar] [CrossRef]
- Abbondio, M.; Fois, I.; Longheu, C.; Azara, E.; Tola, S. Biofilm production, quorum sensing system and analysis of virulence factors of Staphylococcus epidermidis collected from sheep milk samples. Small Rumin. Res. 2019, 174, 83–87. [Google Scholar] [CrossRef]
- Vasileiou, N.G.C.; Chatzopoulos, D.C.; Sarrou, S.; Fragkou, I.A.; Katsafadou, A.I.; Mavrogianni, V.S.; Petinaki, E.; Fthenakis, G.C. Role of staphylococci in mastitis in sheep. J. Dairy Res. 2019, 86, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Kotzekidou, P. Identification of staphylococci and micrococci isolated from an intermediate moisture meat product. J. Food Sci. 1992, 57, 249–251. [Google Scholar] [CrossRef]
- Abrahim, A.; Sergelidis, D.; Kirkoudis, I.; Anagnostou, V.; Kaitsa-Tsiopoulou, E.; Kazila, P.; Papa, A. Isolation and antimicrobial resistance of Staphylococcus spp. in freshwater fish and Greek marketplaces. J. Aquat. Food Prod. Technol. 2010, 19, 93–102. [Google Scholar] [CrossRef]
- Sergelidis, D.; Abrahim, A.; Papadopoulos, T.; Soultos, N.; Martziou, E.; Koulourida, V.; Govaris, A.; Pexara, A.; Zdragas, A.; Papa, A. Isolation of methicillin-resistant Staphylococcus spp. from ready-to-eat fish products. Lett. Appl. Microbiol. 2014, 59, 500–506. [Google Scholar] [CrossRef]
- Turchi, B.; Bertelloni, F.; Marzoli, F.; Cerri, D.; Tola, S.; Azara, E.; Longheu, C.M.; Tassi, R.; Schiavo, M.; Cilia, G.; et al. Coagulase negative staphylococci from ovine milk: Genotypic and phenotypic characterization of susceptibility to antibiotics, disinfectants and biofilm production. Small Rumin. Res. 2020, 183, 106030. [Google Scholar] [CrossRef]
- Kalogridou-Vasileiadou, D. Mastitis related pathogens in milk. Small Rumin. Res. 1991, 4, 203–212. [Google Scholar] [CrossRef]
- Fthenakis, G.C. Prevalence and aetiology of subclinical mastitis in ewes in southern Greece. Small Rumin. Res. 1994, 13, 293–300. [Google Scholar] [CrossRef]
- Kiossis, E.; Brozos, C.N.; Petridou, E.; Boscos, C. Program for the control of subclinical mastitis in dairy Chios breed ewes during lactation. Small Rumin. Res. 2007, 73, 194–199. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific opinion on the welfare risks related to the farming of sheep for wool, meat and milk production. EFSA J. 2014, 12, 3933–4060. [Google Scholar]
- Vasileiou, N.G.C.; Cripps, P.J.; Ioannidi, K.S.; Chatzopoulos, D.C.; Gougoulis, D.A.; Sarrou, S.; Orfanou, D.C.; Politis, A.; Calvo Gonzalez-Valerio, T.; Argyros, S.; et al. Extensive countrywide field investigation of subclinical mastitis in sheep in Greece. J. Dairy Sci. 2018, 101, 7297–7310. [Google Scholar] [CrossRef] [Green Version]
- Saratsis, P.; Alexopoulos, C.; Tzora, A.; Fthenakis, G.C. The effect of experimentally induced subclinical mastitis on the milk yield of dairy ewes. Small. Rumin. Res. 1999, 32, 205–209. [Google Scholar] [CrossRef]
- Lianou, D.T.; Fthenakis, G.C. Scientometrics approach to research in ovine mastitis from 1970 to 2019 (with a complete list of relevant literature references). Pathogens 2020, 9, 585. [Google Scholar] [CrossRef]
- Zhou, Ζ.; Alikhan, N.F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. GrapeTree: Visualization of core genomic relationships among 100,000 bacterial pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef] [Green Version]
- Vasileiou, N.G.C.; Chatzopoulos, D.C.; Cripps, P.J.; Ioannidi, K.S.; Gougoulis, D.A.; Chouzouris, T.M.; Lianou, D.T.; Calvo Gonzalez-Valerio, T.; Guix Vallverdu, R.; Argyros, S.; et al. Evaluation of efficacy of a biofilm-embedded bacteria-based vaccine against staphylococcal mastitis in sheep—A randomized, placebo-controlled field study. J. Dairy Sci. 2019, 102, 9328–9344. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.C.; Vargas, M.R.; Miragaia, M.; Peacock, S.J.; Archer, G.L.; Enright, M.C. Improved multilocus sequence typing scheme for Staphylococcus epidermidis. J. Clin. Microbiol. 2007, 45, 616–619. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Meth. 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Petinaki, E.; Kontos, F.; Miriagou, V.; Maniati, M.; Hatzi, F.; Maniatis, A.N. Survey of methicillin-resistant coagulase-negative staphylococci in the hospitals of central Greece. Int. J. Antimicrob. Agents 2001, 18, 563–566. [Google Scholar] [CrossRef]
- Fragkou, I.A.; Boscos, C.M.; Fthenakis, G.C. Diagnosis of clinical or subclinical mastitis in ewes. Small Rumin. Res. 2014, 118, 86–92. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pybus, O.G.; Rambaut, A. Evolutionary analysis of the dynamics of viral infectious disease. Nat. Rev. Genet. 2009, 10, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Belén, A.; Pavón, I.; Maiden, M.C.J. Multilocus sequence typing. Methods Mol. Biol. 2009, 551, 129–140. [Google Scholar]
- Watts, J.L.; Owens, W.E. Prevalence of staphylococcal species in four dairy 732 herds. Res. Vet. Sci. 1989, 46, 1–4. [Google Scholar] [CrossRef]
- Thorberg, B.M.; Kuhn, I.; Aarestrup, F.M.; Brandstrom, B.; Jonsson, P.; Danielsson-Tham, M.L. Pheno- and genotyping of Staphylococcus epidermidis isolated from bovine milk and human skin. Vet. Microbiol. 2006, 115, 163–172. [Google Scholar] [CrossRef]
- Delgado, D.; Arroyo, R.; Jiménez, E.; Marín, M.L.; del Campo, R.; Fernández, L.; Rodríguez, J.M. Staphylococcus epidermidis strains isolated from breast milk of women suffering infectious mastitis: Potential virulence traits and resistance to antibiotics. BMC Microbiol. 2009, 9, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo, R.; Martin, V.; Maldonado, A.; Jimenez, E.; Fernandez, L.; Rodrıguez, J.M. Treatment of infectious mastitis during lactation: Antibiotics versus oral administration of lactobacilli isolated from breast milk. Clin. Inf. Dis. 2010, 50, 1551–1558. [Google Scholar] [CrossRef] [Green Version]
- Miragaia, M.; Thomas, J.C.; Couto, I.; Enright, M.C.; de Lencastre, H. Inferring a population structure for Staphylococcus epidermidis from Multilocus Sequence Typing Data. J. Bacteriol. 2007, 189, 2540–2552. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Satorius, A.E.; Raff, M.R.; Rivera, A.; Newton, D.W.; Younger, J.G. Multilocus Sequence Typing for interpreting blood isolates of Staphylococcus epidermidis, interdisciplinary perspectives on infectious diseases. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 4. [Google Scholar] [CrossRef] [Green Version]
- Howie, B.; Marchini, J.; Stephens, M. Genotype imputation with thousands of genomes. Genes Genomes Genet. 2011, 1, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Onni, T.; Sanna, G.; Larsen, J.; Tola, S. Antimicrobial susceptibilities and population structure of Staphylococcus epidermidis associated with ovine mastitis. Vet. Microbiol. 2011, 148, 45–50. [Google Scholar] [CrossRef]
- Karakonstantis, S.; Kalemaki, D. Antimicrobial overuse and misuse in the community in Greece and link to antimicrobial resistance using methicillin-resistant S. aureus as an example. J. Infect. Public Health 2019, 12, 460–464. [Google Scholar] [CrossRef]
- Koulenti, D.; Fragkou, P.C.; Tsiodras, S. Editorial for Special Issue ‘Multidrug-Resistant Pathogens’. Microorganisms 2020, 8, 1383. [Google Scholar] [CrossRef]
- Memon, J.; Yang, Y.; Kashifa, J.; Yaqoob, M.; Buriroa, R.; Soomroa, J.; Liping, W.; Hongjie, F. Genotypes, virulence factors and antimicrobial resistance genes of Staphylococcus aureus isolated in bovine subclinical mastitis from Eastern China. Pak. Vet. J. 2013, 33, 486–491. [Google Scholar]
- Mavrogianni, V.S.; Cripps, P.J.; Tzora, A.; Skoufos, I.; Fthenakis, G.C. Effects of hand milking on the bacterial flora of mammary gland and teat duct of ewes. J. Dairy Res. 2006, 73, 353–356. [Google Scholar] [CrossRef]
- Vasileiou, N.G.C.; Chatzopoulos, D.C.; Gougoulis, D.A.; Sarrou, S.; Katsafadou, A.I.; Spyrou, V.; Mavrogianni, V.S.; Petinaki, E.; Fthenakis, G.C. Slime-producing staphylococci as causal agents of subclinical mastitis in sheep. Vet. Microbiol. 2018, 224, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Marco Melero, J.C. Mastitis in Laxta Breed Sheep: Epidemiology, Diagnosis and Control. Ph.D. Thesis, University of Zaragoza, Zaragoza, Spain, 1994. [Google Scholar]
- Albenzio, M.; Taibi, L.; Caroprese, M.; De Rosa, G.; Muscio, A.; Sevi, A. Immune response, udder health and productive traits of machine milked and suckling ewes. Small Rumin. Res. 2003, 48, 189–200. [Google Scholar] [CrossRef]
- Mavrogianni, V.S.; Menzies, P.I.; Fragkou, I.A.; Fthenakis, G.C. Principles of mastitis treatment in sheep and goats. Vet. Clin. N. Am. Food Anim. Pract. 2011, 27, 115–120. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alleles | arcC | aroE | gtr | mutS | pyrR | tpiA | yqiL |
|---|---|---|---|---|---|---|---|
| Total no. of alleles detected | 72 | 67 | 79 | 43 | 58 | 63 | 68 |
| Most frequent allele | 1 | 1 | 2 | 2 | 2 | 1 | 1 |
| Isolates (n) | 623 | 994 | 612 | 596 | 511 | 939 | 758 |
| Second-most frequent allele | 7 | 2 | 1 | 1 | 4 | 16 | 4 |
| Isolates (n) | 223 | 104 | 312 | 302 | 277 | 139 | 101 |
| arcC | yqiL | ||||
|---|---|---|---|---|---|
| Allele | Isolates from Bovine Mastitis (n) | Isolates from Ovine Mastitis (n) | Allele | Isolates from Bovine Mastitis (n) | Isolates from Ovine Mastitis (n) |
| 1 | 33 | 9 | 1 | 21 | 29 |
| 2 | 8 | 18 | 3 | 0 | 1 |
| 7 | 3 | 0 | 4 | 3 | 0 |
| 8 | 0 | 3 | 7 | 7 | 0 |
| 12 | 4 | 1 | 8 | 3 | 1 |
| 19 | 1 | 0 | 10 | 7 | 0 |
| 28 | 1 | 0 | 11 | 0 | 2 |
| 65 | 0 | 2 | 18 | 6 | 0 |
| 19 | 1 | 0 | |||
| 21 | 1 | 0 | |||
| 49 | 1 | 0 | |||
| aroE | tpiA | ||||
|---|---|---|---|---|---|
| Allele | Isolates from Ovine Mastitis (n) | Isolates from Mammary Carriage (n) | Allele | Isolates from Ovine Mastitis (n) | Isolates from Mammary Carriage (n) |
| 1 | 25 | 2 | 1 | 16 | 3 |
| 2 | 1 | 0 | 5 | 1 | 0 |
| 3 | 0 | 1 | 16 | 2 | 0 |
| 13 | 0 | 1 | 19 | 0 | 1 |
| 25 | 1 | 0 | 48 | 1 | 0 |
| 51 | 1 | 0 | 2 | ||
| 59 | 1 | 0 | |||
| arc | pyrR | tpiA | ||||||
|---|---|---|---|---|---|---|---|---|
| Allele | Isolates from Ovine Mastitis (n) | Isolates of Human Origin (n) | Allele | Isolates from Ovine Mastitis (n) | Isolates of Human Origin (n) | Allele | Isolates from Ovine Mastitis (n) | Isolates of Human Origin (n) |
| 1 | 9 | 2 | 1 | 0 | 1 | 1 | 17 | 5 |
| 2 | 18 | 0 | 2 | 18 | 1 | 3 | 0 | 1 |
| 3 | 0 | 1 | 3 | 1 | 2 | 4 | 0 | 1 |
| 7 | 0 | 6 | 4 | 0 | 5 | 5 | 1 | 0 |
| 8 | 3 | 0 | 6 | 1 | 0 | 7 | 0 | 2 |
| 12 | 1 | 0 | 8 | 2 | 0 | 13 | 0 | 1 |
| 22 | 0 | 1 | 9 | 2 | 0 | 16 | 4 | 0 |
| 65 | 2 | 0 | 10 | 0 | 1 | 48 | 2 | 0 |
| 20 | 9 | 0 | ||||||
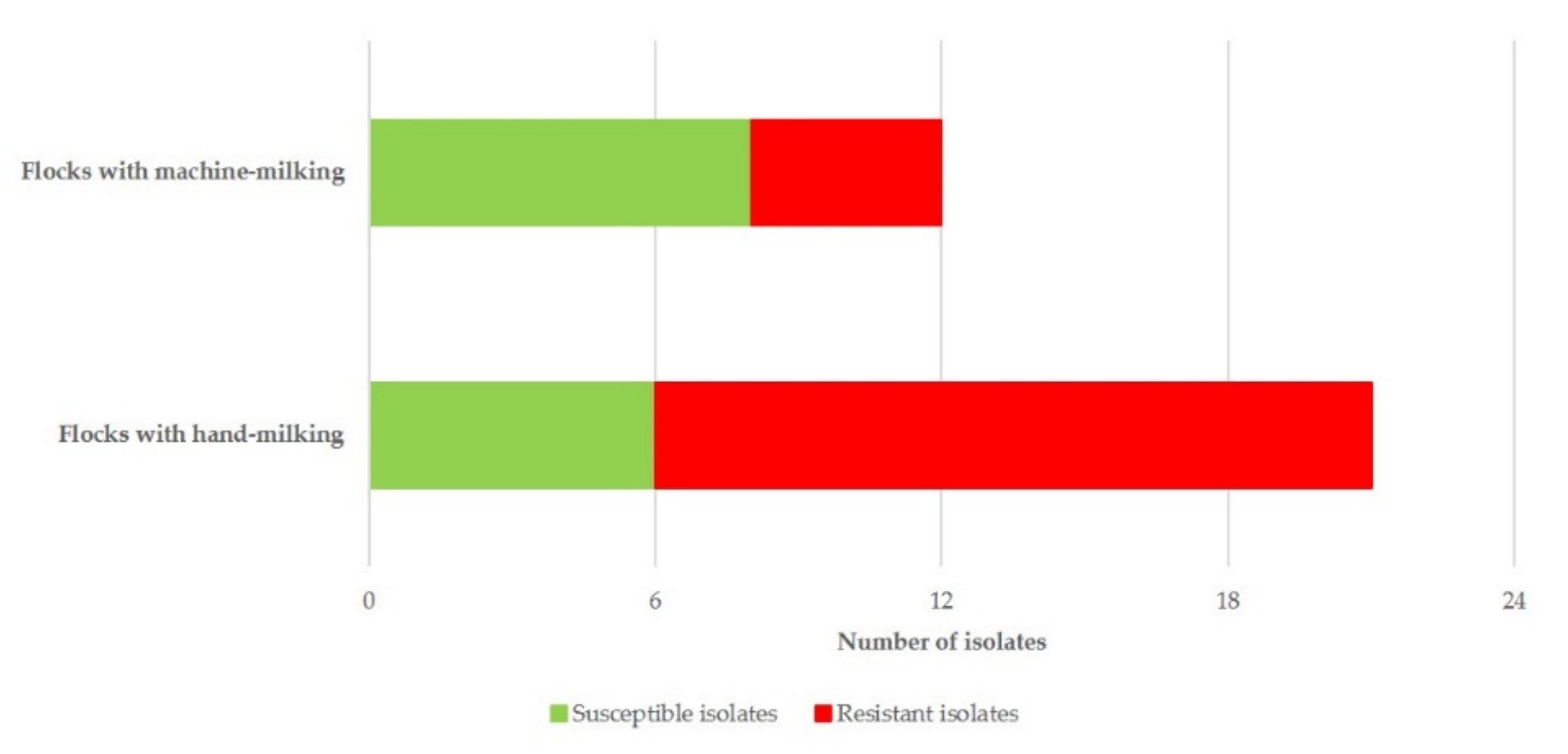
| Milking Techniques Applied in the Flock | Proportion of Flocks in Which Resistant S. epidermidis Were Recovered | Odds Ratio 1 (95%CI 2) | p |
|---|---|---|---|
| Machine-milking (n = 12) | 33.3% | reference | |
| Hand-milking (n = 21) | 71.4% | 5.000 (1.084–23.061) | 0.039 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katsarou, E.I.; Chatzopoulos, D.C.; Giannoulis, T.; Ioannidi, K.S.; Katsafadou, A.I.; Kontou, P.I.; Lianou, D.T.; Mamuris, Z.; Mavrogianni, V.S.; Michael, C.K.; et al. MLST-Based Analysis and Antimicrobial Resistance of Staphylococcus epidermidis from Cases of Sheep Mastitis in Greece. Biology 2021, 10, 170. https://doi.org/10.3390/biology10030170
Katsarou EI, Chatzopoulos DC, Giannoulis T, Ioannidi KS, Katsafadou AI, Kontou PI, Lianou DT, Mamuris Z, Mavrogianni VS, Michael CK, et al. MLST-Based Analysis and Antimicrobial Resistance of Staphylococcus epidermidis from Cases of Sheep Mastitis in Greece. Biology. 2021; 10(3):170. https://doi.org/10.3390/biology10030170
Chicago/Turabian StyleKatsarou, Eleni I., Dimitris C. Chatzopoulos, Themis Giannoulis, Katerina S. Ioannidi, Angeliki I. Katsafadou, Panagiota I. Kontou, Daphne T. Lianou, Zissis Mamuris, Vasia S. Mavrogianni, Charalambia K. Michael, and et al. 2021. "MLST-Based Analysis and Antimicrobial Resistance of Staphylococcus epidermidis from Cases of Sheep Mastitis in Greece" Biology 10, no. 3: 170. https://doi.org/10.3390/biology10030170
APA StyleKatsarou, E. I., Chatzopoulos, D. C., Giannoulis, T., Ioannidi, K. S., Katsafadou, A. I., Kontou, P. I., Lianou, D. T., Mamuris, Z., Mavrogianni, V. S., Michael, C. K., Papadopoulos, E., Petinaki, E., Sarrou, S., Vasileiou, N. G. C., & Fthenakis, G. C. (2021). MLST-Based Analysis and Antimicrobial Resistance of Staphylococcus epidermidis from Cases of Sheep Mastitis in Greece. Biology, 10(3), 170. https://doi.org/10.3390/biology10030170