
Reproduction in Trypanosomatids: Past and Present

 ,
,  ,
,  , and
, and 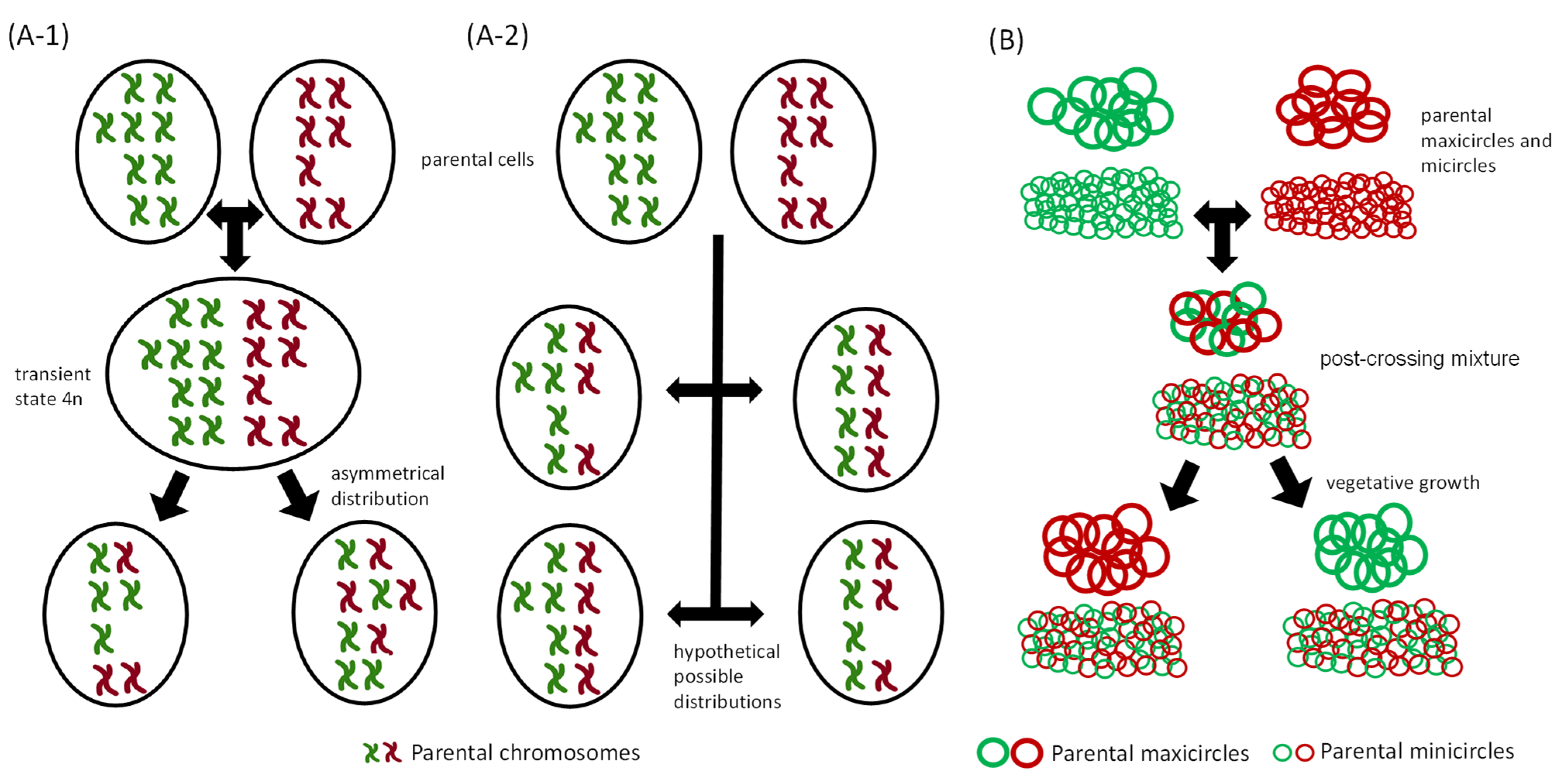{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Clonal Theory or True Sex?
2. Genetic Exchange in Trypanosoma
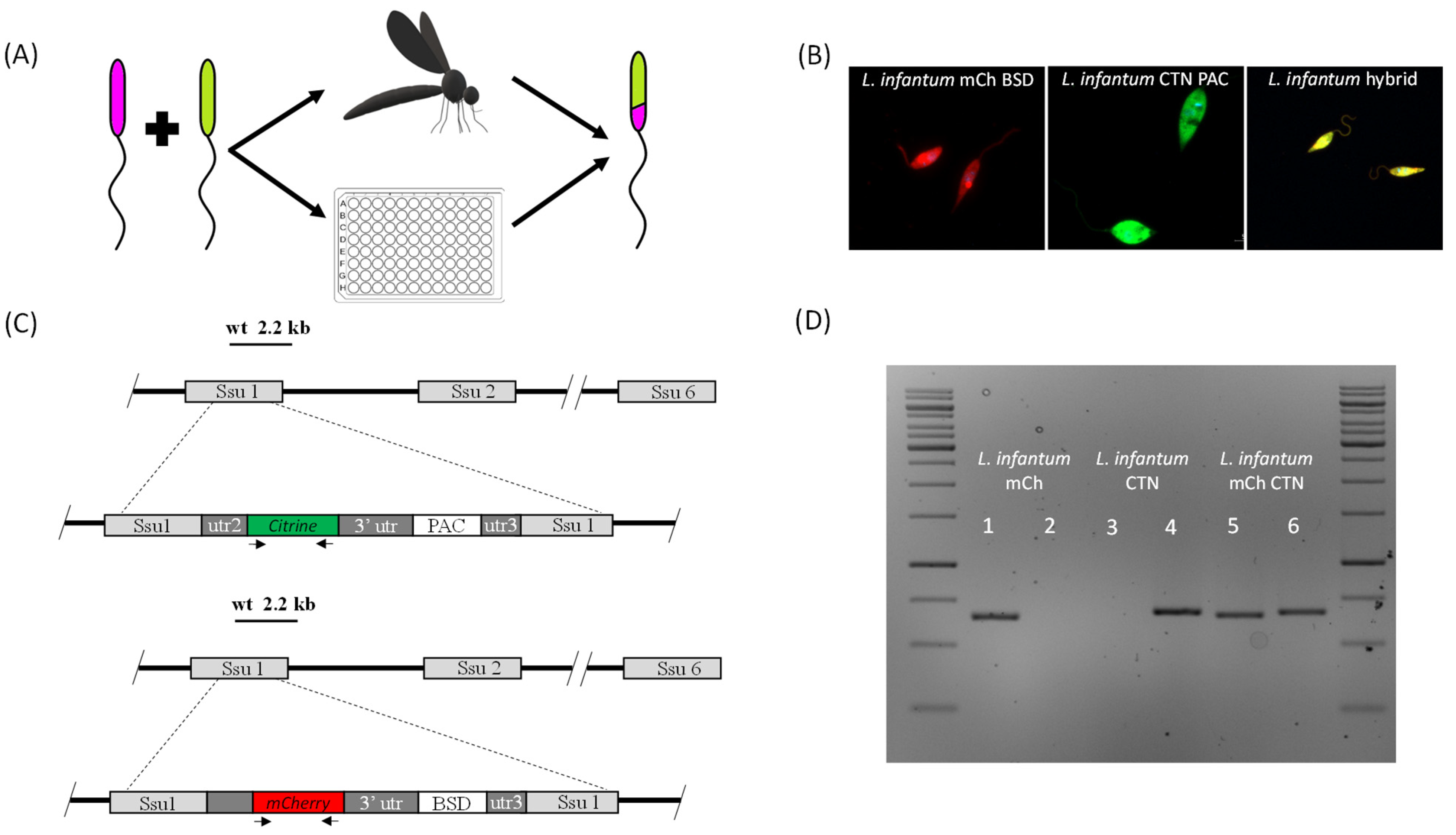
3. Genetic Exchange in Leishmania
4. Genetic Inheritance of kDNA
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Z.; Miao, J.; Cui, L. Gametocytogenesis in malaria parasite: Commitment, development and regulation. Future Microbiol. 2011, 6, 1351–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibayrenc, M.; Kjellberg, F.; Ayala, F.J. A clonal theory of parasitic protozoa: The population structures of Entamoeba, Giardia, Leishmania, Naegleria, Plasmodium, Trichomonas, and Trypanosoma and their medical and taxonomical consequences. Proc. Natl. Acad. Sci. USA 1990, 87, 2414–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, H.J. The relation of recombination to mutational advance. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 1964, 1, 2–9. [Google Scholar] [CrossRef]
- Haigh, J. The accumulation of deleterious genes in a population—Muller’s Ratchet. Theor. Popul. Biol. 1978, 14, 251–267. [Google Scholar] [CrossRef]
- Tibayrenc, M. Clonality in Leishmania. Parasitol. Today 1993, 9, 58. [Google Scholar] [CrossRef]
- Prugnolle, F.; De Meeûs, T. The impact of clonality on parasite population genetic structure. Parasite 2008, 15, 455–457. [Google Scholar] [CrossRef] [Green Version]
- Tibayrenc, M.; Ayala, F.J. Trypanosoma cruzi populations: More clonal than sexual. Parasitol. Today 1987, 3, 189–190. [Google Scholar] [CrossRef] [Green Version]
- Tait, A.; Barry, J.D.; Wink, R.; Sanderson, A.; Crowe, J.S. Enzyme variation in T. brucei ssp. II. Evidence for T. b. rhodesiense being a set of variants of T. b. brucei. Parasitology 1985, 90, 89–100. [Google Scholar] [CrossRef]
- Tibayrenc, M.; Echalar, L.; Dujardin, J.P.; Poch, O.; Desjeux, P. The microdistribution of isoenzymic strains of Trypanosoma cruzi in southern Bolivia; new isoenzyme profiles and further arguments against Mendelian sexuality. Trans. R. Soc. Trop. Med. Hyg. 1984, 78, 519–525. [Google Scholar] [CrossRef]
- Hartl, D.L. Population genetics of microbial organisms. Curr. Opin. Genet. Dev. 1992, 2, 937–942. [Google Scholar] [CrossRef]
- Schulte, R.D.; Makus, C.; Schulenburg, H. Host-parasite coevolution favours parasite genetic diversity and horizontal gene transfer. J. Evol. Biol. 2013, 26, 1836–1840. [Google Scholar] [CrossRef]
- Lively, C.M.; Morran, L.T. The ecology of sexual reproduction. J. Evol. Biol. 2014, 27, 1292–1303. [Google Scholar] [CrossRef] [Green Version]
- Cavalier-Smith, T. Origins of the machinery of recombination and sex. Heredity 2002, 88, 125–141. [Google Scholar] [CrossRef]
- Speijer, D.; Lukeš, J.; Eliáš, M. Sex is a ubiquitous, ancient, and inherent attribute of eukaryotic life. Proc. Natl. Acad. Sci. USA 2015, 112, 8827–8834. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, H.; Bernstein, C. Evolutionary origin of recombination during meiosis. Bioscience 2010, 60, 498–505. [Google Scholar] [CrossRef]
- Heitman, J. Sexual reproduction and the evolution of microbial pathogens. Curr. Biol. 2006, 16, R711–R725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heitman, J. Evolution of eukaryotic microbial pathogens via covert sexual reproduction. Cell Host Microbe 2010, 8, 86–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feretzaki, M.; Heitman, J. Unisexual reproduction drives evolution of eukaryotic microbial pathogens. PLoS Pathog. 2013, 9, e1003674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, W. The sexual side of parasitic protists. Mol. Biochem. Parasitol. 2021, 243, 111371. [Google Scholar] [CrossRef] [PubMed]
- Sterkers, Y.; Crobu, L.; Lachaud, L.; Pagès, M.; Bastien, P. Parasexuality and mosaic aneuploidy in Leishmania: Alternative genetics. Trends Parasitol. 2014, 30, 429–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rougeron, V.; de Meeûs, T.; Bañuls, A.L. Reproduction in Leishmania: A focus on genetic exchange. Infect. Genet. Evol. 2017, 50, 128–132. [Google Scholar] [CrossRef]
- Rougeron, V.; de Meeûs, T.; Bañuls, A.L. A primer for Leishmania population genetic studies. Trends Parasitol. 2015, 31, 52–59. [Google Scholar] [CrossRef]
- Berman, J. Ploidy plasticity: A rapid and reversible strategy for adaptation to stress. FEMS Yeast Res. 2016, 16, fow020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forche, A.; Alby, K.; Schaefer, D.; Johnson, A.D.; Berman, J.; Bennett, R.J. The parasexual cycle in Candida albicans provides an alternative pathway to meiosis for the formation of recombinant strains. PLoS Biol. 2008, 6, e110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gladyshev, E.A.; Arkhipova, I.R. Genome structure of bdelloid rotifers: Shaped by asexuality or desiccation? J. Hered. 2010, 101 (Suppl 1), S85–S93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maciver, S.K.; Koutsogiannis, Z.; De Obeso Fernández Del Valle, A. “Meiotic genes” are constitutively expressed in an asexual amoeba and are not necessarily involved in sexual reproduction. Biol. Lett. 2019, 15, 20180871. [Google Scholar] [CrossRef] [Green Version]
- Tait, A. Evidence for diploidy and mating in trypanosomes. Nature 1980, 287, 536–538. [Google Scholar] [CrossRef]
- Tibayrenc, M.; Ayala, F.J. How clonal are Trypanosoma and Leishmania? Trends Parasitol. 2013, 29, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Dujardin, J.C.; Bañuls, A.L.; Llanos-Cuentas, A.; Alvarez, E.; DeDoncker, S.; Jacquet, D.; Le Ray, D.; Arevalo, J.; Tibayrenc, M. Putative Leishmania hybrids in the Eastern Andean valley of Huanuco, Peru. Acta Trop. 1995, 59, 293–307. [Google Scholar] [CrossRef]
- Tibayrenc, M.; Ayala, F.J. Reproductive clonality of pathogens: A perspective on pathogenic viruses, bacteria, fungi, and parasitic protozoa. Proc. Natl. Acad. Sci. USA 2012, 109, E3305–E3313. [Google Scholar] [CrossRef] [Green Version]
- Tibayrenc, M.; Ayala, F.J. Is predominant clonal evolution a common evolutionary adaptation to parasitism in pathogenic parasitic protozoa, fungi, bacteria, and viruses? Adv. Parasitol. 2017, 97, 243–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Meeûs, T.; Balloux, F. Clonal reproduction and linkage disequilibrium in diploids: A simulation study. Infect. Genet. Evol. 2004, 4, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Prugnolle, F.; de Meeûs, T. Apparent high recombination rates in clonal parasitic organisms due to inappropriate sampling design. Heredity 2010, 104, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, J.D.; Llewellyn, M.S. Reproductive clonality in protozoan pathogens—Truth or artefact? Mol. Ecol. 2014, 23, 4195–4202. [Google Scholar] [CrossRef]
- Rougeron, V.; de Meeûs, T.; Ouraga, S.K.; Hide, M.; Bañuls, A.L. “Everything you always wanted to know about sex (but were afraid to ask)” in Leishmania after two decades of laboratory and field analyses. PLoS Pathog. 2010, 6, 7–8. [Google Scholar] [CrossRef]
- Shaik, J.S.; Dobson, D.E.; Sacks, D.L.; Beverley, S.M. Leishmania sexual reproductive strategies as resolved through computational methods designed for aneuploid genomes. Genes 2021, 12, 167. [Google Scholar] [CrossRef]
- Tait, A. Sexual processes in the Kinetoplastida. Parasitology 1983, 86, 29–57. [Google Scholar] [CrossRef]
- Gibson, W.C.; Osinga, K.A.; Michels, P.A.M.; Borst, P. Trypanosomes of subgenus trypanozoon are diploid for housekeeping genes. Mol. Biochem. Parasitol. 1985, 16, 231–242. [Google Scholar] [CrossRef]
- Jenni, L.; Marti, S.; Schweizer, J.; Betschart, B.; Le Page, R.W.F.; Wells, J.M.; Tait, A.; Paindavoine, P.; Pays, E.; Steinert, M. Hybrid formation between African trypanosomes during cyclical transmission. Nature 1986, 322, 173–175. [Google Scholar] [CrossRef]
- Gibson, W.; Whittington, H. Genetic exchange in Trypanosoma brucei: Selection of hybrid trypanosomes by introduction of genes conferring drug resistance. Mol. Biochem. Parasitol. 1993, 60, 19–26. [Google Scholar] [CrossRef]
- Gibson, W.; Garside, L.; Bailey, M. Trisomy and chromosome size changes in hybrid trypanosomes from a genetic cross between Trypanosoma brucei rhodesiense and T. b. brucei. Mol. Biochem. Parasitol. 1992, 51, 189–199. [Google Scholar] [CrossRef]
- Gibson, W. Molecular ecology of infectious diseases. Trends Microbiol. 1995, 3, 1–2. [Google Scholar] [CrossRef]
- Gibson, W.; Stevens, J. Genetic exchange in the Trypanosomatidae. Adv. Parasitol. 1999, 43, 1–46. [Google Scholar] [PubMed]
- MacLeod, A.; Tweedie, A.; McLellan, S.; Taylor, S.; Cooper, A.; Sweeney, L.; Turner, C.M.R.; Tait, A. Allelic segregation and independent assortment in T. brucei crosses: Proof that the genetic system is Mendelian and involves meiosis. Mol. Biochem. Parasitol. 2005, 143, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Wirtz, E.; Clayton, C. Inducible gene expression in trypanosomes mediated by a prokaryotic repressor. Science 1995, 268, 1179–1183. [Google Scholar] [CrossRef]
- Bingle, L.E.H.; Eastlake, J.L.; Bailey, M.; Gibson, W.C. A novel GFP approach for the analysis of genetic exchange in trypanosomes allowing the in situ detection of mating events. Microbiology 2001, 147, 3231–3240. [Google Scholar] [CrossRef] [Green Version]
- Gibson, W.; Peacock, L.; Ferris, V.; Williams, K.; Bailey, M. The use of yellow fluorescent hybrids to indicate mating in Trypanosoma brucei. Parasites Vectors 2008, 1, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peacock, L.; Ferris, V.; Bailey, M.; Gibson, W. Intraclonal mating occurs during tsetse transmission of Trypanosoma brucei. Parasites Vectors 2009, 2, 43. [Google Scholar] [CrossRef] [Green Version]
- Malik, S.-B.; Pightling, A.W.; Stefaniak, L.M.; Schurko, A.M.; Logsdon, J.M. An expanded inventory of conserved meiotic genes provides evidence for sex in Trichomonas vaginalis. PLoS ONE 2008, 3, e2879. [Google Scholar] [CrossRef]
- Burki, F.; Roger, A.J.; Brown, M.W.; Simpson, A. The new Tree of Eukaryotes. Trends Ecol. Evol. 2020, 35, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Peacock, L.; Ferris, V.; Sharma, R.; Sunter, J.; Bailey, M.; Carrington, M.; Gibson, W. Identification of the meiotic life cycle stage of Trypanosoma brucei in the tsetse fly. Proc. Natl. Acad. Sci. USA 2011, 108, 3671–3676. [Google Scholar] [CrossRef] [Green Version]
- Minia, I.; Merce, C.; Terrao, M.; Clayton, C. Translation regulation and RNA granule formation after heat shock of procyclic form Trypanosoma brucei: Many heat-induced mRNAs are also increased during differentiation to mammalian-infective forms. PLoS Negl. Trop. Dis. 2016, 10, e0004982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peacock, L.; Bailey, M.; Carrington, M.; Gibson, W. Meiosis and haploid gametes in the pathogen Trypanosoma brucei. Curr. Biol. 2014, 24, 181–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peacock, L.; Ferris, V.; Bailey, M.; Gibson, W. Mating compatibility in the parasitic protist Trypanosoma brucei. Parasites Vectors 2014, 7, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peacock, L.; Kay, C.; Farren, C.; Bailey, M.; Carrington, M.; Gibson, W. Sequential production of gametes during meiosis in trypanosomes. Commun. Biol. 2021, 4, 555. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.S.F.; Salazar-Sánchez, R.; Castillo-Neyra, R.; Borrini-Mayorí, K.; Chipana-Ramos, C.; Vargas-Maquera, M.; Ancca-Juarez, J.; Náquira-Velarde, C.; Levy, M.Z.; Brisson, D. Immigration and establishment of Trypanosoma cruzi in Arequipa, Peru. PLoS ONE 2019, 14, e0221678. [Google Scholar] [CrossRef]
- Messenger, L.A.; Miles, M.A. Evidence and importance of genetic exchange among field populations of Trypanosoma cruzi. Acta Trop. 2015, 151, 150–155. [Google Scholar] [CrossRef]
- Yeo, M.; Mauricio, I.L.; Messenger, L.A.; Lewis, M.D.; Llewellyn, M.S.; Acosta, N.; Bhattacharyya, T.; Diosque, P.; Carrasco, H.J.; Miles, M.A. Multilocus sequence typing (MLST) for lineage assignment and high resolution diversity studies in Trypanosoma cruzi. PLoS Negl. Trop. Dis. 2011, 5, e1049. [Google Scholar] [CrossRef]
- Ramírez, J.C.; Torres, C.; Curto, M.d.l.A.; Schijman, A.G. New insights into Trypanosoma cruzi evolution, genotyping and molecular diagnostics from satellite DNA sequence analysis. PLoS Negl. Trop. Dis. 2017, 11, e0006139. [Google Scholar] [CrossRef] [Green Version]
- Schwabl, P.; Imamura, H.; Van den Broeck, F.; Costales, J.A.; Maiguashca-Sánchez, J.; Miles, M.A.; Andersson, B.; Grijalva, M.J.; Llewellyn, M.S. Meiotic sex in Chagas disease parasite Trypanosoma cruzi. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Westenberger, S.J.; Barnabé, C.; Campbell, D.A.; Sturm, N.R. Two hybridization events define the population structure of Trypanosoma cruzi. Genetics 2005, 171, 527–543. [Google Scholar] [CrossRef] [Green Version]
- De Freitas, J.M.; Augusto-Pinto, L.; Pimenta, J.R.; Bastos-Rodrigues, L.; Gonçalves, V.F.; Teixeira, S.M.R.; Chiari, E.; Junqueira, Â.C.V.; Fernandes, O.; Macedo, A.M.; et al. Ancestral genomes, sex, and the population structure of Trypanosoma cruzi. PLoS Pathog. 2006, 2, e24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasini, N.; Diosque, P. Evolution of Trypanosoma cruzi: Clarifying hybridisations, mitochondrial introgressions and phylogenetic relationships between major lineages. Memórias Inst. Oswaldo Cruz 2015, 110, 403–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaunt, M.W.; Yeo, M.; Frame, J.A.; Stothard, J.R.; Carrasco, H.J.; Taylor, M.C.; Mena, S.S.; Veazey, P.; Miles, G.A.J.; Acosta, N.; et al. Mechanism of genetic exchange in American trypanosomes. Nature 2003, 421, 936–939. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.D.; Llewellyn, M.S.; Gaunt, M.W.; Yeo, M.; Carrasco, H.J.; Miles, M.A. Flow cytometric analysis and microsatellite genotyping reveal extensive DNA content variation in Trypanosoma cruzi populations and expose contrasts between natural and experimental hybrids. Int. J. Parasitol. 2009, 39, 1305–1317. [Google Scholar] [CrossRef] [Green Version]
- Iovannisci, D.M.; Beverley, S.M. Structural alterations of chromosome 2 in Leishmania major as evidence for diploidy, including spontaneous amplification of the mini-exon array. Mol. Biochem. Parasitol. 1989, 34, 177–188. [Google Scholar] [CrossRef]
- Beck, J.T.; Ullman, B. Biopterin conversion to reduced folates by Leishmania donovani promastigotes. Mol. Biochem. Parasitol. 1991, 49, 21–28. [Google Scholar] [CrossRef]
- Cruz, A.K.; Titus, R.; Beverley, S.M. Plasticity in chromosome number and testing of essential genes in Leishmania by targeting. Proc. Natl. Acad. Sci. USA 1993, 90, 1599–1603. [Google Scholar] [CrossRef] [Green Version]
- Reguera, R.M.; Redondo, C.M.; Pérez-Pertejo, Y.; Balaña-Fouce, R. S-Adenosylmethionine in protozoan parasites: Functions, synthesis and regulation. Mol. Biochem. Parasitol. 2007, 152, 1–10. [Google Scholar] [CrossRef]
- Gueiros-Filho, F.J.; Beverley, S.M. Selection against the dihydrofolate reductase-thymidylate synthase (DHFR-TS) locus as a probe of genetic alterations in Leishmania major. Mol. Cell. Biol. 1996, 16, 5655–5663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachaud, L.; Bourgeois, N.; Kuk, N.; Morelle, C.; Crobu, L.; Merlin, G.; Bastien, P.; Pagès, M.; Sterkers, Y. Constitutive mosaic aneuploidy is a unique genetic feature widespread in the Leishmania genus. Microbes Infect. 2014, 16, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Kelly, J.M.; Law, J.M.; Chapman, C.J.; Van, V.E.; Evans, D.A. Evidence of genetic recombination in Leishmania. Mol. Biochem. Parasitol. 1991, 46, 253–263. [Google Scholar] [CrossRef]
- Belli, A.A.; Miles, M.A.; Kelly, J.M. A putative Leishmania panamensis/Leishmania braziliensis hybrid is a causative agent of human cutaneous leishmaniasis in Nicaragua. Parasitology 1994, 109, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Nolder, D.; Roncal, N.; Davies, C.R.; Llanos-Cuentas, A.; Miles, M.A. Multiple hybrid genotypes of Leishmania (Viannia) in a focus of mucocutaneous leishmaniasis. Am. J. Trop. Med. Hyg. 2007, 76, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Ravel, C.; Cortes, S.; Pratlong, F.; Morio, F.; Dedet, J.P.; Campino, L. First report of genetic hybrids between two very divergent Leishmania species: Leishmania infantum and Leishmania major. Int. J. Parasitol. 2006, 36, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Volf, P.; Benkova, I.; Myskova, J.; Sadlova, J.; Campino, L.; Ravel, C. Increased transmission potential of Leishmania major/Leishmania infantum hybrids. Int. J. Parasitol. 2007, 37, 589–593. [Google Scholar] [CrossRef] [Green Version]
- Rogers, M.B.; Downing, T.; Smith, B.A.; Imamura, H.; Sanders, M.; Svobodova, M.; Volf, P.; Berriman, M.; Cotton, J.A.; Smith, D.F. Genomic confirmation of hybridisation and recent inbreeding in a vector-isolated Leishmania population. PLoS Genet. 2014, 10, e1004092. [Google Scholar] [CrossRef] [Green Version]
- Akopyants, N.S.; Kimblin, N.; Secundino, N.; Patrick, R.; Peters, N.; Lawyer, P.; Dobson, D.E.; Beverley, S.M.; Sacks, D.L. Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector. Science 2009, 324, 265–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inbar, E.; Akopyants, N.S.; Charmoy, M.; Romano, A.; Lawyer, P.; Elnaiem, D.E.A.; Kauffmann, F.; Barhoumi, M.; Grigg, M.; Owens, K.; et al. The mating competence of geographically diverse Leishmania major strains in their natural and unnatural sand fly vectors. PLoS Genet. 2013, 9, e1003672. [Google Scholar] [CrossRef]
- Romano, A.; Inbar, E.; Debrabant, A.; Charmoy, M.; Lawyer, P.; Ribeiro-Gomes, F.; Barhoumi, M.; Grigg, M.; Shaik, J.; Dobson, D.; et al. Cross-species genetic exchange between visceral and cutaneous strains of Leishmania in the sand fly vector. Proc. Natl. Acad. Sci. USA 2014, 111, 16808–16813. [Google Scholar] [CrossRef] [Green Version]
- Sadlova, J.; Yeo, M.; Seblova, V.; Lewis, M.D.; Mauricio, I.; Volf, P.; Miles, M.A. Visualisation of Leishmania donovani fluorescent hybrids during early stage development in the sand fly vector. PLoS ONE 2011, 6, e19851. [Google Scholar] [CrossRef]
- Calvo-Álvarez, E.; Álvarez-Velilla, R.; Jiménez, M.; Molina, R.; Pérez-Pertejo, Y.; Balaña-Fouce, R.; Reguera, R.M. First evidence of intraclonal genetic exchange in trypanosomatids using two Leishmania infantum fluorescent transgenic clones. PLoS Negl. Trop. Dis. 2014, 8, e3075. [Google Scholar] [CrossRef]
- Inbar, E.; Shaik, J.; Iantorno, S.A.; Romano, A.; Nzelu, C.O.; Owens, K.; Sanders, M.J.; Dobson, D.; Cotton, J.A.; Grigg, M.E.; et al. Whole genome sequencing of experimental hybrids supports meiosis-like sexual recombination in Leishmania. PLoS Genet. 2019, 15, e1008042. [Google Scholar] [CrossRef] [Green Version]
- MacLeod, A.; Tweedie, A.; McLellan, S.; Taylor, S.; Hall, N.; Berriman, M.; El-Sayed, N.M.; Hope, M.; Turner, C.M.R.; Tait, A. The genetic map and comparative analysis with the physical map of Trypanosoma brucei. Nucleic Acids Res. 2005, 33, 6688–6693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louradour, I.; Ferreira, T.R.; Ghosh, K.; Shaik, J.; Sacks, D. In vitro generation of Leishmania hybrids. Cell Rep. 2020, 31, 107507. [Google Scholar] [CrossRef] [PubMed]
- Telittchenko, R.; Descoteaux, A. Study on the occurrence of genetic exchange among parasites of the Leishmania mexicana complex. Front. Cell. Infect. Microbiol. 2020, 10, 607253. [Google Scholar] [CrossRef]
- Jensen, R.E.; Englund, P.T. Network news: The replication of kinetoplast DNA. Annu. Rev. Microbiol. 2012, 66, 473–491. [Google Scholar] [CrossRef] [PubMed]
- Stuart, K.; Panigrahi, A.K. RNA editing: Complexity and complications. Mol. Microbiol. 2002, 45, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.H.; Lai, D.H.; Zheng, L.L.; Wu, J.; Lukeš, J.; Hide, G.; Lun, Z.R. Analysis of the mitochondrial maxicircle of Trypanosoma lewisi, a neglected human pathogen. Parasites Vectors 2015, 8, 665. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Liu, Y.; Motyka, S.A.; Agbo, E.E.C.; Englund, P.T. Fellowship of the rings: The replication of kinetoplast DNA. Trends Parasitol. 2005, 21, 363–369. [Google Scholar] [CrossRef]
- Sternberg, J.; Tait, A.; Haley, S.; Wells, J.M.; Le Page, R.W.F.; Schweizer, J.; Jenni, L. Gene exchange in African trypanosomes: Characterisation of a new hybrid genotype. Mol. Biochem. Parasitol. 1988, 27, 191–200. [Google Scholar] [CrossRef]
- Sternberg, J.; Turner, C.M.R.; Wells, J.M.; Ranford-Cartwright, L.C.; Le Page, R.W.F.; Tait, A. Gene exchange in African trypanosomes: Frequency and allelic segregation. Mol. Biochem. Parasitol. 1989, 34, 269–279. [Google Scholar] [CrossRef]
- Gibson, W.C. Analysis of a genetic cross between Trypanosoma brucei rhodesiense and T. b. brucei. Parasitology 1989, 99, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Michael, C.; Turner, R.; Hide, G.; Buchanan, N.; Tait, A. Trypanosoma brucei: Inheritance of kinetoplast DNA maxicircles in a genetic cross and their segregation during vegetative growth. Exp. Parasitol. 1995, 80, 234–241. [Google Scholar]
- Gibson, W.; Garside, L. Kinetoplast DNA minicircles are inherited from both parents in genetic hybrids of Trypanosoma brucei. Mol. Biochem. Parasitol. 1990, 42, 45–53. [Google Scholar] [CrossRef]
- Birky, C.W. Relaxed cellular controls and organelle heredity. Science 1983, 222, 468–475. [Google Scholar] [CrossRef]
- Gibson, W.; Crow, M.; Kearns, J. Kinetoplast DNA minicircles are inherited from both parents in genetic crosses of Trypanosoma brucei. Parasitol. Res. 1997, 83, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.L.; Repolês, B.M.; da Silva, M.S.; Mendes, I.C.; Marin, P.A.; Aguiar, P.H.N.; Santos, S.d.S.; Franco, G.R.; Macedo, A.M.; Pena, S.D.J.; et al. The recombinase Rad51 plays a key role in events of genetic exchange in Trypanosoma cruzi. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Rusman, F.; Tomasini, N.; Yapur, N.-F.; Puebla, A.F.; Ragone, P.G.; Diosque, P. Elucidating diversity in the class composition of the minicircle hypervariable region of Trypanosoma cruzi: New perspectives on typing and kDNA inheritance. PLoS Negl. Trop. Dis. 2019, 13, e0007536. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Corbo, C.; Domínguez-Asenjo, B.; Martínez-Valladares, M.; Pérez-Pertejo, Y.; García-Estrada, C.; Balaña-Fouce, R.; Reguera, R.M. Reproduction in Trypanosomatids: Past and Present. Biology 2021, 10, 471. https://doi.org/10.3390/biology10060471
Gutiérrez-Corbo C, Domínguez-Asenjo B, Martínez-Valladares M, Pérez-Pertejo Y, García-Estrada C, Balaña-Fouce R, Reguera RM. Reproduction in Trypanosomatids: Past and Present. Biology. 2021; 10(6):471. https://doi.org/10.3390/biology10060471
Chicago/Turabian StyleGutiérrez-Corbo, Camino, Bárbara Domínguez-Asenjo, María Martínez-Valladares, Yolanda Pérez-Pertejo, Carlos García-Estrada, Rafael Balaña-Fouce, and Rosa M. Reguera. 2021. "Reproduction in Trypanosomatids: Past and Present" Biology 10, no. 6: 471. https://doi.org/10.3390/biology10060471
APA StyleGutiérrez-Corbo, C., Domínguez-Asenjo, B., Martínez-Valladares, M., Pérez-Pertejo, Y., García-Estrada, C., Balaña-Fouce, R., & Reguera, R. M. (2021). Reproduction in Trypanosomatids: Past and Present. Biology, 10(6), 471. https://doi.org/10.3390/biology10060471