
The Role of Microbiota in the Pathogenesis of Esophageal Adenocarcinoma


Abstract
:Simple Summary
Abstract
1. Introduction
2. Microbiota Expression: Normal vs. Pathologic
2.1. Mutualistic and Dysbiotic Roles of Microbiota
2.2. Microbiota Composition in Healthy Esophageal Tissue

{kind=link}
| Article | Publication Year | Method | Sample Size (n) | Number of Identified Genera | Dominant Phyla | Dominant Genera |
|---|---|---|---|---|---|---|
| Peter et al. [42] | 2020 | 16S rRNA gene DNA sequencing | 12 | N/A | Firmicutes (47.81%), Proteobacteria (20.67%), Bacteroidetes (16.93%), Actinobacteria (5.57%), Fusobacteria (4.76%) | Tissierella soehngenia (16.67%) Lactobacillus (7.15%) Streptococcus (7.27%) Acinetobacter (5.80%) Prevotella (5.24%) |
| Dong et al. [43] | 2018 | 16S rRNA Illumina sequencing | 27 | 594 | Firmicutes (37.42%) Proteobacteria (43.61%) Bacteroidetes (13.17%) Actinobacteria (2.53%) Fusobacteria (1.22%) TM7 (1.06%) | Streptococcus Neisseria Prevotella Actinobacillus Veillonella |
| Blackett et al. [45] | 2013 | Selective media cultures + 16S rRNA PCR | 39 | 19 | N/A | Streptococcus Prevotella Staphylococcus Rothia Actinomyces Bifidobacterium Staphylococcus Neisseria |
| Yang et al. [46] | 2009 | 16S rRNA Sanger Sequencing | 12 | N/A | Firmicutes | Streptococcus (78.750%) Prevotella (4.300%) Gemella (3.400%) Veilonella (3.075%) Pasteurellaceae (2.075%) Haemophilus (1.925%) Rothia (1.025%) |
| Zilberstein et al. [40] | 2007 | Selective media cultures | 10 | N/A | N/A | Streptococcus (40%) Staphylococcus (20%) Corynebacterium (10%) Lactobacillus (10%) Peptococcus (10%) |
| Pei et al. [37] | 2004 | Broad-range 16S rDNA PCR | 4 | 41 | Firmicutes Bacteroidetes Actinobacteria Fusobacteria TM7 | Streptococcus (39%) Prevotella (17%) Veilonella (14%) |
2.3. Microbiota Composition in Pathological Esophageal Tissue
| Article | Publication Year | Method | Sample Size | Findings | |||
|---|---|---|---|---|---|---|---|
| Control | GERD | BE | EAC | ||||
| Peter et al. [42] | 2020 | 16S rRNA | 12 | N/A | 31 | 10 |
|
| Zhou et al. [50] | 2020 | 16S rRNA | 16 | N/A | 17 | 6 |
|
| Lopetuso et al. [48] | 2020 | 16S rRNA | 10 | N/A | 10 | 6 |
|
| Snider et al. [49] | 2019 | 16S rRNA | 16 | N/A | 25 | 4 |
|
| Deshpande et al. [9] | 2018 | 16S rRNA 18S rRNA | 59 | 29 | 5 | 1 |
|
| Elliott et al. [47] | 2017 | 16S rRNA | 16 | N/A | 17 | 15 |
|
| Gall et al. [51] | 2015 | 16S rRNA | N/A | N/A | 12 | N/A |
|
| Amir et al. [52] | 2014 | 16S rRNA | 15 | N/A | 6 | N/A |
|
| Yang et al. [46] | 2009 | 16S rRNA | 12 | N/A | 10 | N/A |
|
| Macfarlane et al. [53] | 2007 | 16S rDNA | 7 | N/A | 7 | N/A |
|
2.4. Controversial Associations between Helicobacter pylori and the EAC Cascade
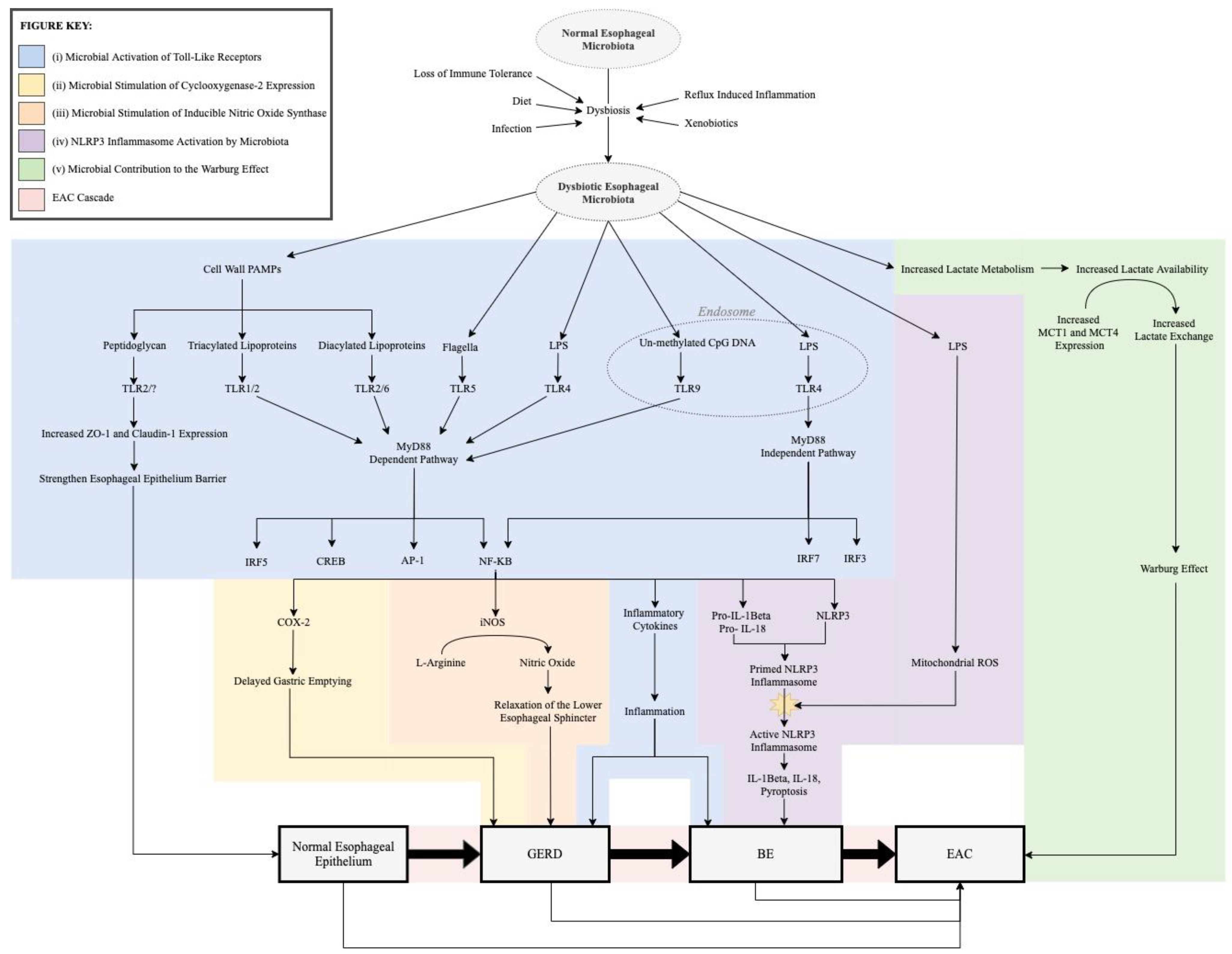
3. Role of Microbiota in the Pathogenesis of the EAC Cascade
3.1. Microbial Activation of Toll-Like Receptors
3.1.1. Background
3.1.2. TLR1, TLR2, and TLR6
3.1.3. TLR 4
3.1.4. TLR 5
3.1.5. TLR 9
3.1.6. Summary of TLRs in the EAC Cascade
3.2. Microbial Stimulation of Cyclooxygenase-2 Expression
3.3. Microbial Stimulation of Inducible Nitric Oxide Synthase
3.4. NLRP3 Inflammasome Activation by Microbiota
3.5. Microbial Contribution to the Warburg Effect
4. Therapeutic Potential of Esophageal Microbiota
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Arnold, M.; Laversanne, M.; Brown, L.M.; Devesa, S.S.; Bray, F. Predicting the Future Burden of Esophageal Cancer by Histological Subtype: International Trends in Incidence up to 2030. Am. J. Gastroenterol. 2017, 112, 1247–1255. [Google Scholar] [CrossRef]
- Coleman, H.G.; Xie, S.-H.; Lagergren, J. The Epidemiology of Esophageal Adenocarcinoma. Gastroenterology 2018, 154, 390–405. [Google Scholar] [CrossRef]
- Pohl, H.; Welch, H.G. The Role of Overdiagnosis and Reclassification in the Marked Increase of Esophageal Adenocarcinoma Incidence. J. Natl. Cancer Inst. 2005, 97, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA A Cancer J. Clin. 2020, 70, 145–164. [Google Scholar] [CrossRef] [Green Version]
- SEER*Explorer: An Interactive Website for SEER Cancer Statistics. Surveillance Research Program, National Cancer Institute. 2020. Available online: https://seer.cancer.gov/explorer/ (accessed on 1 November 2020).
- Zhang, Y. Epidemiology of esophageal cancer. World J. Gastroenterol. 2013, 19, 5598–5606. [Google Scholar] [CrossRef]
- Giroux, V.; Rustgi, A.K. Metaplasia: Tissue injury adaptation and a precursor to the dysplasia–cancer sequence. Nat. Rev. Cancer 2017, 17, 594–604. [Google Scholar] [CrossRef]
- Deshpande, N.P.; Riordan, S.M.; Castaño-Rodríguez, N.; Wilkins, M.R.; Kaakoush, N.O. Signatures within the esophageal microbiome are associated with host genetics, age, and disease. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zachariah, R.A.; Goo, T.; Lee, R.H. Mechanism and Pathophysiology of Gastroesophageal Reflux Disease. Gastrointest. Endosc. Clin. N. Am. 2020, 30, 209–226. [Google Scholar] [CrossRef]
- De Giorgi, F.; Palmiero, M.; Esposito, I.; Mosca, F.; Cuomo, R. Pathophysiology of gastro-oesophageal reflux disease. Acta Otorhinolaryngol. Ital. 2006, 26, 241–246. Available online: https://pubmed.ncbi.nlm.nih.gov/17345925 (accessed on 12 June 2021). [PubMed]
- Sarosiek, J.; Feng, T.; McCallum, R.W. The Interrelationship Between Salivary Epidermal Growth Factor and the Functional Integrity of the Esophageal Mucosal Barrier in the Rat. Am. J. Med. Sci. 1991, 302, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Helm, J.F.; Dodds, W.J.; Riedel, D.R.; Teeter, B.C.; Hogan, W.J.; Arndorfer, R.C. Determinants of esophageal acid clearance in normal subjects. Gastroenterology 1983, 85, 607–612. [Google Scholar] [CrossRef]
- Takashima, T.; Tsukano, M.; Kawano, H. Digestive Endocrine Cell Numbers Contribute to Contraction/Relaxation of the Lower Esophageal Sphincter. SAGE Open Nurs. 2016, 2. [Google Scholar] [CrossRef]
- Parikh, A.; Thevenin, C. Physiology, Gastrointestinal Hormonal Control; StatPearls Publishing: Treasure Island, FL, USA, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK537284/?report=classic (accessed on 13 June 2021).
- Ceranowicz, P.; Warzecha, Z.; Dembinski, A. Peptidyl hormones of endocrine cells origin in the gut-their discovery and physiological relevance. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2015, 66, 11–27. [Google Scholar]
- Stefanidis, D.; The SAGES Guidelines Committee; Hope, W.W.; Kohn, G.; Reardon, P.R.; Richardson, W.S.; Fanelli, R.D. Guidelines for surgical treatment of gastroesophageal reflux disease. Surg. Endosc. 2010, 24, 2647–2669. [Google Scholar] [CrossRef]
- Woodland, P.; Sifrim, D. The refluxate: The impact of its magnitude, composition and distribution. Best Pract. Res. Clin. Gastroenterol. 2010, 24, 861–871. [Google Scholar] [CrossRef]
- Chen, C.-L.; Hsu, P.-I. Current Advances in the Diagnosis and Treatment of Nonerosive Reflux Disease. Gastroenterol. Res. Pract. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gokulan, R.C.; Garcia-Buitrago, M.T.; Zaika, A.I. From genetics to signaling pathways: Molecular pathogenesis of esophageal adenocarcinoma. Biochim. Biophys. Acta Bioenerg. 2019, 1872, 37–48. [Google Scholar] [CrossRef]
- Rubenstein, J.H.; Taylor, J.B. Meta-analysis: The association of oesophageal adenocarcinoma with symptoms of gastro-oesophageal reflux. Aliment. Pharmacol. Ther. 2010, 32, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, J.H.; Scheiman, J.M.; Sadeghi, S.; Whiteman, D.; Inadomi, J. Esophageal Adenocarcinoma Incidence in Individuals with Gastroesophageal Reflux: Synthesis and Estimates from Population Studies. Am. J. Gastroenterol. 2011, 106, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Haider, S.H.; Kwon, S.; Lam, R.; Lee, A.K.; Caraher, E.J.; Crowley, G.; Zhang, L.; Schwartz, T.M.; Zeig-Owens, R.; Liu, M.; et al. Predictive Biomarkers of Gastroesophageal Reflux Disease and Barrett’s Esophagus in World Trade Center Exposed Firefighters: A 15 Year Longitudinal Study. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hazelton, W.D.; Curtius, K.; Inadomi, J.; Vaughan, T.L.; Meza, R.; Rubenstein, J.H.; Hur, C.; Luebeck, E.G. The Role of Gastroesophageal Reflux and Other Factors during Progression to Esophageal Adenocarcinoma. Cancer Epidemiol. Biomark. Prev. 2015, 24, 1012–1023. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.J.; Ott, B.J.; Payne, W.S. The Incidence of Adenocarcinoma in Columnar-Lined (Barrett’s) Esophagus. N. Engl. J. Med. 1985, 313, 857–859. [Google Scholar] [CrossRef]
- Rubenstein, J.H.; Shaheen, N.J. Epidemiology, Diagnosis, and Management of Esophageal Adenocarcinoma. Gastroenterology 2015, 149, 302–317. [Google Scholar] [CrossRef] [Green Version]
- Levy, M.; Kolodziejczyk, A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Roy, S.; Trinchieri, S.R.G. Microbiota: A key orchestrator of cancer therapy. Nat. Rev. Cancer 2017, 17, 271–285. [Google Scholar] [CrossRef]
- Goodman, B.; Gardner, H. The microbiome and cancer. J. Pathol. 2018, 244, 667–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheflin, A.; Whitney, A.; Weir, T.L. Cancer-Promoting Effects of Microbial Dysbiosis. Curr. Oncol. Rep. 2014, 16, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Li, B.; Selmi, C.; Tang, R.; Gershwin, M.E.; Ma, X. The microbiome and autoimmunity: A paradigm from the gut–liver axis. Cell. Mol. Immunol. 2018, 15, 595–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017, 10, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Sarosiek, J.; McCallum, R.W. What Role Do Salivary Inorganic Components Play in Health and Disease of the Esophageal Mucosa? Digestion 1995, 56, 24–31. [Google Scholar] [CrossRef]
- Rourk, R.M.; Namiot, Z.; Sarosiek, J.; Yu, Z.; McCallum, R.W. Impairment of salivary epidermal growth factor secretory response to esophageal mechanical and chemical stimulation in patients with reflux esophagitis. Am. J. Gastroenterol. 1994, 89, 237–244. [Google Scholar]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Pei, Z.; Bini, E.J.; Yang, L.; Zhou, M.; Francois, F.; Blaser, M.J. Bacterial biota in the human distal esophagus. Proc. Natl. Acad. Sci. USA 2004, 101, 4250–4255. [Google Scholar] [CrossRef] [Green Version]
- Grusell, E.N.; Dahlén, G.; Ruth, M.; Ny, L.; Bergquist, H.; Bove, M.; Quiding-Järbrink, M. Bacterial flora of the human oral cavity, and the upper and lower esophagus. Dis. Esophagus 2012, 26, 84–90. [Google Scholar] [CrossRef]
- Pei, Z.; Yang, L.; Peek, R.M.; Levine, J.S.M.; Pride, D.T.; Blaser, M.J. Bacterial biota in reflux esophagitis and Barrett’s esophagus. World J. Gastroenterol. 2005, 11, 7277–7283. [Google Scholar] [CrossRef]
- Zilberstein, B.; Quintanilha, A.G.; Santos, M.A.A.; Pajecki, D.; de Moura, E.; Alves, P.R.A.; Filho, F.M.; De Souza, J.A.U.; Gama-Rodrigues, J. Digestive tract microbiota in healthy volunteers. Clin. Sci. 2007, 62, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parshina, S.N.; Stams, A.J.M. Soehngenia. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 1–3. [Google Scholar]
- Peter, S.; Pendergraft, A.; Vanderpol, W.; Wilcox, C.M.; Baig, K.R.K.K.; Morrow, C.; Izard, J.; Mannon, P.J. Mucosa-Associated Microbiota in Barrett’s Esophagus, Dysplasia, and Esophageal Adenocarcinoma Differ Similarly Compared with Healthy Controls. Clin. Transl. Gastroenterol. 2020, 11, e00199. [Google Scholar] [CrossRef]
- Dong, L.; Yin, J.; Zhao, J.; Ma, S.-R.; Wang, H.-R.; Wang, M.; Chen, W.; Wei, W.-Q. Microbial Similarity and Preference for Specific Sites in Healthy Oral Cavity and Esophagus. Front. Microbiol. 2018, 9, 1603. [Google Scholar] [CrossRef]
- Yin, J.; Dong, L.; Zhao, J.; Wang, H.; Li, J.; Yu, A.; Chen, W.; Wei, W. Composition and consistence of the bacterial microbiome in upper, middle and lower esophagus before and after Lugol’s iodine staining in the esophagus cancer screening. Scand. J. Gastroenterol. 2020, 55, 1467–1474. [Google Scholar] [CrossRef]
- Blackett, K.L.; Siddhi, S.S.; Cleary, S.; Steed, H.; Miller, M.H.; Macfarlane, S.; Macfarlane, G.T.; Dillon, J.F. Oesophageal bacterial biofilm changes in gastro-oesophageal reflux disease, Barrett’s and oesophageal carcinoma: Association or causality? Aliment. Pharmacol. Ther. 2013, 37, 1084–1092. [Google Scholar] [CrossRef]
- Yang, L.; Lu, X.; Nossa, C.W.; Francois, F.; Peek, R.M.; Pei, Z. Inflammation and Intestinal Metaplasia of the Distal Esophagus Are Associated with Alterations in the Microbiome. Gastroenterology 2009, 137, 588–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, D.R.F.; Walker, A.; O’Donovan, M.; Parkhill, J.; Fitzgerald, R.C. A non-endoscopic device to sample the oesophageal microbiota: A case-control study. Lancet Gastroenterol. Hepatol. 2017, 2, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Lopetuso, L.R.; Severgnini, M.; Pecere, S.; Ponziani, F.R.; Boskoski, I.; Larghi, A.; Quaranta, G.; Masucci, L.; Ianiro, G.; Camboni, T.; et al. Esophageal microbiome signature in patients with Barrett’s esophagus and esophageal adenocarcinoma. PLoS ONE 2020, 15, e0231789. [Google Scholar] [CrossRef]
- Snider, E.J.; Compres, G.; Freedberg, D.E.; Khiabanian, H.; Nobel, Y.R.; Stump, S.; Uhlemann, A.-C.; Lightdale, C.J.; Abrams, J.A. Alterations to the Esophageal Microbiome Associated with Progression from Barrett’s Esophagus to Esophageal Adenocarcinoma. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1687–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Shrestha, P.; Qiu, Z.; Harman, D.G.; Teoh, W.-C.; Al-Sohaily, S.; Liem, H.; Turner, I.; Ho, V. Distinct Microbiota Dysbiosis in Patients with Non-Erosive Reflux Disease and Esophageal Adenocarcinoma. J. Clin. Med. 2020, 9, 2162. [Google Scholar] [CrossRef]
- Gall, A.; Fero, J.; McCoy, C.; Claywell, B.C.; Sanchez, C.A.; Blount, P.L.; Li, X.; Vaughan, T.L.; Matsen, F.A.; Reid, B.J.; et al. Bacterial Composition of the Human Upper Gastrointestinal Tract Microbiome Is Dynamic and Associated with Genomic Instability in a Barrett’s Esophagus Cohort. PLoS ONE 2015, 10, e0129055. [Google Scholar] [CrossRef]
- Amir, I.; Konikoff, F.M.; Oppenheim, M.; Gophna, U.; Half, E.E. Gastric microbiota is altered in oesophagitis and Barrett’s oesophagus and further modified by proton pump inhibitors. Environ. Microbiol. 2014, 16, 2905–2914. [Google Scholar] [CrossRef]
- Macfarlane, S.; Furrie, E.; Macfarlane, G.T.; Dillon, J. Microbial Colonization of the Upper Gastrointestinal Tract in Patients with Barrett’s Esophagus. Clin. Infect. Dis. 2007, 45, 29–38. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Y.; Hu, J.; Wang, X.; Ren, M.; Lu, G.; Lu, X.; Zhang, D.; He, S. The Effect of Helicobacter pylori Eradication in Patients with Gastroesophageal Reflux Disease: A Meta-Analysis of Randomized Controlled Studies. Dig. Dis. 2020, 38, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Yaghoobi, M.; Farrokhyar, F.; Yuan, Y.; Hunt, R.H. Is There an Increased Risk of GERD After Helicobacter pylori Eradication?: A Meta-Analysis. Am. J. Gastroenterol. 2010, 105, 1007–1013. [Google Scholar] [CrossRef]
- Saad, A.M.; Choudhary, A.; Bechtold, M.L. Effect of Helicobacter pyloritreatment on gastroesophageal reflux disease (GERD): Meta-analysis of randomized controlled trials. Scand. J. Gastroenterol. 2012, 47, 129–135. [Google Scholar] [CrossRef]
- Erőss, B.; Farkas, N.; Vincze, A.; Tinusz, B.; Szapáry, L.; Garami, A.; Balaskó, M.; Sarlós, P.; Czopf, L.; Alizadeh, H.; et al. Helicobacter pyloriinfection reduces the risk of Barrett’s esophagus: A meta-analysis and systematic review. Helicobacter 2018, 23, e12504. [Google Scholar] [CrossRef] [Green Version]
- Sonnenberg, A.; Turner, K.O.; Genta, R.M. Interaction Between Ethnicity and Helicobacter pylori Infection in the Occurrence of Reflux Disease. J. Clin. Gastroenterol. 2019, 54, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Doorakkers, E.; Lagergren, J.; Santoni, G.; Engstrand, L.; Brusselaers, N. Helicobacter pylori eradication treatment and the risk of Barrett’s esophagus and esophageal adenocarcinoma. Helicobacter 2020, 25, e12688. [Google Scholar] [CrossRef] [Green Version]
- Nie, S.; Chen, T.; Yang, X.; Huai, P.; Lu, M. Association of Helicobacter pyloriinfection with esophageal adenocarcinoma and squamous cell carcinoma: A meta-analysis. Dis. Esophagus 2014, 27, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Zeglinas, C.; Artemaki, F.; Doulberis, M.; Kazakos, E.; Katsinelos, P.; Kountouras, J. Helicobacter pylori infection and esophageal adenocarcinoma: A review and a personal view. Ann. Gastroenterol. 2017, 31, 8–13. [Google Scholar] [CrossRef]
- Pasare, C.; Medzhitov, R. Toll-Like Receptors: Linking Innate and Adaptive Immunity. Adv. Exp. Med. Biol. 2005, 560, 11–18. [Google Scholar] [CrossRef]
- Schnare, M.; Barton, G.; Holt, A.C.; Takeda, K.; Akira, S.; Medzhitov, R. Toll-like receptors control activation of adaptive immune responses. Nat. Immunol. 2001, 2, 947–950. [Google Scholar] [CrossRef]
- El-Zayat, S.R.; Sibaii, H.; Mannaa, F.A. Toll-like receptors activation, signaling, and targeting: An overview. Bull. Natl. Res. Cent. 2019, 43, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Blander, J.M. Regulation of Phagosome Maturation by Signals from Toll-Like Receptors. Science 2004, 304, 1014–1018. [Google Scholar] [CrossRef]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. TLR signaling. Cell Death Differ. 2006, 13, 816–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dajon, M.; Iribarren, K.; Cremer, I. Toll-like receptor stimulation in cancer: A pro- and anti-tumor double-edged sword. Immunobiology 2017, 222, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Pradère, J.-P.; Dapito, D.H.; Schwabe, R.F. The Yin and Yang of Toll-like receptors in cancer. Oncogene 2014, 33, 3485–3495. [Google Scholar] [CrossRef] [Green Version]
- Uronis, J.M.; Mühlbauer, M.; Herfarth, H.H.; Rubinas, T.C.; Jones, G.; Jobin, C. Modulation of the Intestinal Microbiota Alters Colitis-Associated Colorectal Cancer Susceptibility. PLoS ONE 2009, 4, e6026. [Google Scholar] [CrossRef] [Green Version]
- Cen, X.; Liu, S.; Cheng, K. The Role of Toll-Like Receptor in Inflammation and Tumor Immunity. Front. Pharmacol. 2018, 9, 878. [Google Scholar] [CrossRef] [Green Version]
- Mulder, D.; Lobo, D.; Mak, N.; Justinich, C.J. Expression of Toll-Like Receptors 2 and 3 on Esophageal Epithelial Cell Lines and on Eosinophils During Esophagitis. Dig. Dis. Sci. 2011, 57, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Verbeek, R.E.; Siersema, P.D.; Vleggaar, F.P.; Kate, F.J.T.; Posthuma, G.; Souza, R.F.; de Haan, J.; Van Baal, J.W. Toll-like Receptor 2 Signalling and the Lysosomal Machinery in Barrett’s Esophagus. J. Gastrointest. Liver Dis. 2016, 25, 273–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huhta, H.; Helminen, O.; Lehenkari, P.P.; Saarnio, J.; Karttunen, T.J.; Kauppila, J.H. Toll-like receptors 1, 2, 4 and 6 in esophageal epithelium, Barrett’s esophagus, dysplasia and adenocarcinoma. Oncotarget 2016, 7, 23658–23667. [Google Scholar] [CrossRef]
- Zaidi, A.H.; Kelly, L.A.; Kreft, R.E.; Barlek, M.; Omstead, A.N.; Matsui, D.; Boyd, N.H.; Gazarik, K.E.; Heit, M.I.; Nistico, L.; et al. Associations of microbiota and toll-like receptor signaling pathway in esophageal adenocarcinoma. BMC Cancer 2016, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- De Oliviera Nascimento, L.; Massari, P.; Wetzler, L.M. The Role of TLR2 in Infection and Immunity. Front. Immunol. 2012, 3, 79. [Google Scholar] [CrossRef] [Green Version]
- Hörmann, N.; Brandão, I.; Jäckel, S.; Ens, N.; Lillich, M.; Walter, U.; Reinhardt, C. Gut Microbial Colonization Orchestrates TLR2 Expression, Signaling and Epithelial Proliferation in the Small Intestinal Mucosa. PLoS ONE 2014, 9, e113080. [Google Scholar] [CrossRef] [Green Version]
- Ruffner, M.A.; Song, L.; Maurer, K.; Shi, L.; Carroll, M.C.; Wang, J.X.; Muir, A.B.; Spergel, J.M.; Sullivan, K.E. Toll-like receptor 2 stimulation augments esophageal barrier integrity. Allergy 2019, 74, 2449–2460. [Google Scholar] [CrossRef] [PubMed]
- Kohtz, P.D.; Halpern, A.L.; Eldeiry, M.A.; Hazel, K.; Kalatardi, S.; Ao, L.; Meng, X.; Reece, T.B.; Fullerton, D.A.; Weyant, M.J. Toll-Like Receptor-4 Is a Mediator of Proliferation in Esophageal Adenocarcinoma. Ann. Thorac. Surg. 2019, 107, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Verbeek, R.E.; Siersema, P.D.; Kate, F.J.T.; Fluiter, K.; Souza, R.F.; Vleggaar, F.P.; Bus, P.; Van Baal, J.W.P.M. Toll-like receptor 4 activation in Barrett’s esophagus results in a strong increase in COX-2 expression. J. Gastroenterol. 2014, 49, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- Ang, Y. The role of interleukin-8 in cancer cells and microenvironment interaction. Front. Biosci. 2005, 10, 853–865. [Google Scholar] [CrossRef] [Green Version]
- Helminen, O.; Huhta, H.; Takala, H.; Lehenkari, P.P.; Saarnio, J.; Kauppila, J.H.; Karttunen, T.J. Increased Toll-like receptor 5 expression indicates esophageal columnar dysplasia. Virchows Archiv 2013, 464, 11–18. [Google Scholar] [CrossRef]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.; Akira, S.; Underhill, D.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nat. Cell Biol. 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Kauppila, J.H.; Takala, H.; Selander, K.S.; Lehenkari, P.P.; Saarnio, J.; Karttunen, T.J. Increased Toll-like receptor 9 expression indicates adverse prognosis in oesophageal adenocarcinoma. Histopathology 2011, 59, 643–649. [Google Scholar] [CrossRef]
- Bauer, S.; Kirschning, C.J.; Hacker, H.; Redecke, V.; Hausmann, S.; Akira, S.; Wagner, H.; Lipford, G.B. Human TLR9 confers responsiveness to bacterial DNA via species-specific CpG motif recognition. Proc. Natl. Acad. Sci. USA 2001, 98, 9237–9242. [Google Scholar] [CrossRef] [Green Version]
- Smyth, E.M.; Grosser, T.; Wang, M.; Yu, Y.; Fitz-Gerald, G.A. Prostanoids in health and disease. J. Lipid Res. 2009, 50, S423–S428. [Google Scholar] [CrossRef] [Green Version]
- Turman, M.V.; Marnett, L.J. Prostaglandin Endoperoxide Synthases. Compr. Nat. Prod. II 2010, 2010, 35–63. [Google Scholar] [CrossRef]
- Morris, C.D.; Armstrong, G.R.; Bigley, G.; Green, H.; Attwood, S.E. Cyclooxygenase-2 Expression in The Barrettʼs Metaplasia–Dysplasia–Adenocarcinoma Sequence. Am. J. Gastroenterol. 2001, 96, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.T. Cyclooxygenase-2 in oncogenesis. Clin. Chim. Acta 2011, 412, 671–687. [Google Scholar] [CrossRef] [PubMed]
- Greenhough, A.; Smartt, H.J.M.; Moore, A.E.; Roberts, H.R.; Williams, A.; Paraskevaand, C.; Kaidi, A. The COX-2/PGE2 pathway: Key roles in the hallmarks of cancer and adaptation to the tumour microenvironment. Carcinogenesis 2009, 30, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, K.T.; Fu, S.; Ramanujam, K.S.; Meltzer, S.J. Increased expression of inducible nitric oxide synthase and cyclooxygenase-2 in Barrett’s esophagus and associated adenocarcinomas. Cancer Res. 1998, 58, 2929–2934. [Google Scholar] [PubMed]
- Kandil, H.M.; Tanner, G.; Smalley, W.; Halter, S.; Radhika, A.; Dubois, R.N. Cyclooxygenase-2 expression in Barrett’s esophagus. Dig. Dis. Sci. 2001, 46, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Collares, E. Effect of bacterial lipopolysaccharide on gastric emptying of liquids in rats. Braz. J. Med. Biol. Res. 1997, 30, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Calatayud, S.; García-Zaragozá, E.; Hernández, C.; Quintana, E.; Felipo, V.; Esplugues, J.V.; Barrachina, M.D. Downregulation of nNOS and synthesis of PGs associated with endotoxin-induced delay in gastric emptying. Am. J. Physiol. Liver Physiol. 2002, 283, G1360–G1367. [Google Scholar] [CrossRef] [Green Version]
- Lipham, J.C.; Sandhu, K.K. Options to Address Delayed Gastric Emptying in Gastroesophageal Reflux Disease. Shackelford Surg. Aliment. Tract 2 Vol. Set 2019, 2019, 262–268. [Google Scholar] [CrossRef]
- Gourcerol, G.; Benanni, Y.; Boueyre, E.; Leroi, A.M.; Ducrotté, P. Influence of gastric emptying on gastro-esophageal reflux: A combined pH-impedance study. Neurogastroenterol. Motil. 2013, 25. [Google Scholar] [CrossRef]
- Emerenziani, S.; Sifrim, D. Gastroesophageal reflux and gastric emptying, revisited. Curr. Gastroenterol. Rep. 2005, 7, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Maddern, G.; Chatterton, B.E.; Collins, P.J.; Horowitz, M.; Shearman, D.J.C.; Jamieson, G.G. Solid and liquid gastric emptying in patients with gastro-oesophageal reflux. BJS 2005, 72, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, K.M.; Horowitz, M.; Riddell, P.S.; Maddern, G.; Myers, J.; Holloway, R.H.; Wishart, J.M.; Jamieson, G.G. Relations among autonomic nerve dysfunction, oesophageal motility, and gastric emptying in gastro-oesophageal reflux disease. Gut 1991, 32, 1436–1440. [Google Scholar] [CrossRef] [Green Version]
- Majka, J.; Rembiasz, K.; Migaczewski, M.; Budzynski, A.; Ptak-Belowska, A.; Pabianczyk, R.; Urbanczyk, K.; Zub-Pokrowiecka, A.; Matlok, M.; Brzozowski, T. Cyclooxygenase-2 (COX-2) is the key event in pathophysiology of Barrett’s esophagus. Lesson from experimental animal model and human subjects. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2010, 61, 409–418. [Google Scholar]
- Hu, Z.; Yang, Y.; Zhao, Y.; Huang, Y. The prognostic value of cyclooxygenase-2 expression in patients with esophageal cancer: Evidence from a meta-analysis. Onco Targets Ther. 2017, 10, 2893–2901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- McAdam, E.; Haboubi, H.N.; Forrester, G.; Eltahir, Z.; Spencer-Harty, S.; Davies, C.; Griffiths, A.P.; Baxter, J.N.; Jenkins, G.J.S. Inducible Nitric Oxide Synthase (iNOS) and Nitric Oxide (NO) are Important Mediators of Reflux-induced Cell Signalling in Esophageal Cells. Carcinogenesis 2012, 33, 2035–2043. [Google Scholar] [CrossRef] [Green Version]
- Vannini, F.; Kashfi, K.; Nath, N. The dual role of iNOS in cancer. Redox Biol. 2015, 6, 334–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemons, N.J.; Shannon, N.; Abeyratne, L.R.; Walker, C.E.; Saadi, A.; O’Donovan, M.L.; Lao-Sirieix, P.P.; Fitzgerald, R.C. Nitric oxide-mediated invasion in Barrett’s high-grade dysplasia and adenocarcinoma. Carcinogenesis 2010, 31, 1669–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.D.; Jung, K.H.; Ahn, W.S.; Bae, S.H.; Jang, T.J. Expression of Inducible Nitric Oxide Synthase Is Increased in Rat Barrett’s Esophagus Induced by Duodenal Contents Reflux. J. Korean Med. Sci. 2005, 20, 56–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.; Calrk, E.; Cullen, J.J.; Conklin, J.L. Effect of endotoxin on opossum oesophageal motor function. Neurogastroenterol. Motil. 2000, 12, 215–221. [Google Scholar] [CrossRef]
- Fan, Y.-P.; Chakder, S.; Gao, F.; Rattan, S. Inducible and neuronal nitric oxide synthase involvement in lipopolysaccharide-induced sphincteric dysfunction. Am. J. Physiol. Liver Physiol. 2001, 280, G32–G42. [Google Scholar] [CrossRef]
- Park, H.; Clark, E.; Cullen, J.J.; Koland, J.G.; Kim, M.S.; Conklin, J.L. Expression of inducible nitric oxide synthase in the lower esophageal sphincter of the endotoxemic opossum. J. Gastroenterol. 2002, 37, 1000–1004. [Google Scholar] [CrossRef]
- Feng, Y.; Huang, Y.; Wang, Y.; Wang, P.; Song, H.; Wang, F. Antibiotics induced intestinal tight junction barrier dysfunction is associated with microbiota dysbiosis, activated NLRP3 inflammasome and autophagy. PLoS ONE 2019, 14, e0218384. [Google Scholar] [CrossRef]
- Jo, E.-K.; Kim, J.K.; Shin, D.-M.; Sasakawa, C. Molecular mechanisms regulating NLRP3 inflammasome activation. Cell. Mol. Immunol. 2016, 13, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; Zhang, C.; Xing, Y.; Xue, G.; Zhang, Q.; Pan, F.; Wu, G.; Hu, Y.; Guo, Q.; Lu, A.; et al. Remodelling of the gut microbiota by hyperactive NLRP3 induces regulatory T cells to maintain homeostasis. Nat. Commun. 2017, 8, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Hirota, S.A.; Ng, J.; Lueng, A.; Khajah, M.; Parhar, K.K.S.; Li, Y.; Lam, V.; Potentier, M.S.; Ng, K.; Bawa, M.; et al. NLRP3 inflammasome plays a key role in the regulation of intestinal homeostasis. Inflamm. Bowel Dis. 2011, 17, 1359–1372. [Google Scholar] [CrossRef]
- Li, X.; He, C.; Li, N.; Ding, L.; Chen, H.; Wan, J.; Yang, X.; Xia, L.; He, W.; Xiong, H.; et al. The interplay between the gut microbiota and NLRP3 activation affects the severity of acute pancreatitis in mice. Gut Microbes 2020, 11, 1774–1789. [Google Scholar] [CrossRef] [PubMed]
- Nadatani, Y.; Huo, X.; Zhang, X.; Yu, C.; Cheng, E.; Zhang, Q.; Dunbar, K.B.; Theiss, A.; Pham, T.H.; Wang, D.H.; et al. NOD-Like Receptor Protein 3 Inflammasome Priming and Activation in Barrett’s Epithelial Cells. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 439–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moossavi, M.; Parsamanesh, N.; Bahrami, A.; Atkin, S.L.; Sahebkar, A. Role of the NLRP3 inflammasome in cancer. Mol. Cancer 2018, 17, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Fu, Y.; Tian, D.; Yan, W. The contrasting roles of inflammasomes in cancer. Am. J. Cancer Res. 2018, 8, 566–583. [Google Scholar]
- Baker, K.J.; Houston, A.; Brint, E. IL-1 Family Members in Cancer; Two Sides to Every Story. Front. Immunol. 2019, 10, 1197. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Yin, J.; Miao, J.; Li, S.; Huang, C.; Huang, N.; Fan, T.; Li, X.; Wang, Y.; Han, S.; et al. Activation of NLRP3 inflammasome promotes the proliferation and migration of esophageal squamous cell carcinoma. Oncol. Rep. 2020, 43, 1113–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San-Millán, I.; Brooks, G.A. Reexamining cancer metabolism: Lactate production for carcinogenesis could be the purpose and explanation of the Warburg Effect. Carcinogenesis 2016, 38, 119–133. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, L.; Shen, J.; Cao, B.; Cheng, T.; Zhao, T.; Liu, X.; Zhang, H. Metabolic signatures of esophageal cancer: NMR-based metabolomics and UHPLC-based focused metabolomics of blood serum. Biochim. Biophys. Acta Mol. Basis Dis. 2013, 1832, 1207–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huhta, H.; Helminen, O.; Palomäki, S.; Kauppila, J.H.; Saarnio, J.; Lehenkari, P.; Karttunen, T. Intratumoral lactate metabolism in Barrett’s esophagus and adenocarcinoma. Oncotarget 2017, 8, 22894–22902. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Bowers, J.; Liu, L.; Wei, S.; Gowda, G.A.N.; Hammoud, Z.; Raftery, D. Esophageal Cancer Metabolite Biomarkers Detected by LC-MS and NMR Methods. PLoS ONE 2012, 7, e30181. [Google Scholar] [CrossRef]
- Selling, J.; Swann, P.; Madsen, L.R., 2nd; Oswald, J. Improvement in Gastroesophageal Reflux Symptoms from a Food-grade Maltosyl-isomaltooligosaccharide Soluble Fiber Supplement: A Case Series. Integr. Med. 2018, 17, 40–42. Available online: https://pubmed.ncbi.nlm.nih.gov/31043918/ (accessed on 12 June 2021).
- Cheng, J.; Ouwehand, A.C. Gastroesophageal Reflux Disease and Probiotics: A Systematic Review. Nutrients 2020, 12, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Lin, C.; Yu, J.; Qi, Q.; Wang, Q. Bioengineered Escherichia coli Nissle 1917 for tumour-targeting therapy. Microb. Biotechnol. 2020, 13, 629–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrega, M.J.; Rodríguez-Nogales, A.; Mesa, J.G.; Algieri, F.; Badía, J.; Giménez, R.; Galvez, J.; Baldomà, L. Intestinal Anti-inflammatory Effects of Outer Membrane Vesicles from Escherichia coli Nissle 1917 in DSS-Experimental Colitis in Mice. Front. Microbiol. 2017, 8, 1274. [Google Scholar] [CrossRef] [PubMed]
- Teng, F.; Nair, S.S.D.; Zhu, P.; Li, S.; Huang, S.; Li, X.; Xu, J.; Yang, F. Impact of DNA extraction method and targeted 16S-rRNA hypervariable region on oral microbiota profiling. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
| Toll-Like Receptor | TLR 1 + TLR 2 | TLR 2 + TLR 6 | TLR 3 | TLR 4 | TLR 5 | TLR 7 | TLR 9 | |
|---|---|---|---|---|---|---|---|---|
| Cellular Expression | Monocytes | + | + | + | + | + | + | |
| Macrophages | + | + | + | + | + | + | ||
| Dendritic cells | + | + | + | + | + | + | ||
| NK cells | + | |||||||
| Mast cells | + | + | ||||||
| B cells | + | + | + | + | + | |||
| T cells | + | + | ||||||
| Esophageal epithelium | + | + | + | + | + | + | + | |
| Membrane Expression | Plasma membrane | Plasma membrane | Endosome | Plasma membrane (Endosomal membrane) | Plasma membrane | Endosomal membrane | Endosomal membrane | |
| PAMPs | Bacterial cell wall Triacyl Lipoproteins | Bacterial cell wall Diacyl lipoproteins Lipoteichoic Acid Zymosan | Viral dsRNA Viral ssRNA | Lipopolysaccharide Mannan Glycoinositolphospholipids Envelope Proteins | Flagellin | Viral ssRNA Imidazoquinolines | Unmethylated CpG DNAViral dsDNA | |
| Downstream Signaling Pathway | MyD88 Dependent | MyD88 Dependent | MyD88 Independent | MyD88 Dependent (MyD88 Independent) | MyD88 Dependent | MyD88 Dependent | MyD88 Dependent | |
| Downstream Signaling Factors | NFKB | + | + | + | + (+) | + | + | + |
| CREB | + | + | + | + | + | |||
| AP-1 | + | + | + | + | ||||
| IRF | IRF5 | IRF5 | IRF3, IRF7 | IRF5 IRF3, IRF7 (+) | IRF5 | IRF7 | IRF5, IRF7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gillespie, M.R.; Rai, V.; Agrawal, S.; Nandipati, K.C. The Role of Microbiota in the Pathogenesis of Esophageal Adenocarcinoma. Biology 2021, 10, 697. https://doi.org/10.3390/biology10080697
Gillespie MR, Rai V, Agrawal S, Nandipati KC. The Role of Microbiota in the Pathogenesis of Esophageal Adenocarcinoma. Biology. 2021; 10(8):697. https://doi.org/10.3390/biology10080697
Chicago/Turabian StyleGillespie, Megan R., Vikrant Rai, Swati Agrawal, and Kalyana C. Nandipati. 2021. "The Role of Microbiota in the Pathogenesis of Esophageal Adenocarcinoma" Biology 10, no. 8: 697. https://doi.org/10.3390/biology10080697