
Nutritional Enhancement of Health Beneficial Omega-3 Long-Chain Polyunsaturated Fatty Acids in the Muscle, Liver, Kidney, and Heart of Tattykeel Australian White MARGRA Lambs Fed Pellets Fortified with Omega-3 Oil in a Feedlot System

 , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Dietary Treatments, and Experimental Design
2.2. Feed Sample Processing and Nutrient Composition Analysis
2.3. Fatty Acid Analysis
2.4. Statistical Analyses
3. Results
3.1. Fatty Acid Composition of Basal and Supplementary Feeds
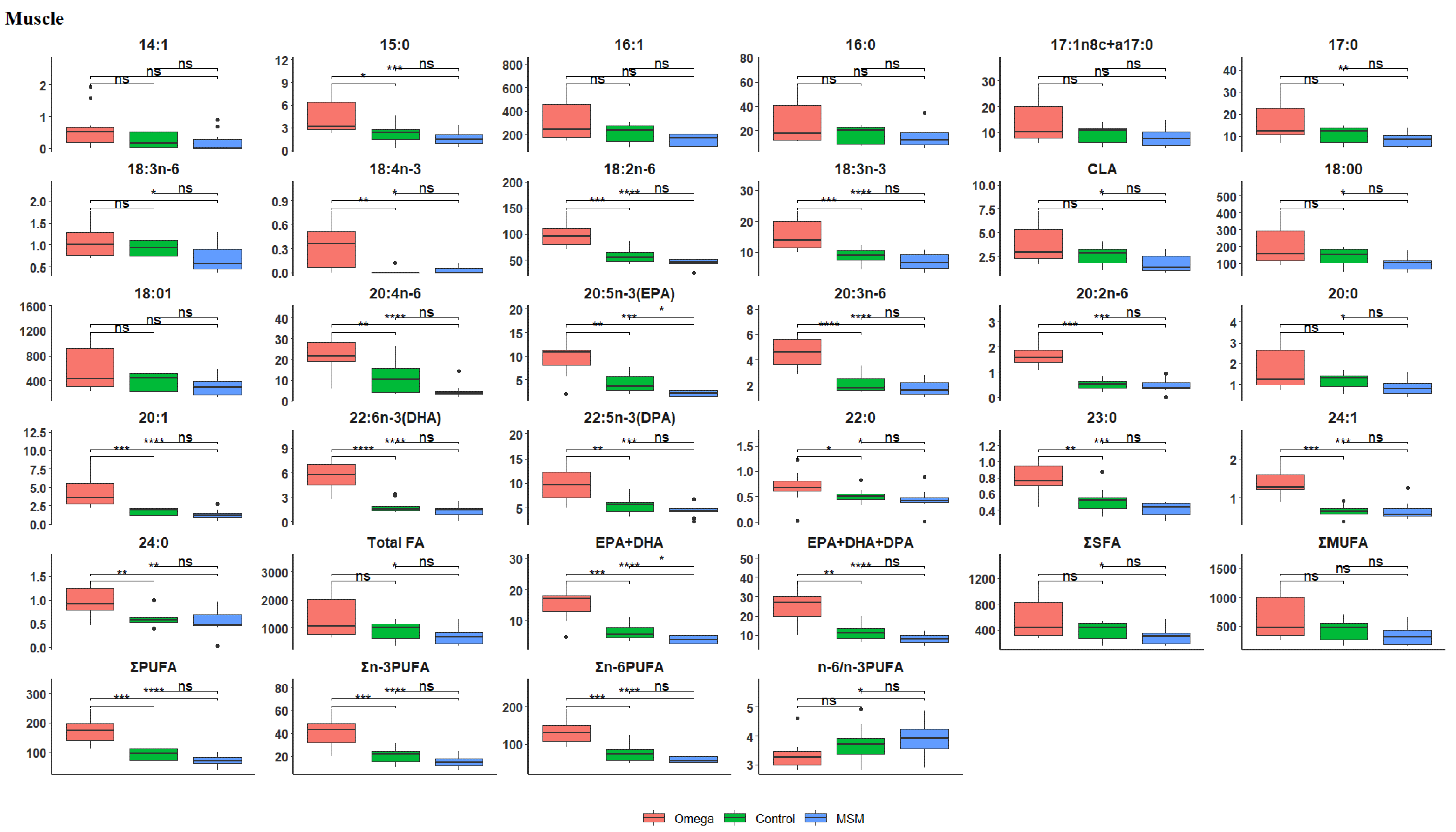
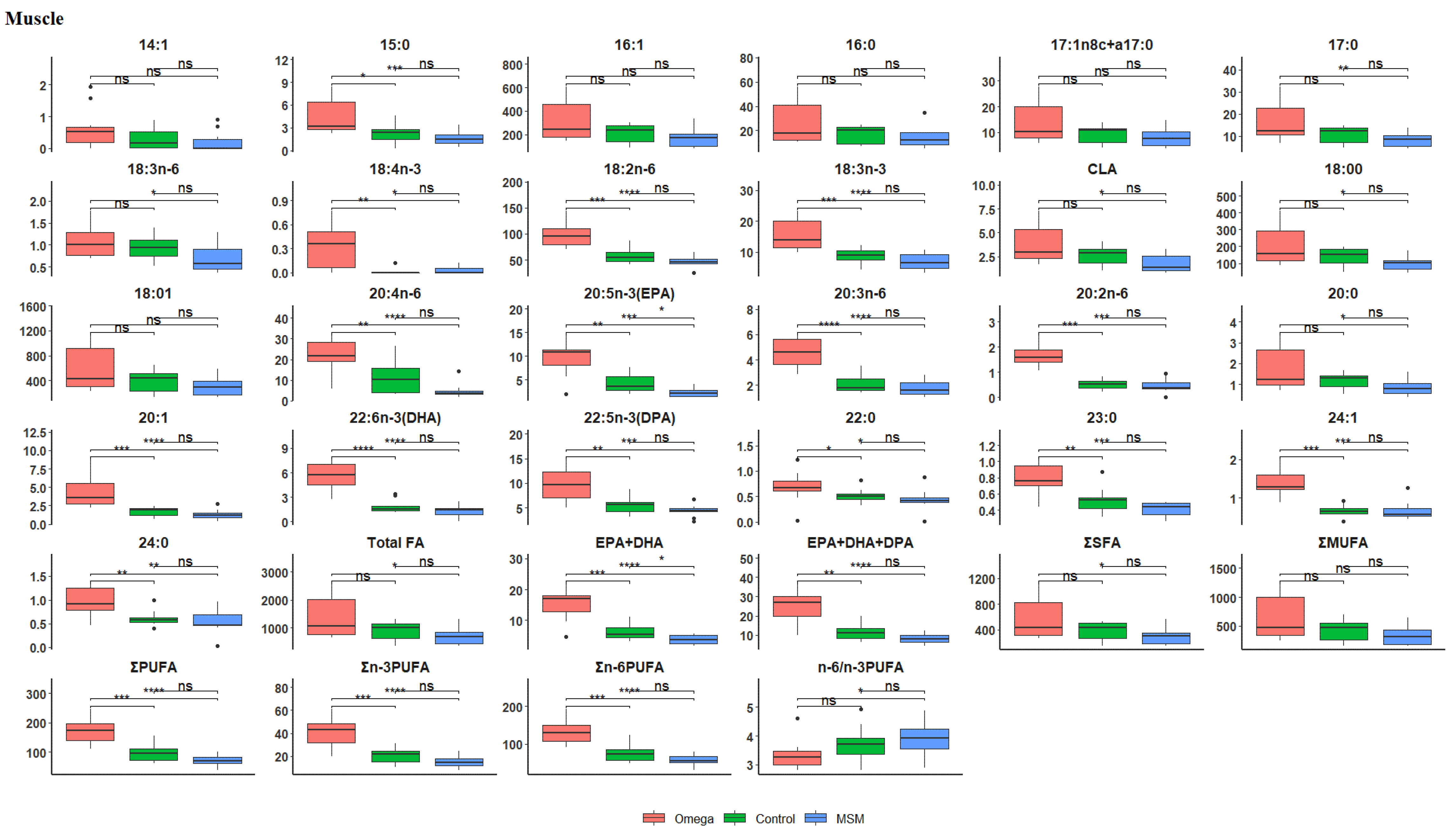
3.2. Fatty Acid Profile of the Longissimus thoracis et lumborum Muscle
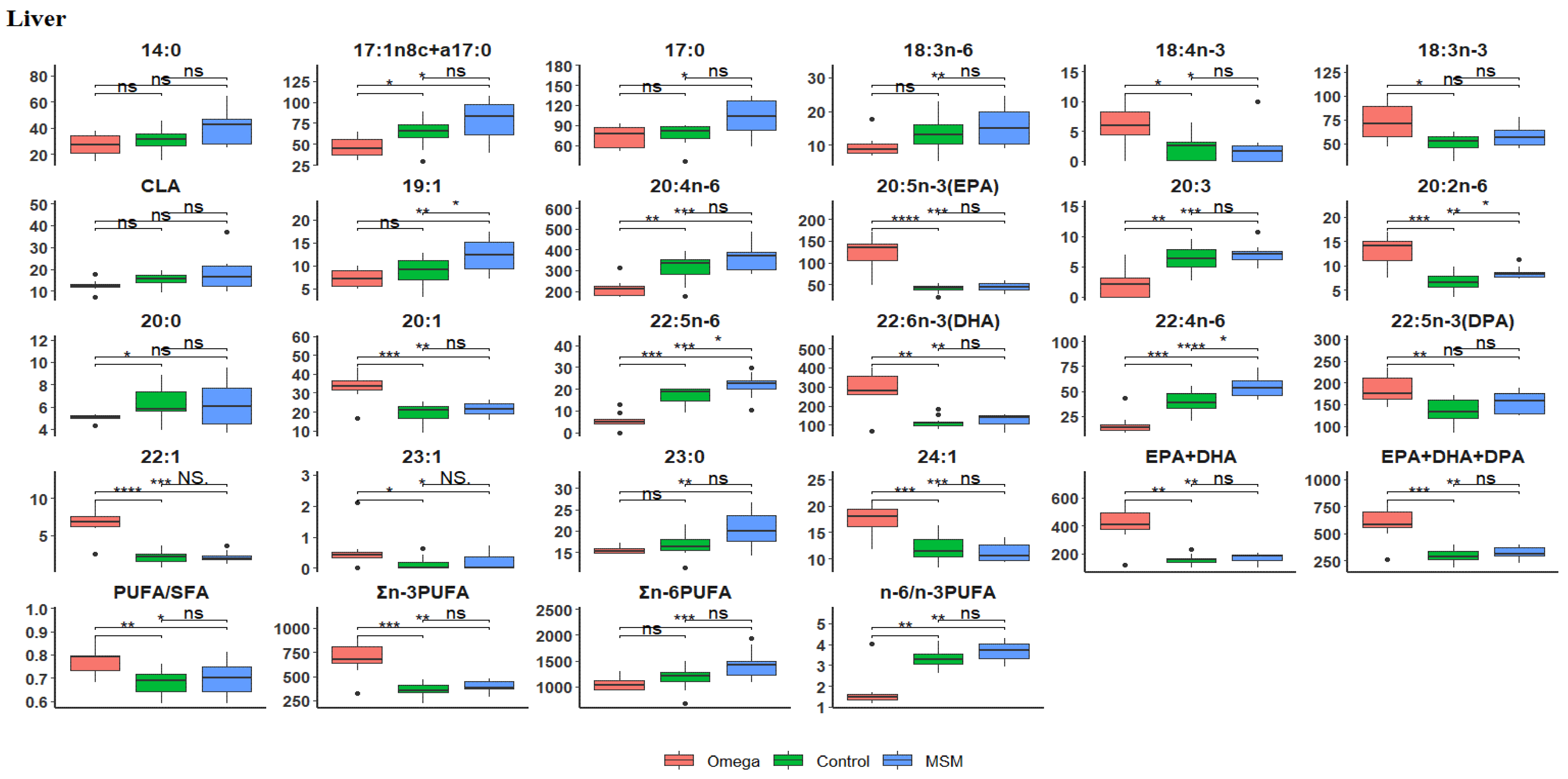
3.3. Fatty Acid Content of Liver
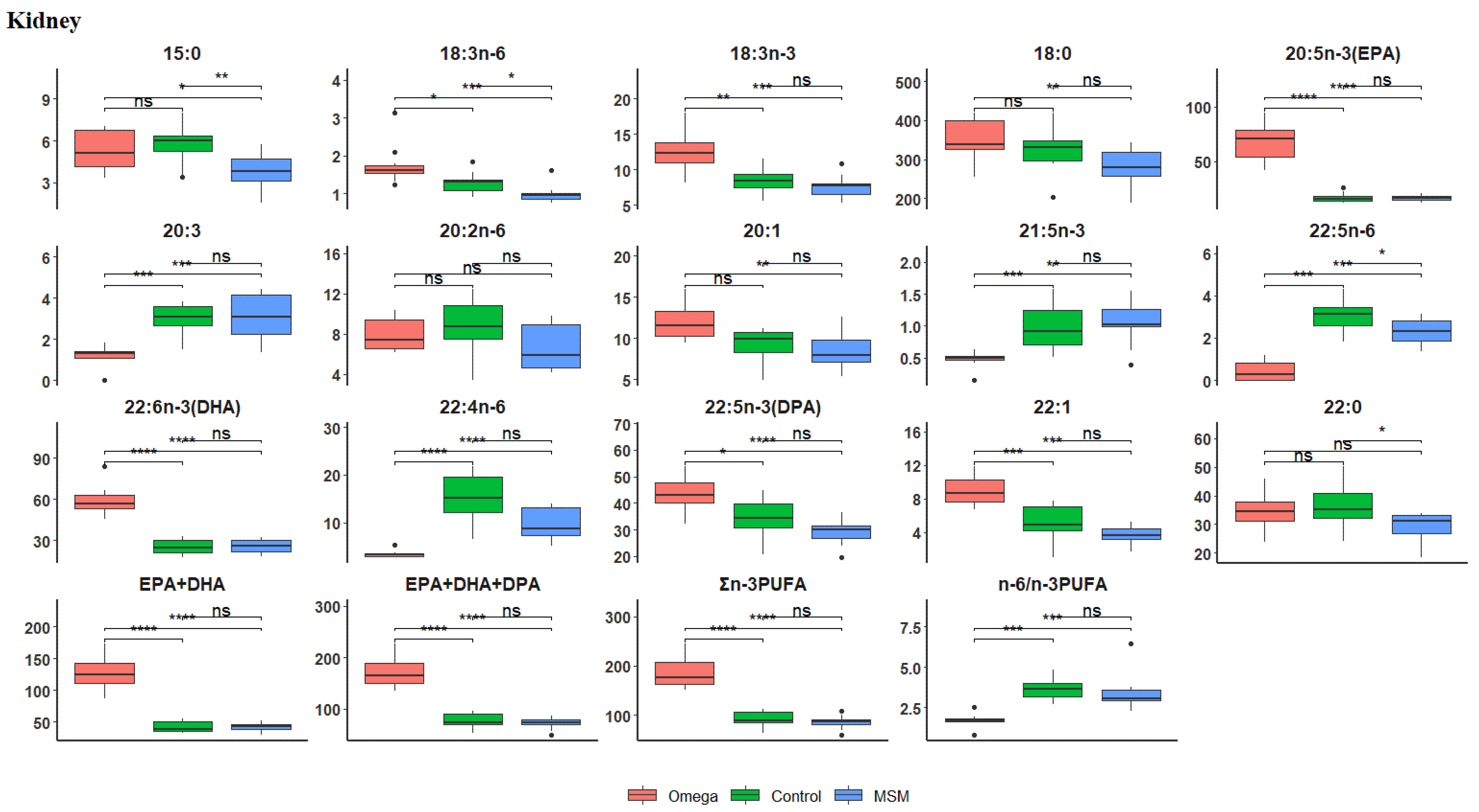
3.4. Fatty Acid Profile of the Kidney
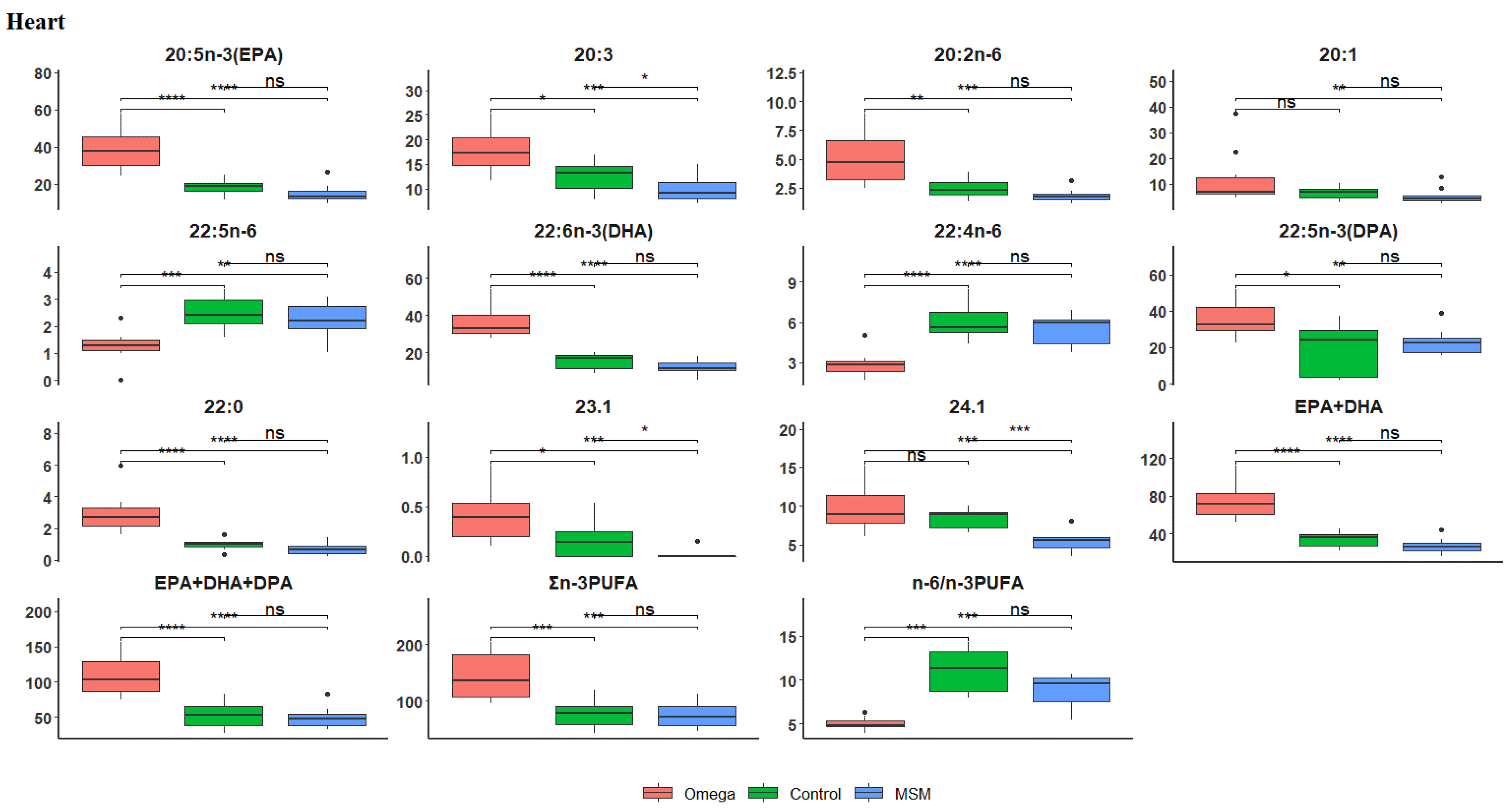
3.5. Fatty Acid Profile of the Heart
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boncinelli, F.; Piracci, G.; Casini, L. Understanding the role of information and taste heterogeneity in consumer preferences for functional beef: The case of the omega-3 enriched burger. Meat Sci. 2021, 181, 108614. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Perez-Alvarez, J.A.; Pateiro, M.; Viuda-Matos, M.; Fernandez-Lopez, J.; Lorenzo, J.M. Satiety from healthier and functional foods. Trends Food Sci. Technol. 2021, 113, 397–410. [Google Scholar] [CrossRef]
- Ansorena, D.; Astiasarán, I. Enrichment of meat products with omega-3 fatty acids by methods other than modification of animal diet. In Food Enrichment with Omega-3 Fatty Acids; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Sawston, UK, 2013; Chapter 10; pp. 299–318. [Google Scholar]
- Morsy, M.K.; Elsabagh, R. Quality parameters and oxidative stability of functional beef burgers fortified with microencapsulated cod liver oil. LWT Food Sci. Technol. 2021, 142, 110959. [Google Scholar] [CrossRef]
- Barros, J.C.; Munekata, P.E.S.; Carvalho, F.A.L.; Dominguez, R.; Trindade, M.A.; Pateiro, M.; Lorenzo, J.M. Healthy beef burgers: Effect of animal fat replacement by algal and wheat germ oil emulsions. Meat Sci. 2021, 173, 108396. [Google Scholar] [CrossRef]
- Van Vliet, S.; Kronberg, S.L.; Provenza, F.D. Plant-based meats, human health, and climate change. Front. Sustain. Food Syst. 2020, 4, 128. [Google Scholar] [CrossRef]
- Bohrer, B.M. Nutrient density and nutritional value of meat products and non-meat foods high in protein. Trends Food Sci. Technol. 2017, 65, 103–112. [Google Scholar] [CrossRef]
- Juárez, M.; Lam, S.; Bohrer, B.M.; Dugan, M.E.; Vahmani, P.; Aalhus, J.; Juárez, A.; López-Campos, O.; Prieto, N.; Segura, J. Enhancing the nutritional value of red meat through genetic and feeding strategies. Foods 2021, 10, 872. [Google Scholar] [CrossRef]
- Fu, Y.; Wang, Y.; Gao, H.; Li, D.; Jiang, R.; Ge, L.; Tong, C.; Xu, K. Associations among dietary omega-3 polyunsaturated fatty acids, the gut microbiota, and intestinal immunity. Mediat. Inflamm. 2021, 2021, 8879227. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Sinclair, A.J.; Holman, B.W. The sources, synthesis and biological actions of omega-3 and omega-6 fatty acids in red meat: An overview. Foods 2021, 10, 1358. [Google Scholar] [CrossRef]
- Rizos, E.C.; Markozannes, G.; Tsapas, A.; Mantzoros, C.S.; Ntzani, E.E. Omega-3 supplementation and cardiovascular disease: Formulation-based systematic review and meta-analysis with trial sequential analysis. Heart 2021, 107, 150–158. [Google Scholar] [CrossRef]
- Chappus-McCendie, H.; Chevalier, L.; Roberge, C.; Plourde, M. Omega-3 PUFA metabolism and brain modifications during aging. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 94, 109662. [Google Scholar] [CrossRef] [Green Version]
- Suito, T.; Nagao, K.; Takeuchi, K.; Juni, N.; Hara, Y.; Umeda, M. Functional expression of Δ12 fatty acid desaturase modulates thermoregulatory behaviour in Drosophila. Sci. Rep. 2020, 10, 11798. [Google Scholar] [CrossRef]
- Mazzocchi, A.; De Cosmi, V.; Risé, P.; Milani, G.P.; Turolo, S.; Syrén, M.-L.; Sala, A.; Agostoni, C. Bioactive compounds in edible oils and their role in oxidative stress and inflammation. Front. Physiol. 2021, 12, 659551. [Google Scholar] [CrossRef]
- Núñez-Sánchez, N.; Avilés Ramírez, C.; Peña Blanco, F.; Gómez-Cortés, P.; de la Fuente, M.Á.; Vioque Amor, M.; Horcada Ibáñez, A.; Martínez Marín, A.L. Effects of algae meal supplementation in feedlot lambs with competent reticular groove reflex on growth performance, carcass traits and meat characteristics. Foods 2021, 10, 857. [Google Scholar] [CrossRef] [PubMed]
- Sharafi, Y.; Majidi, M.M.; Goli, S.A.H.; Rashidi, F. Oil content and fatty acids composition in Brassica species. Int. J. Food Prop. 2015, 18, 2145–2154. [Google Scholar] [CrossRef] [Green Version]
- Jokić, S.; Sudar, R.; Svilović, S.; Vidović, S.; Bilić, M.; Velić, D.; Jurković, V. Fatty acid composition of oil obtained from soybeans by extraction with supercritical carbon dioxide. Czech J. Food Sci. 2013, 31, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Giuffrè, A.M.; Capocasale, M. Physicochemical composition of tomato seed oil for an edible use: The effect of cultivar. Int. Food Res. J. 2016, 23, 583–591. [Google Scholar]
- Angelo, M.G.; Nobile, R. Citrus bergamia, Risso: The peel, the juice and the seed oil of the bergamot fruit of Reggio Calabria (South Italy). Emir. J. Food Agric. 2020, 32, 522–532. [Google Scholar]
- Gonzales-Barron, U.; Popova, T.; Piedra, R.B.; Tolsdorf, A.; Geß, A.; Pires, J.; Domínguez, R.; Chiesa, F.; Brugiapaglia, A.; Viola, I.; et al. Fatty acid composition of lamb meat from Italian and German local breeds. Small Rum. Res. 2021, 200, 106384. [Google Scholar] [CrossRef]
- Hoffman, L.C.; Claasen, B.; der Merwe, V.; André, D.; Cloete, S.W.P.; Cloete, J.J.E. The effects of production system and sex on the sensory quality characteristics of Dorper lamb. Foods 2020, 9, 725. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Nguyen, D.V.; Le, H.V.; Nguyen, Q.V.; Otto, J.R.; Malau-Aduli, B.S.; Nichols, P.D. Correlations between growth and wool quality traits of genetically divergent Australian lambs in response to canola or flaxseed oil supplementation. PLoS ONE 2019, 14, e0208229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhuiyan, M.; Lee, D.; Kim, H.; Lee, S.; Cho, S.; Yang, B.; Kim, S.D.; Lee, S.H. Estimates of genetic parameters for fatty acid compositions in the longissimus dorsi muscle of Hanwoo cattle. Animal 2018, 12, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Junior, F.M.V.; Martins, C.F.; Feijó, G.L.D.; Teixeira, A.; Leonardo, A.P.; de Almeida Ricardo, H.; Fernandez, A.R.M.; Reis, F.A. Evaluation of genotype on fatty acid profile and sensory of meat of indigenous Pantaneiro sheep and Texel or Santa Inês crossbred finished on feedlot. Small Rum. Res. 2019, 173, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Sarı, M.; Aksoy, Y.; Önk, K.; Erinç, H.; Işık, S.A.; Tilki, M. Effects of genotype and fattening system on the quality of male lamb meat–Part 1: Technological properties and carcass measurements. Arch. Anim. Breed 2019, 62, 605–614. [Google Scholar] [CrossRef] [Green Version]
- Belaunzaran, X.; Lavín, P.; Mantecón, A.; Kramer, J.; Aldai, N. Effect of slaughter age and feeding system on the neutral and polar lipid composition of horse meat. Animal 2018, 12, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Miguel, E.; Blázquez, B.; Ruiz de Huidobro, F. Liveweight and sex effects on sensory quality of Rubia de El Molar autochthonous ovine breed meat. Animals 2021, 11, 1293. [Google Scholar] [CrossRef]
- Matar, A.M.; Abdelrahman, M.M.; Alhidary, I.A.; Ayadi, M.A.; Alobre, M.M.; Aljumaah, R.S. Effects of roughage quality and particle size on rumen parameters and fatty acid profiles of Longissimus dorsi fat of lambs fed complete feed. Animals 2020, 10, 2182. [Google Scholar] [CrossRef]
- Wood, J.; Enser, M. Manipulating the fatty acid composition of meat to improve nutritional value and meat quality. In New Aspects of Meat Quality; Peter, P.P., Ed.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Sawston, UK, 2017; Volume 501–535, 744p. [Google Scholar]
- Vahmani, P.; Ponnampalam, E.N.; Kraft, J.; Mapiye, C.; Bermingham, E.N.; Watkins, P.J.; Proctor, S.D.; Dugan, M.E.R. Bioactivity and health effects of ruminant meat lipids. Invited Review. Meat Sci. 2020, 165, 108114. [Google Scholar] [CrossRef]
- Garcia-Galicia, I.A.; Arras-Acosta, J.A.; Huerta-Jimenez, M.; Rentería-Monterrubio, A.L.; Loya-Olguin, J.L.; Carrillo-Lopez, L.M.; Tirado-Gallegos, J.M.; Alarcon-Rojo, A.D. Natural oregano essential oil may replace antibiotics in lamb diets: Effects on meat quality. Antibiotics 2020, 9, 248. [Google Scholar] [CrossRef]
- Dewanckele, L.; Vlaeminck, B.; Hernandez-Sanabria, E.; Ruiz-González, A.; Debruyne, S.; Jeyanathan, J.; Veerle, F. Rumen biohydrogenation and microbial community changes upon early life supplementation of 22: 6n-3 enriched microalgae to goats. Front. Microbiol. 2018, 9, 573. [Google Scholar] [CrossRef] [Green Version]
- Holman, B.W.B.; Kerr, M.J.; Refshauge, G.; Diffey, S.M.; Hayes, R.C.; Newell, M.T.; Hopkins, D.L. Post-mortem pH decline in lamb semitendinosus muscle and its relationship to the pH decline parameters of the longissimus lumborum muscle: A pilot study. Meat Sci. 2021, 176, 108473. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis of AOAC International, 16th ed.; AOAC International: Gaithersburg, MD, USA, 1995. [Google Scholar]
- Malau-Aduli, A.E.O.; Holman, B.W.B.; Kashani, A.; Nichols, P.D. Sire breed and sex effects on the fatty acid composition and content of heart, kidney, liver, adipose and muscle tissues of purebred and first-cross prime lambs. Anim. Prod. Sci. 2016, 56, 2122–2132. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Siebert, B.D.; Bottema, C.D.K.; Pitchford, W.S. Breed comparison of the fatty acid composition of muscle phospholipids in Jersey and Limousin cattle. J. Anim. Sci. 1998, 76, 766–773. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.R.; Nichols, P.D.; Barnes, J.; Davies, N.W.; Peacock, E.J.; Carter, C.G. Regiospecificity profiles of storage and membrane lipids from the gill and muscle tissue of Atlantic salmon (Salmo salar L.) grown at elevated temperature. Lipids 2006, 41, 865–876. [Google Scholar] [CrossRef]
- Clayton, E. Graham Centre Monograph no. 4: Long-Chain Omega-3 Polyunsaturated Fatty Acids in Ruminant Nutrition: Benefits to Animals and Humans; Nugent, T., Nicholls, C., Eds.; New South Wales Department of Primary Industries: Wagga Wagga, Australia, 2014; ISBN 978 1 74256 678 8. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing, Rstudio version 1.3.1056; R Foundation for Statistical Computing: Vienna, Austria, 2021; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 20 August 2021).
- Pohlert, T. The pairwise multiple comparison of mean ranks package (PMCMR). R Package 2014, 27, 9. [Google Scholar]
- Dunn, O.J. Multiple comparisons using rank sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Benjamin, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Kendall, M.G. A new measure of rank correlation. Biometrika 1938, 30, 81–93. [Google Scholar] [CrossRef]
- Al-Shaar, L.; Satija, A.; Wang, D.D.; Rimm, E.B.; Smith-Warner, S.A.; Stampfer, M.J.; Hu, F.B.; Willett, W.C. Red meat intake and risk of coronary heart disease among US men: Prospective cohort study. BMJ 2020, 371, m4141. [Google Scholar] [CrossRef]
- Jo, G.; Oh, H.; Singh, G.M.; Park, D.; Shin, M.-J. Impact of dietary risk factors on cardiometabolic and cancer mortality burden among Korean adults: Results from nationally representative repeated cross-sectional surveys 1998–2016. Nutr. Res. Pract. 2020, 14, 384–400. [Google Scholar] [CrossRef]
- Alba, H.D.; Freitas Júnior, J.E.; Leite, L.C.; Azevêdo, J.A.; Santos, S.A.; Pina, D.S.; Cirne, L.G.A.; Rodrigues, C.S.; Silva, W.P.; Lima, V.G.O.; et al. Protected or unprotected fat addition for feedlot lambs: Feeding behavior, carcass traits, and meat quality. Animals 2021, 11, 328. [Google Scholar] [CrossRef]
- Urrutia, O.; Mendizabal, J.A.; Alfonso, L.; Soret, B.; Insausti, K.; Arana, A. Adipose tissue modification through feeding strategies and their implication on adipogenesis and adipose tissue metabolism in ruminants. Int. J. Mol. Sci. 2020, 21, 3183. [Google Scholar] [CrossRef] [PubMed]
- Le, V.H.; Nguyen, D.V.; Nguyen, Q.V.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Fatty acid profiles of muscle, liver, heart and kidney of Australian prime lambs fed different polyunsaturated fatty acids enriched pellets in a feedlot system. Sci. Rep. 2019, 9, 1238. [Google Scholar] [PubMed]
- Nguyen, Q.V.; Malau-Aduli, B.S.; Cavalieri, J.; Nichols, P.D.; Malau-Aduli, A.E.O. Enhancing omega-3 long-chain polyunsaturated fatty acid content of dairy-derived foods for human consumption. Nutrients 2019, 11, 743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.V.; Le, V.H.; Nguyen, Q.V.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Omega–3 long-chain fatty acids in the heart, kidney, liver and plasma metabolite profiles of Australian prime lambs supplemented with pelleted canola and flaxseed oils. Nutrients 2017, 9, 893. [Google Scholar] [CrossRef] [Green Version]
- Celada, P.; Sánchez-Múniz, F.J. Are meat and meat product consumptions harmful? Their relationship with the risk of colorectal cancer and other degenerative diseases. An. Real. Acad. Farm. 2016, 82, 68–90. [Google Scholar]
- Gruffat, D.; Durand, D.; Rivaroli, D.; Do Prado, I.; Prache, S. Comparison of muscle fatty acid composition and lipid stability in lambs stall-fed or pasture-fed alfalfa with or without sainfoin pellet supplementation. Animal 2020, 14, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Renna, M.; Brugiapaglia, A.; Zanardi, E.; Destefanis, G.; Prandini, A.; Moschini, M.; Sigolo, S.; Lussiana, C. Fatty acid profile, meat quality and flavour acceptability of beef from double-muscled Piemontese young bulls fed ground flaxseed. Ital. J. Anim. Sci. 2019, 18, 355–365. [Google Scholar] [CrossRef] [Green Version]
- De Lima Júnior, D.M.; do Nascimento Rangel, A.H.; Urbano, S.A.; Moreno, G.M.B. Oxidação lipídica e qualidade da carne ovina. Acta Vet. Bras. 2013, 7, 14–28. [Google Scholar]
- Pewan, S.B.; Otto, J.R.; Kinobe, R.T.; Adegboye, O.A.; Malau-Aduli, A.E.O. MARGRA lamb eating quality and human health-promoting omega-3 long-chain polyunsaturated fatty acid profiles of Tattykeel Australian White sheep: Linebreeding and gender effects. Antioxidants 2020, 9, 1118. [Google Scholar] [CrossRef] [PubMed]
- Facciolongo, A.M.; Lestingi, A.; Colonna, M.A.; Nicastro, F.; De Marzo, D.; Toteda, F. Effect of diet lipid source (linseed vs. soybean) and gender on performance, meat quality and intramuscular fatty acid composition in fattening lambs. Small Rum. Res. 2018, 159, 11–17. [Google Scholar] [CrossRef]
- Den Hartigh, L.J. Conjugated linoleic acid effects on cancer, obesity, and atherosclerosis: A review of pre-clinical and human trials with current perspectives. Nutrients 2019, 11, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simopoulos, A.P. Genetic variants in the metabolism of omega-6 and omega-3 fatty acids: Their role in the determination of nutritional requirements and chronic disease risk. Exp. Biol. Med. 2010, 235, 785–795. [Google Scholar] [CrossRef]
- Urrutia, O.; Mendizabal, J.A.; Insausti, K.; Soret, B.; Purroy, A.; Arana, A. Effects of addition of linseed and marine algae to the diet on adipose tissue development, fatty acid profile, lipogenic gene expression, and meat quality in lambs. PLoS ONE 2016, 11, e0156765. [Google Scholar] [CrossRef] [Green Version]
- Widmann, P.; Nuernberg, K.; Kuehn, C.; Weikard, R. Association of an ACSL1 gene variant with polyunsaturated fatty acids in bovine skeletal muscle. BMC Genet. 2011, 12, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Lee, H.; Kang, S.; Park, W.J. Fatty acid desaturases, polyunsaturated fatty acid regulation, and biotechnological advances. Nutrients 2016, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Mueller-Harvey, I.; Bee, G.; Dohme-Meier, F.; Hoste, H.; Karonen, M.; Kölliker, R.; Lüscher, A.; Niderkorn, V.; Pellikaan, W.F.; Salminen, J.P.; et al. Benefits of condensed tannins in forages fed to ruminants: Importance of structure, concentration and diet. Crop. Sci. 2019, 59, 861–885. [Google Scholar] [CrossRef] [Green Version]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef]
- Freitas, N.; Araújo, M.; Oliveira, R.; Lanna, D.; Marques, C.; Torreão, J.; Santos, C.B.; Silva Junior, J.M.; Edvan, R.L.; Bezerra, L.R. Production, composition, fatty acid profile and sensory traits of milk from goats fed crude glycerin from waste frying oils used in biodiesel production. Livest. Sci. 2020, 238, 104060. [Google Scholar] [CrossRef]
- Wood, J.; Enser, M.; Fisher, A.; Nute, G.; Sheard, P.; Richardson, R.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Lee, S.; Lee, J.; Choi, I.J.; Kim, Y.-W.; Ryu, K.W.; Kim, Y.-I.; Kim, J. Dietary n-3 and n-6 polyunsaturated fatty acids, the FADS gene, and the risk of gastric cancer in a Korean population. Sci. Rep. 2018, 8, 3823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C. n− 3 Polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, 1505S–1519S. [Google Scholar] [CrossRef] [PubMed]
- Yates, C.M.; Calder, P.C.; Rainger, G.E. Pharmacology and therapeutics of omega-3 polyunsaturated fatty acids in chronic inflammatory disease. Pharmacol. Ther. 2014, 141, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Ren, X.-L.; Fu, Y.-Q.; Gao, J.-L.; Li, D. Ratio of n-3/n-6 PUFAs and risk of breast cancer: A meta-analysis of 274135 adult females from 11 independent prospective studies. BMC Cancer 2014, 14, 105. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Choi, J.; Alpergin, E.S.S.; Zhao, L.; Hartung, T.; Scafidi, S.; Riddle, R.C.; Wolfgang, M.J. Loss of hepatic mitochondrial long-chain fatty acid oxidation confers resistance to diet-induced obesity and glucose intolerance. Cell Rep. 2017, 20, 655–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo Vargas, J.A. Metabolism and function of lipids in the adipose and liver tissues of production ruminants: A review. CES Med. Vet. Zootec. 2019, 14, 30–44. [Google Scholar] [CrossRef] [Green Version]
- Byelashov, O.A.; Sinclair, A.J.; Kaur, G. Dietary sources, current intakes, and nutritional role of omega-3 docosapentaenoic acid. Lipid Technol. 2015, 27, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Kuhnt, K.; Degen, C.; Jahreis, G. Evaluation of the impact of ruminant trans fatty acids on human health: Important aspects to consider. Crit. Rev. Food Sci. Nutr. 2016, 56, 1964–1980. [Google Scholar] [CrossRef]
- Hennessy, A.A.; Ross, R.P.; Devery, R.; Stanton, C. The health promoting properties of the conjugated isomers of α-linolenic acid. Lipids 2011, 46, 105–119. [Google Scholar] [CrossRef]
- Gómez, I.; Mendizabal, J.; Sarriés, M.; Insausti, K.; Albertí, P.; Realini, C.; Perez-Juan, M.; Oliver, M.A.; Purroy, A.; Beriain, M.J. Fatty acid composition of young Holstein bulls fed whole linseed and rumen-protected conjugated linoleic acid enriched diets. Livest. Sci. 2015, 180, 106–112. [Google Scholar] [CrossRef]
- Guerrero, A.; Sañudo, C.; Campo, M.; Olleta, J.; Muela, E.; Macedo, R.; Macedo, F.A.F. Effect of linseed supplementation level and feeding duration on performance, carcass and meat quality of cull ewes. Small Rum. Res. 2018, 167, 70–77. [Google Scholar] [CrossRef]
- Moibi, J.A.; Christopherson, R.J. Effect of environmental temperature and a protected lipid supplement on the fatty acid profile of ovine longissimus dorsi muscle, liver and adipose tissues. Livest. Prod. Sci. 2001, 69, 245–254. [Google Scholar] [CrossRef]
- Borowiec, F.; Micek, P.; Marcinski, M.; Barteczko, J.; Zajac, T. Linseed-based diets for sheep. 2. Performance and chemical composition of meat and liver. J. Anim. Feed Sci. 2004, 13, 19–22. [Google Scholar] [CrossRef] [Green Version]
- Coleman, D.N.; Martin, A.C.C.; Jin, Y.; Lee, K.; Relling, A.E. Prepartum fatty acid supplementation in sheep. IV. Effect of calcium salts with eicosapentaenoic acid and docosahexaenoic acid in the maternal and finishing diet on lamb liver and adipose tissue during the lamb finishing period. J. Anim. Sci. 2019, 97, 3071–3088. [Google Scholar] [CrossRef]
- Bernacka, H.; Peter, E.; Mistrzak, M. Fatty acid content in meat, heart, and liver of conventionally bred Polish Merino lambs. Fatty acid content in meat, heart, and liver of conventionally bred Polish Merino lambs. Med. Weter. 2013, 69, 424–427. [Google Scholar]
- Demirel, G.; Wood, J.D.; Enser, M. Conjugated linoleic acid content of the lamb muscle and liver fed different supplements. Small Rum. Res. 2004, 53, 23–28. [Google Scholar] [CrossRef]
- Kim, S.C.; Adesogan, A.T.; Badinga, L.; Staples, C.R. Effects of dietary n-6: N-3 fatty acid ratio on feed intake, digestibility, and fatty acid profiles of the ruminal contents, liver, and muscle of growing lambs. J. Anim. Sci. 2007, 85, 706–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biel, W.; Czerniawska-Piątkowska, E.; Kowalczyk, A. Offal chemical composition from veal, beef, and lamb maintained in organic production systems. Animals 2019, 9, 489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florek, M.; Litwińczuk, Z.; Skałecki, P.; Kędzierska-Matysek, M.; Grodzicki, T. Chemical composition and inherent properties of offal from calves maintained under two production systems. Meat Sci. 2012, 90, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Hicks, T.M.; Knowles, S.O.; Farouk, M.M. Global provisioning of red meat for flexitarian diets. Front. Nutr. 2018, 5, 50. [Google Scholar] [CrossRef]
- Mashek, D.G.; Coleman, R.A. Cellular fatty acid uptake: The contribution of metabolism. Curr. Opin. Lipidol. 2006, 17, 274–278. [Google Scholar] [CrossRef]
- Hagve, T.A.; Woldseth, B.; Brox, J.; Narce, M.; Poisson, J.P. Membrane fluidity and fatty acid metabolism in kidney cells from rats fed purified eicosapentaenoic acid or purified docosahexaenoic acid. Scand. J. Clin. Lab. Investig. 1998, 58, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Liabo, J.; Odden, N.; Christiansen, E.N.; Hagve, T.A. Metabolism of long-chain polyunsaturated fatty acids in rat kidney cells. Ann. Nutr. Metab. 2003, 47, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Schaap, F.G.; van der Vusse, G.J.; Glatz, J.F.C. Fatty acid-binding proteins in the heart. Card. Metab. Health Dis. 1998, 180, 43–51. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient Composition (%DM) | Experimental and Basal Diets | |||
|---|---|---|---|---|
| Control | Omega-3 | MSM Whole Grain | Hay | |
| Dry Matter (DM) | 90.8 | 91.7 | 90.3 | 93.4 |
| Moisture | 9.2 | 8.3 | 9.7 | 6.6 |
| Acid Detergent Fiber (ADF) | 7.6 | 8.2 | 6.3 | 39.4 |
| Neutral Detergent Fiber (NDF) | 23.8 | 23.0 | 21.8 | 60.9 |
| Crude Protein (CP) | 16.9 | 17.0 | 16.4 | 7.5 |
| Ash | 7.8 | 8.2 | 7.2 | 8.1 |
| Ether Extract (EE) | 6.1 | 10.3 | 6.0 | 3.3 |
| Metabolizable Energy (ME) MJ/kg | 14.1 | 15.1 | 14.4 | 8.3 |
| Dry Matter Digestibility (DMD) | 84.9 | 83.8 | 87.5 | 46.8 |
| Digestible Organic Matter (DOMD) | 83.7 | 82.6 | 86.2 | 47.1 |
| Fatty Acid | Omega-3 | Control | MSM Whole Grain | Basal Hay |
|---|---|---|---|---|
| 13:0 | 0.00 | 0.00 | 0.01 | 0.00 |
| 14:1 | 0.00 | 0.00 | 0.00 | 0.00 |
| 14:0 | 2.23 | 0.45 | 0.25 | 0.33 |
| 15:0 | 2.62 | 0.36 | 0.30 | 0.21 |
| 16:1 | 1.02 | 0.79 | 1.05 | 1.88 |
| 16:0 | 35.96 | 27.19 | 30.52 | 12.53 |
| 17:1n8c + a17:0 | 0.64 | 0.27 | 0.25 | 0.18 |
| 17:0 | 1.09 | 0.33 | 0.30 | 0.13 |
| 18:3n6 | 0.32 | 0.00 | 0.00 | 0.24 |
| 18:4n3 | 0.32 | 0.00 | 0.00 | 0.20 |
| 18:2n6 (LA) | 25.25 | 81.75 | 92.38 | 30.02 |
| 18:3n3 (ALA) | 15.39 | 8.84 | 11.21 | 6.13 |
| CLA | 0.65 | 0.10 | 0.29 | 0.00 |
| 18:0 | 8.25 | 4.59 | 4.13 | 2.74 |
| 18:1 | 31.92 | 57.34 | 75.06 | 51.23 |
| 19:1 | 0.00 | 0.04 | 0.06 | 0.04 |
| 20:4n6 (ARA) | 0.00 | 0.00 | 0.00 | 0.53 |
| 20:5n3 (EPA) | 0.22 | 0.18 | 0.23 | 0.00 |
| 20:3 | 0.00 | 0.00 | 0.01 | 0.00 |
| 20:3n6 | 0.80 | 0.15 | 0.16 | 0.18 |
| 20:4n3 | 0.30 | 0.00 | 0.00 | 0.02 |
| 20:2n6 | 0.32 | 0.13 | 0.21 | 0.33 |
| 20:1 | 1.51 | 1.71 | 2.02 | 1.79 |
| 20:0 | 2.80 | 0.60 | 0.81 | 0.42 |
| 21:5n3 | 0.43 | 0.00 | 0.00 | 0.00 |
| 21:0 | 1.17 | 0.16 | 0.08 | 0.00 |
| 22:5n6 | 0.87 | 0.01 | 0.08 | 0.00 |
| 22:6n3 (DHA) | 1.53 | 0.03 | 0.05 | 0.93 |
| 22:4n6 | 0.60 | 0.08 | 0.05 | 0.00 |
| 22:5n3 (DPA) | 0.99 | 0.00 | 0.17 | 0.22 |
| 22:1 | 2.39 | 0.47 | 0.34 | 0.25 |
| 22:0 | 4.63 | 0.44 | 0.60 | 0.23 |
| 23:1 | 0.64 | 0.06 | 0.00 | 0.00 |
| 23:0 | 1.72 | 0.01 | 0.14 | 0.02 |
| 24:1 | 0.94 | 0.25 | 0.28 | 0.14 |
| 24:0 | 3.54 | 0.40 | 0.54 | 0.15 |
| Total FA | 147.11 | 185.91 | 220.96 | 110.69 |
| EPA + DHA | 1.74 | 0.21 | 0.28 | 0.93 |
| EPA + DHA + DPA | 2.74 | 0.21 | 0.45 | 1.16 |
| SFA | 64.02 | 34.52 | 37.66 | 16.77 |
| MUFA | 35.09 | 60.13 | 78.46 | 55.12 |
| PUFA | 48.00 | 91.26 | 104.84 | 38.81 |
| PUFA/SFA | 0.75 | 2.64 | 2.78 | 2.31 |
| ∑n3PUFA | 19.18 | 9.05 | 11.66 | 7.51 |
| ∑n6PUFA | 28.16 | 82.12 | 92.88 | 31.30 |
| n6/n3PUFA | 1.47 | 9.08 | 7.97 | 4.17 |
| Fatty Acid | Omega-3 | Control | MSM Whole Grain | p-Value |
|---|---|---|---|---|
| 13:0 | 0.05 ± 0.11 | 0.03 ± 0.04 | 0.00 ± 0.00 | 0.1108 |
| 14:1 | 0.63 ± 0.65 | 0.31 ± 0.33 | 0.19 ± 0.34 | 0.0417 |
| 14:0 | 24.25 ± 18.62 | 16.80 ± 7.84 | 14.22 ± 8.06 | 0.0809 |
| 15:1 | 4.44 ± 2.36 | 2.26 ± 1.22 | 1.58 ± 0.85 | 0.0005 |
| 16:0 | 320.50 ± 182.90 | 209.83 ± 83.05 | 169.06 ± 79.43 | 0.0106 |
| 16:1 | 25.86 ± 17.40 | 16.93 ± 7.29 | 14.43 ± 8.57 | 0.0398 |
| 17:1n8c + a17:0 | 13.83 ± 7.99 | 9.40 ± 3.68 | 7.85 ± 3.55 | 0.0201 |
| 17:0 | 16.85 ± 9.12 | 10.80 ± 4.08 | 8.31 ± 3.10 | 0.0035 |
| 18:3n6 | 1.09 ± 0.38 | 0.93 ± 0.28 | 0.71 ± 0.31 | 0.0116 |
| 18:4n-3 | 0.34 ± 0.28 | 0.01 ± 0.04 | 0.03 ± 0.05 | 0.0006 |
| 18:2n-6 (LA) | 99.35 ± 25.12 | 57.06 ± 14.44 | 46.89 ± 10.66 | 0.0000 |
| 18:3n-3 (ALA) | 15.70 ± 5.01 | 8.76 ± 2.46 | 6.87 ± 2.64 | 0.0000 |
| CLA | 3.83 ± 2.05 | 2.65 ± 1.03 | 1.79 ± 0.93 | 0.0030 |
| 18:0 | 208.83 ± 119.80 | 139.11 ± 54.40 | 99.63 ± 40.82 | 0.0042 |
| 18:1 | 582.31 ± 364.95 | 395.67 ± 173.74 | 305.06 ± 147.93 | 0.0174 |
| 19:1 | 1.31 ± 0.68 | 0.89 ± 0.37 | 0.93 ± 0.51 | 0.1222 |
| 20:4n-6 (ARA) | 21.87 ± 8.20 | 11.26 ± 8.18 | 4.78 ± 3.64 | 0.0000 |
| 20:5n-3 (EPA) | 9.68 ± 3.68 | 4.25 ± 2.15 | 2.36 ± 1.01 | 0.0000 |
| 22:3 | 0.19 ± 0.31 | 0.21 ± 0.22 | 0.18 ± 0.23 | 0.9168 |
| 20:3n-6 | 4.61 ± 1.17 | 2.10 ± 0.68 | 1.74 ± 0.58 | 0.0000 |
| 20:4n-3 | 0.02 ± 0.05 | 0.10 ± 0.18 | 0.15 ± 0.24 | 0.0945 |
| 20:2n-6 | 1.66 ± 0.41 | 0.52 ± 0.20 | 0.45 ± 0.29 | 0.0000 |
| 20:0 | 1.71 ± 1.01 | 1.15 ± 0.40 | 0.82 ± 0.37 | 0.0051 |
| 20:1 | 4.45 ± 2.34 | 1.71 ± 0.63 | 1.30 ± 0.68 | 0.0001 |
| 21:5n-3 | 0.15 ± 0.13 | 0.14 ± 0.12 | 0.11 ± 0.12 | 0.4905 |
| 21:0 | 0.13 ± 0.22 | 0.04 ± 0.09 | 0.10 ± 0.14 | 0.5992 |
| 22:5n-6 | 0.27 ± 0.22 | 0.26 ± 0.25 | 0.24 ± 0.11 | 0.6978 |
| 22:6n-3 (DHA) | 5.59 ± 1.63 | 1.85 ± 0.81 | 1.28 ± 0.67 | 0.0000 |
| 22:4n-6 | 1.10 ± 0.39 | 0.93 ± 0.33 | 0.91 ± 0.25 | 0.2010 |
| 22:5n-3 (DPA) | 9.69 ± 3.36 | 5.48 ± 1.75 | 4.42 ± 1.25 | 0.0000 |
| 22:0 | 0.48 ± 0.27 | 0.34 ± 0.17 | 0.36 ± 0.43 | 0.3908 |
| 22:1 | 0.68 ± 0.31 | 0.52 ± 0.14 | 0.43 ± 0.21 | 0.0218 |
| 23:1 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.02 ± 0.05 | 0.2268 |
| 23:0 | 0.78 ± 0.18 | 0.52 ± 0.16 | 0.41 ± 0.09 | 0.0000 |
| 24:1 | 1.37 ± 0.33 | 0.66 ± 0.16 | 0.66 ± 0.24 | 0.0000 |
| 24:0 | 0.98 ± 0.29 | 0.61 ± 0.17 | 0.53 ± 0.25 | 0.0004 |
| Total FA | 1384.60 ± 766.56 | 904.11 ± 342.20 | 698.78 ± 307.88 | 0.0056 |
| EPA + DHA | 15.28 ± 5.12 | 6.11 ± 2.88 | 3.64 ± 1.49 | 0.0000 |
| EPA + DHA + DPA | 24.97 ± 8.27 | 11.58 ± 4.54 | 8.05 ± 2.56 | 0.0000 |
| ∑SFA | 579.40 ± 333.52 | 381.87 ± 149.39 | 295.27 ± 131.95 | 0.0079 |
| ∑MUFA | 630.23 ± 393.61 | 425.92 ± 185.28 | 330.79 ± 161.51 | 0.0172 |
| ∑PUFA | 175.14 ± 47.59 | 96.53 ± 28.59 | 72.90 ± 19.03 | 0.0000 |
| PUFA/SFA | 0.36 ± 0.14 | 0.29 ± 0.12 | 0.27 ± 0.10 | 0.1210 |
| ∑n-3PUFA | 41.36 ± 12.83 | 20.81 ± 6.43 | 15.38 ± 5.24 | 0.0000 |
| ∑n-6PUFA | 133.78 ± 35.37 | 75.71 ± 22.91 | 57.51 ± 14.31 | 0.0000 |
| n-6/n-3PUFA | 3.33 ± 0.52 | 3.72 ± 0.63 | 3.87 ± 0.65 | 0.0499 |
| Fatty Acid | Omega-3 | Control | MSM Whole Grain | p-Value |
|---|---|---|---|---|
| 13:0 | 0.24 ± 0.26 | 0.16 ± 0.26 | 0.36 ± 0.50 | 0.3890 |
| 14:1 | 0.84 ± 0.92 | 1.45 ± 1.21 | 0.55 ± 0.86 | 0.6411 |
| 14:0 | 26.95 ± 8.36 | 31.22 ± 8.74 | 40.09 ± 13.89 | 0.0190 |
| 15:1 | 19.20 ± 6.55 | 15.49 ± 5.47 | 18.81 ± 7.39 | 0.7370 |
| 16:0 | 755.15 ± 151.51 | 765.95 ± 134.32 | 906.84 ± 205.56 | 0.0938 |
| 16:1 | 66.47 ± 18.67 | 83.97 ± 26.54 | 89.50 ± 28.94 | 0.0783 |
| 17:1n8c + a17:0 | 46.43 ± 11.84 | 62.51 ± 16.78 | 78.31 ± 25.78 | 0.0017 |
| 17:0 | 73.54 ± 16.08 | 76.10 ± 16.51 | 101.28 ± 26.67 | 0.0143 |
| 18:3n6 | 9.67 ± 3.17 | 13.31 ± 4.89 | 15.45 ± 5.60 | 0.0160 |
| 18:4n-3 | 6.10 ± 3.34 | 2.34 ± 2.22 | 2.15 ± 3.18 | 0.0098 |
| 18:2n-6 (LA) | 508.28 ± 68.59 | 438.04 ± 84.82 | 570.01 ± 99.23 | 0.2330 |
| 18:3n-3 (ALA) | 72.78 ± 18.68 | 50.46 ± 9.52 | 58.65 ± 12.48 | 0.0655 |
| CLA | 12.45 ± 2.57 | 15.13 ± 3.13 | 18.05 ± 8.47 | 0.0346 |
| 18:0 | 1050.30 ± 82.46 | 879.11 ± 139.95 | 1058.24 ± 190.38 | 0.9770 |
| 18:1 | 1414.42 ± 210.44 | 1414.52 ± 268.81 | 1521.50 ± 345.97 | 0.4875 |
| 19:1 | 7.25 ± 1.88 | 8.93 ± 2.98 | 12.35 ± 3.47 | 0.0010 |
| 20:4n-6 (ARA) | 213.71 ± 41.72 | 310.53 ± 69.00 | 368.35 ± 71.25 | 0.0000 |
| 20:5n-3 (EPA) | 122.60 ± 36.37 | 40.03 ± 9.65 | 43.99 ± 10.29 | 0.0000 |
| 22:3 | 2.08 ± 2.23 | 6.35 ± 2.14 | 7.16 ± 1.72 | 0.0001 |
| 20:3n-6 | 58.65 ± 9.41 | 33.88 ± 5.74 | 49.26 ± 16.35 | 0.1700 |
| 20:4n-3 | 6.07 ± 0.84 | 5.00 ± 2.59 | 6.03 ± 1.75 | 0.9550 |
| 20:2n-6 | 12.96 ± 3.02 | 6.72 ± 1.79 | 8.59 ± 1.24 | 0.0033 |
| 20:0 | 33.42 ± 7.26 | 19.28 ± 5.06 | 21.45 ± 3.60 | 0.0652 |
| 20:1 | 5.02 ± 0.28 | 6.25 ± 1.61 | 6.38 ± 1.97 | 0.0007 |
| 21:5n-3 | 1.58 ± 0.71 | 3.27 ± 2.79 | 2.76 ± 2.20 | 0.2770 |
| 21:0 | 0.37 ± 0.21 | 0.28 ± 0.29 | 0.51 ± 0.26 | 0.2643 |
| 22:5n-6 | 5.23 ± 3.89 | 17.08 ± 3.63 | 21.53 ± 5.93 | 0.0000 |
| 22:6n-3 (DHA) | 286.77 ± 95.79 | 116.09 ± 30.92 | 124.56 ± 30.70 | 0.0001 |
| 22:4n-6 | 16.44 ± 9.99 | 39.84 ± 11.41 | 54.47 ± 11.01 | 0.0000 |
| 22:5n-3 (DPA) | 185.12 ± 30.84 | 133.09 ± 29.99 | 154.08 ± 24.89 | 0.0515 |
| 22:0 | 6.78 ± 1.87 | 2.05 ± 0.87 | 2.12 ± 0.74 | 0.0000 |
| 22:1 | 9.76 ± 0.84 | 8.20 ± 1.11 | 9.31 ± 1.81 | 0.5057 |
| 23:1 | 0.56 ± 0.57 | 0.14 ± 0.22 | 0.17 ± 0.27 | 0.0504 |
| 23:0 | 15.48 ± 0.82 | 16.69 ± 2.77 | 20.36 ± 4.07 | 0.0011 |
| 24:1 | 17.44 ± 2.66 | 11.81 ± 2.62 | 11.14 ± 1.71 | 0.0000 |
| 24:0 | 16.10 ± 1.00 | 15.49 ± 2.52 | 17.48 ± 2.82 | 0.2397 |
| Total FA | 5085.02 ± 632.20 | 4650.60 ± 806.74 | 5421.65 ± 1044.59 | 0.5016 |
| EPA + DHA | 409.37 ± 127.95 | 156.12 ± 37.43 | 168.54 ± 35.69 | 0.0000 |
| EPA + DHA + DPA | 594.49 ± 153.14 | 289.22 ± 63.49 | 322.62 ± 56.93 | 0.0002 |
| ∑SFA | 1972.10 ± 229.59 | 1814.93 ± 295.51 | 2179.67 ± 431.38 | 0.2769 |
| ∑MUFA | 1593.04 ± 242.26 | 1604.51 ± 317.62 | 1736.91 ± 404.01 | 0.4181 |
| ∑PUFA | 1519.88 ± 191.39 | 1231.16 ± 228.83 | 1505.07 ± 261.64 | 0.8519 |
| PUFA/SFA | 0.77 ± 0.06 | 0.68 ± 0.06 | 0.70 ± 0.07 | 0.0365 |
| ∑n-3PUFA | 682.48 ± 170.17 | 356.63 ± 72.64 | 399.37 ± 68.16 | 0.0004 |
| ∑n-6PUFA | 1045.88 ± 117.45 | 1167.97 ± 233.56 | 1452.52 ± 275.64 | 0.0008 |
| n-6/n-3PUFA | 1.69 ± 0.83 | 3.30 ± 0.43 | 3.66 ± 0.49 | 0.0000 |
| Fatty Acid | Omega-3 | Control | MSM Whole Grain | p-Value |
|---|---|---|---|---|
| 13:0 | 0.22 ± 0.21 | 0.16 ± 0.22 | 0.28 ± 0.22 | 0.5721 |
| 14:1 | 0.1 ± 0.14 | 0.05 ± 0.15 | 0.04 ± 0.07 | 0.2296 |
| 14:0 | 7 ± 2.29 | 6.96 ± 2.3 | 5.42 ± 1.27 | 0.0886 |
| 15:1 | 5.33 ± 1.39 | 5.73 ± 1.45 | 3.72 ± 1.33 | 0.0217 |
| 16:0 | 317.14 ± 52.86 | 339.32 ± 63.53 | 283.75 ± 48.67 | 0.2055 |
| 16:1 | 10.57 ± 2.87 | 11.21 ± 2.56 | 11.39 ± 2.94 | 0.5105 |
| 17:1n8c + a17:0 | 12.44 ± 1.91 | 14.29 ± 3.33 | 12.04 ± 2.77 | 0.7594 |
| 17:0 | 25.4 ± 3.66 | 28.7 ± 7.39 | 23.32 ± 5.09 | 0.4353 |
| 18:3n6 | 1.75 ± 0.54 | 1.28 ± 0.28 | 0.98 ± 0.25 | 0.0001 |
| 18:4n-3 | 0.00 ± 0.00 | 0.04 ± 0.12 | 0.01 ± 0.03 | 0.7995 |
| 18:2n-6 (LA) | 281.42 ± 66.94 | 280.63 ± 73.32 | 249.57 ± 56.92 | 0.2849 |
| 18:3n-3 (ALA) | 12.43 ± 3.02 | 8.4 ± 1.62 | 7.63 ± 1.61 | 0.0001 |
| CLA | 3.5 ± 0.82 | 3.78 ± 0.85 | 3.11 ± 1.14 | 0.3653 |
| 18:0 | 351.69 ± 51.59 | 327.73 ± 62.89 | 279.14 ± 47.83 | 0.0054 |
| 18:1 | 320.27 ± 53.92 | 314.51 ± 56.7 | 293.23 ± 58.49 | 0.2854 |
| 19:1 | 2.17 ± 0.44 | 3.84 ± 1.28 | 2.9 ± 1.01 | 0.1645 |
| 20:4n-6 (ARA) | 169.81 ± 27.05 | 246.86 ± 57.97 | 209.05 ± 37.93 | 0.0940 |
| 20:5n-3 (EPA) | 69.1 ± 17.67 | 16.87 ± 4.66 | 16.57 ± 2.71 | 0.0000 |
| 20:3 | 1.1 ± 0.62 | 2.98 ± 0.75 | 3.06 ± 1.12 | 0.0001 |
| 20:3n-6 | 19.6 ± 3.26 | 17.41 ± 5.3 | 12.88 ± 3.59 | 0.0010 |
| 20:4n-3 | 1.95 ± 0.45 | 1.91 ± 0.92 | 2.54 ± 1.71 | 0.2627 |
| 20:2n-6 | 7.9 ± 1.65 | 8.77 ± 2.68 | 6.62 ± 2.3 | 0.2320 |
| 20:0 | 5.35 ± 1.12 | 5.84 ± 1.13 | 4.91 ± 0.85 | 0.3655 |
| 20:1 | 11.88 ± 2.14 | 9.14 ± 2.2 | 8.52 ± 2.19 | 0.0020 |
| 21:5n-3 | 0.48 ± 0.13 | 0.99 ± 0.38 | 1.04 ± 0.35 | 0.0006 |
| 21:0 | 0.69 ± 0.13 | 0.82 ± 0.16 | 0.65 ± 0.13 | 0.5464 |
| 22:5n-6 | 0.44 ± 0.49 | 3.03 ± 0.73 | 2.33 ± 0.62 | 0.0003 |
| 22:6n-3 (DHA) | 58.76 ± 10.76 | 25.17 ± 5.59 | 25.3 ± 5.21 | 0.0000 |
| 22:4n-6 | 3.52 ± 0.76 | 15.04 ± 5.17 | 9.88 ± 3.32 | 0.0133 |
| 22:5n-3 (DPA) | 43.39 ± 6.14 | 33.76 ± 8.04 | 29.14 ± 4.96 | 0.0000 |
| 22:0 | 35.02 ± 6.6 | 36.52 ± 8.02 | 29.06 ± 5.3 | 0.0000 |
| 22:1 | 9.01 ± 1.71 | 5.12 ± 2.16 | 3.62 ± 1.08 | 0.0659 |
| 23:1 | 0.56 ± 0.23 | 0.96 ± 0.34 | 0.8 ± 0.28 | 0.1012 |
| 23:0 | 8.84 ± 1.35 | 9.96 ± 1.97 | 7.94 ± 1.71 | 0.2826 |
| 24:1 | 30.55 ± 5.54 | 33.12 ± 7.47 | 32.73 ± 7.26 | 0.4763 |
| 24:0 | 34.42 ± 5.47 | 37.53 ± 7.62 | 31.21 ± 6.79 | 0.3120 |
| Total FA | 1499.2 ± 208.97 | 1515.42 ± 308.98 | 1322.14 ± 193.73 | 0.1155 |
| EPA + DHA | 127.86 ± 26.2 | 42.04 ± 9.14 | 41.87 ± 7.28 | 0.0000 |
| EPA + DHA + DPA | 171.26 ± 29.76 | 75.8 ± 15.82 | 71.01 ± 10.98 | 0.0000 |
| ∑SFA | 439.41 ± 67.09 | 471.55 ± 89.38 | 390.25 ± 66.92 | 0.1743 |
| ∑MUFA | 385.11 ± 61.94 | 377.95 ± 69.57 | 353.22 ± 71.1 | 0.2933 |
| ∑PUFA | 674.69 ± 91.83 | 665.92 ± 156.17 | 578.66 ± 79.86 | 0.0698 |
| PUFA/SFA | 1.54 ± 0.13 | 1.4 ± 0.12 | 1.51 ± 0.29 | 0.7367 |
| ∑n-3PUFA | 187.22 ± 32.15 | 90.12 ± 18.16 | 85.28 ± 13.7 | 0.0000 |
| ∑n-6PUFA | 318.13 ± 70.45 | 329.97 ± 86.61 | 285.38 ± 60.57 | 0.3264 |
| n-6/n-3PUFA | 1.74 ± 0.42 | 3.65 ± 0.64 | 3.44 ± 1.14 | 0.0003 |
| Fatty Acid | Omega-3 | Control | MSM Whole Grain | p-Value |
|---|---|---|---|---|
| 13:0 | 0.14 ± 0.35 | 0.02 ± 0.07 | 0.03 ± 0.1 | 0.2533 |
| 14:1 | 0.51 ± 0.35 | 0.61 ± 0.3 | 0.36 ± 0.47 | 0.3935 |
| 14:0 | 45.97 ± 95.07 | 13.63 ± 9.65 | 23.69 ± 32.81 | 0.3994 |
| 15:1 | 11.42 ± 20.53 | 4.75 ± 2.84 | 6.68 ± 5.79 | 0.3391 |
| 16:0 | 539.22 ± 573.68 | 402.04 ± 132.98 | 389.7 ± 233.75 | 0.3616 |
| 16:1 | 59.06 ± 91.19 | 34.61 ± 18.23 | 41.34 ± 35.23 | 0.4919 |
| 17:1n8c + a17:0 | 33.28 ± 40.65 | 21.73 ± 10.42 | 26.43 ± 18.68 | 0.5655 |
| 17:0 | 57.87 ± 82.96 | 37.03 ± 17.28 | 41.53 ± 29.75 | 0.4822 |
| 18:3n6 | 2.81 ± 1.01 | 2.55 ± 0.55 | 2.24 ± 0.57 | 0.0919 |
| 18:4n-3 | 0.31 ± 0.46 | 0.15 ± 0.29 | 0.06 ± 0.12 | 0.0823 |
| 18:2n-6 (LA) | 543.33 ± 141.44 | 591.85 ± 100.17 | 466.72 ± 122.48 | 0.1901 |
| 18:3n-3 (ALA) | 31.32 ± 26.24 | 18.55 ± 7.28 | 22.03 ± 15.47 | 0.2636 |
| CLA | 709.39 ± 866.87 | 492.75 ± 233.75 | 459.86 ± 332.73 | 0.3144 |
| 18:0 | 862.44 ± 1163.49 | 557.84 ± 334.03 | 601.47 ± 462.38 | 0.4367 |
| 18:1 | 9.67 ± 10.06 | 6.03 ± 2.24 | 5.68 ± 3 | 0.1578 |
| 19:1 | 3.32 ± 3.49 | 2.59 ± 1.28 | 3.41 ± 2.54 | 0.9383 |
| 20:4n-6 (ARA) | 128.74 ± 35.44 | 166.3 ± 33.44 | 143.74 ± 48.1 | 0.4257 |
| 20:5n-3 (EPA) | 38.59 ± 11.44 | 18.01 ± 3.9 | 14.87 ± 4.96 | 0.0000 |
| 22:3 | 1.6 ± 0.83 | 3.05 ± 0.79 | 2.23 ± 0.62 | 0.1398 |
| 20:3n-6 | 18.01 ± 4.67 | 12.78 ± 3.01 | 9.71 ± 2.57 | 0.0000 |
| 20:4n-3 | 0.22 ± 0.28 | 0.48 ± 0.72 | 0.11 ± 0.24 | 0.6342 |
| 20:2n-6 | 5.12 ± 2.19 | 2.45 ± 0.87 | 1.82 ± 0.6 | 0.0000 |
| 20:0 | 6.11 ± 8.08 | 4.76 ± 2.14 | 4.27 ± 2.66 | 0.4158 |
| 20:1 | 11.82 ± 10.49 | 6.45 ± 2.36 | 5.24 ± 3.19 | 0.0301 |
| 21:5n-3 | 0.29 ± 0.55 | 0.62 ± 0.32 | 0.39 ± 0.26 | 0.6112 |
| 21:0 | 0.62 ± 0.74 | 0.48 ± 0.13 | 0.61 ± 0.46 | 0.9544 |
| 22:5n-6 | 1.26 ± 0.58 | 2.49 ± 0.57 | 2.21 ± 0.64 | 0.0045 |
| 22:6n-3 (DHA) | 36.74 ± 9.42 | 15.37 ± 4.1 | 11.99 ± 3.73 | 0.0000 |
| 22:4n-6 | 2.91 ± 0.89 | 6.06 ± 1.29 | 5.5 ± 1.16 | 0.0003 |
| 22:5n-3 (DPA) | 34.82 ± 9.38 | 19.03 ± 14.44 | 23 ± 6.92 | 0.0301 |
| 22:0 | 2.95 ± 1.25 | 0.97 ± 0.33 | 0.69 ± 0.38 | 0.0000 |
| 22:1 | 4.47 ± 2.07 | 5.03 ± 1.97 | 3.72 ± 1.83 | 0.4011 |
| 23:1 | 0.43 ± 0.28 | 0.17 ± 0.19 | 0.02 ± 0.05 | 0.0000 |
| 23:0 | 4.58 ± 1.46 | 6.38 ± 1.24 | 4.43 ± 1.42 | 0.8409 |
| 24:1 | 10.03 ± 3.11 | 8.43 ± 1.31 | 5.59 ± 1.55 | 0.0000 |
| 24:0 | 4.19 ± 1.24 | 4.65 ± 1.82 | 3.72 ± 1.03 | 0.4712 |
| Total FA | 3223.54 ± 3074.77 | 2470.68 ± 880 | 2335.08 ± 1176.42 | 0.3140 |
| EPA + DHA | 75.32 ± 20.32 | 33.38 ± 7.73 | 26.86 ± 8.05 | 0.0000 |
| EPA + DHA + DPA | 110.14 ± 28.19 | 52.41 ± 18.48 | 49.86 ± 14.91 | 0.0000 |
| ∑SFA | 1384 ± 1645.79 | 971.52 ± 396.97 | 938.25 ± 632.49 | 0.3409 |
| ∑MUFA | 983.83 ± 1309.31 | 633.39 ± 366.3 | 684.53 ± 519.99 | 0.4274 |
| ∑PUFA | 855.71 ± 225.88 | 865.77 ± 149 | 712.3 ± 183.06 | 0.1016 |
| PUFA/SFA | 1 ± 0.42 | 0.98 ± 0.25 | 0.99 ± 0.48 | 0.9636 |
| ∑n-3PUFA | 143.88 ± 41.03 | 75.26 ± 23.68 | 74.68 ± 23.06 | 0.0000 |
| ∑n-6PUFA | 711.84 ± 189.55 | 790.5 ± 130.94 | 637.62 ± 168.09 | 0.3408 |
| n-6/n-3PUFA | 5.01 ± 0.68 | 11.12 ± 2.43 | 8.82 ± 1.92 | 0.0041 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pewan, S.B.; Otto, J.R.; Kinobe, R.T.; Adegboye, O.A.; Malau-Aduli, A.E.O. Nutritional Enhancement of Health Beneficial Omega-3 Long-Chain Polyunsaturated Fatty Acids in the Muscle, Liver, Kidney, and Heart of Tattykeel Australian White MARGRA Lambs Fed Pellets Fortified with Omega-3 Oil in a Feedlot System. Biology 2021, 10, 912. https://doi.org/10.3390/biology10090912
Pewan SB, Otto JR, Kinobe RT, Adegboye OA, Malau-Aduli AEO. Nutritional Enhancement of Health Beneficial Omega-3 Long-Chain Polyunsaturated Fatty Acids in the Muscle, Liver, Kidney, and Heart of Tattykeel Australian White MARGRA Lambs Fed Pellets Fortified with Omega-3 Oil in a Feedlot System. Biology. 2021; 10(9):912. https://doi.org/10.3390/biology10090912
Chicago/Turabian StylePewan, Shedrach Benjamin, John Roger Otto, Robert Tumwesigye Kinobe, Oyelola Abdulwasiu Adegboye, and Aduli Enoch Othniel Malau-Aduli. 2021. "Nutritional Enhancement of Health Beneficial Omega-3 Long-Chain Polyunsaturated Fatty Acids in the Muscle, Liver, Kidney, and Heart of Tattykeel Australian White MARGRA Lambs Fed Pellets Fortified with Omega-3 Oil in a Feedlot System" Biology 10, no. 9: 912. https://doi.org/10.3390/biology10090912
APA StylePewan, S. B., Otto, J. R., Kinobe, R. T., Adegboye, O. A., & Malau-Aduli, A. E. O. (2021). Nutritional Enhancement of Health Beneficial Omega-3 Long-Chain Polyunsaturated Fatty Acids in the Muscle, Liver, Kidney, and Heart of Tattykeel Australian White MARGRA Lambs Fed Pellets Fortified with Omega-3 Oil in a Feedlot System. Biology, 10(9), 912. https://doi.org/10.3390/biology10090912