1. Introduction
The American Academy of Pediatrics evidenced the prophylactic and therapeutic roles of human milk in educating infants’ immune systems and providing protection against many infectious diseases such as gastrointestinal, respiratory, inflammatory bowel, and allergic diseases [
1,
2]. These protective effects of breast milk are due to the orchestrated action of several bioactive molecules, such as oligosaccharides, fatty acids, immunoglobulins, cytokines, immune cells, lactoferrin, immunomodulating factors, and healthy microbial communities [
3].
Breast milk is the second integral source of infant microbes after the birth canal in vaginally born infants [
4]. It has been predicted that an infant takes approximately 10
5–10
7 commensal bacteria every day via consuming 800 mL of breast milk. Human breast milk contributes a distinctive role in the initiation, development, and composition of the neonatal gut microbiota. The human milk microbiome has a diverse array of bacterial species, including beneficial, commensal, and potentially probiotic bacteria [
5].
Probiotics are live bacteria that deliver health benefits to the host when consumed at adequate levels as described by the Food and Agriculture Organization (FAO) and the World Health Organization (WHO) [
6]. Probiotic bacteria have many beneficial characteristics such as the ability to colonize and dominate in the neonatal gut, ability to resist stomach acid and bile salts, adherence to the intestinal mucosa, initiation of anti-inflammatory responses, inhibition of pathogens by the production of antimicrobial constituents, and augmentation of the immune system [
7,
8,
9].
A new era of therapeutics is in perspective in which probiotics and their purified molecules will be employed as a wise, safe alternative to medication and other treatments to control health imbalance and diseases in humans and animals [
10]. In this manner, several studies have reported the role of probiotics in the prevention and treatment of inflammatory bowel disease [
11,
12] food hypersensitivity [
13], cardiometabolic disorders [
14], and antitumor activity [
15].
Previous studies have evidenced the richness of human milk and feces samples with promising novel probiotics. The investigation by Lee et al. [
16] reported the presence of novel
Lactobacillus gasseri EJL and
Bifidobacterium breve JTL strains in the milk and feces samples of Korean Mother-infant pairs. A recent interesting study by Li et al. [
17] identified novel LAB bacterial strains belonging to
Lactobacillus gasseri,
Lactiplantibacillus plantarum, and
Lacticaseibacillus rhamnosus from Chinese infants with potential probiotic characteristics against inflammation and oxidative stress-related human diseases.
The microbial diversity of human milk and consequently, infants’ gut is controlled by environmental, demographic and genetic factors, and the maternal lifestyle. Bioprospecting the gut microbiota, and selecting promising probiotic candidates, is of great importance to the new insights of personalized medicine. In the current investigation into future perspectives in treating chronic diseases, we prospect for such healthy probiotics from Egyptian populations characterized by unique immune systems. Targeted bacterial isolates were profiled in terms of morphological, biochemical, and ultrastructural properties. Probiotic tests, antibiotic susceptibility, and antibacterial activity were also considered. The taxonomic evidence of these isolates was demonstrated based on 16S rRNA gene sequence and phylogenetic tree analysis.
4. Discussion
In this investigation, we seek new potent probiotics from the breast milk of Egyptian mothers and stool samples of their infants. The novel aspects of our study include (1) exploring human milk microbiota diversity from Egyptian samples for the first time and (2) isolating innovative probiotics from Egyptian infants’ feces that are characterized by a unique immune system as prophylactic and therapeutic agents for controlling chronic diseases.
Where lactic acid bacteria colonize gut epithelial cells and withstand pathogens and reactive oxygen species (ROS) associated with gut diseases, it should have the ability to endure harsh conditions in the human body (intestinal juice, low pH, and salivary enzymes) to maintain gut microbiota balance, immune homeostasis and monitor beneficial physiological roles in human health [
33,
34]. In this manner and based on morphological and physiological characteristics, we selected only gram-positive, catalase-negative, positive microaerophilic and non-endospore forming lactic acid bacterial isolates to be investigated in the current study. Rod isolates revealed a positive ability to grow at 4.0% bile salt, pH 6.8, pH 9.6, 6.5% NaCl, at 45 °C with positive abilities to hydrolyze arginine and coagulate milk in accordance with the investigations of Soni et al. [
35] and Lackey et al. [
36]. All isolates exhibited high abilities to ferment various carbohydrates and produce acid from lactose, galactose, glucose, fructose, maltose, sucrose, mannose, rhamnose, arabinose, and melibiose. However, these isolates displayed a negative ability to produce acid from ribose, mannitol, ribose, salicin, and sorbitol. These results agreed with the recent report of Li et al. [
17] who isolated 27 gram-positive and catalase-negative strains from healthy infant feces and evidenced their negative profile to mannitol and sorbitol. Interestingly, unlike all isolates, ASO57, ASO55, ASO5, and ASO100 showed negative profiles for producing acid from xylose, raffinose, and trehalose reflecting unique profiles and possible different probiotic characteristics.
Pairwise sequence alignment of 16S rRNA sequences revealed the presence of three genera,
Lactobacillus,
Enterococcus, and
Lactococcus.
Lactobacillus was the most common in human milk and feces samples with a high incidence of its different species (
L. paracasei,
L. delbrueckii,
L. plantarum,
L. gasseri and
L. casei); these results were matched with the previous study of Zhang et al. [
37] who reported a high incidence of
Lactobacillus strains in feces samples of Chinese babies. Interestingly, six of our isolates evidenced low identity ratios ranging from 81.39% to 94.63% (less than 95%) with database sequences. The isolate that revealed the lowest identity ratio (
L. delbrueckii, ASO 100) expressed the highest antibiotic resistance, antibacterial and probiotic activity. This isolate could be a new species as reported by Thompson et al. [
38] and Badr et al. [
39], and strains from different microbial species share less than 95% Average Nucleotide Identity (ANI). The low identity ratio of this isolate specifically was evidenced by chemical, physiological and probiotic features. Hence, this isolate and probably the other five isolates that shared less than 95% Average Nucleotide Identity could be a new probiotic species with novel and unique characteristics. These isolates showed the nearest similarity to
E. faecium and
L. delbrueckii strains. The previous investigation by Evivie et al. [
40] confirmed the medicinal usage of
L. delbrueckii isolates as a probiotic against foodborne pathogens. Moreover, recent studies have evidenced the role of
E. faecium as a promising probiotic candidate for both human and animal use [
41,
42,
43,
44]. Despite 16S rRNA being a very conserved region, six of our isolates revealed huge divergence in that region and this could be explained by the effect of environmental, demographic, genetic factors, and the maternal lifestyle on modifying the microbial diversity of human milk and infants’ gut [
45].
Previous investigations evidenced the role of current isolated strains as a promising candidate probiotic for medicinal and industrial usage for humans and animals. An interesting study by Salaris et al. [
46] revealed that
L. paracasei is a promising candidate probiotic that exhibits prophylactic potential effect against SARS-CoV-2 infection. Moreover, Otaka et al. [
47] indicated that
L. paracasei was useful to alleviate depressive symptoms, partly through its association with an abundance of actinobacteria in the gut microbiota. Another investigation by Guerra et al. [
48] showed that lactobacilli isolates from newborn stools exhibited different probiotic properties such as gastrointestinal tolerance, antibiotic susceptibility, inhibition of pathogen biofilm formation, absence of alfa or gamma-blood hemolysis, and lysozyme sensibility. Besides, Hill et al.’s investigation [
49] showed that the
L. casei clusters have the potential to be used prophylactically or therapeutically in diseases related to a disturbance to the gut microbiota. Probiotics like
L. plantarum are beneficial bacteria that stimulate the digestive system, fight pathogenic microbes, and help the human body to produce vitamins. Many people take
L. plantarum probiotic pills to heal or prevent complaints, including seasonal allergies and irritable bowel syndrome [
50].
For testing the competing probiotic characteristics of our isolates, we selected nine representative isolates from different genera for further experiments based on literature evidence of their probiotic characteristics and the uniqueness of their biochemical, physiological, and 16S rRNA molecular profiles. We utilized scanning electron microscopy investigation for deep visualization of morphological ultrastructure features of these isolates confirming clear differences in cell shapes and assemblies of all investigated isolates. From the selected isolates there were three cocci isolates (E. faecalis ASO44, E. faecium ASO292, L. lactis ASO26) and six bacilli isolates (L. delbrueckii ASO100, L. plantarum ASO50, L. casei ASO53, L. rhamnosus ASO20, L. gasseri ASO25, L. paracasei ASO32). All isolates revealed aggregation ability confirming their ability to colonize gut epithelial cells.
Owing to market competition and demand, probiotics must be able to endure challenging environments including the acidic environment of the gastrointestinal tract (GIT), intestinal bile salts, and digestive enzymes. Most exogenous microbes die when ingested into the GIT because of the very low pH of the secreted gastric juice (pH of 2.0). It is expected that probiotic strains should be able to adapt and tolerate the acidic nature of the GIT as they pass by to colonize the gut of their host [
51,
52]. Moreover, pH tolerance is important for the improvement of fermented foods like yogurt and cheese that affect strain sustainability due to their high acidity. Interestingly, all the investigated isolates in this study revealed acid tolerance of the isolated LAB to the pH value from 6 to 3 for 4 h after incubation. LAB are known for their capability to tolerate acidic pH [
53]. Our result resembles preceding studies that reported the survival of LAB strains against simulated gastric juice with a pH of 2.0 [
54,
55].
One of the major requirements for probiotic selection is the ability to survive and grow in the GIT, so they should be able to tolerate the intestinal bile salt. Probiotic physiological alterations, including exopolysaccharide synthesis and carbohydrate fermentation, are related to the resistance to elevated bile salts [
56]. The adaptation of probiotics to bile salts is also connected to the structure of membrane proteins and fatty acids as well as the prevention of pathogen adherence to human mucus [
57,
58]. To compete with pathogens when employed in functional foods, probiotic strains must possess resistance to bile salts. Tolerance of an average level of 0.3% of the bile salt has been estimated in many studies for potential probiotic LAB candidates [
59]. Interestingly, our results estimated that, except for the
L. lactis ASO 26, all the LAB isolates in this study exhibited tolerance to 0.5% bile salts for a 4 h incubation period.
Regarding antibiotic resistance in probiotics, it is considered a safety concern, as antibiotic resistance encoding genes could transfer among the microorganism community of the gut. The genomic context of the antibiotic resistance determinants of the current study’s probiotic strains is unknown but a future follow-up study will be performed to go through whole genome sequencing of these isolates to ensure that the antibiotic resistance determinants are not present as part of mobile genetic elements. Previous evidence by [
60] reported the lack of cytochrome-mediated electron transport in Lactobacillus genera, and the presence of D-Ala-D-lactate in their peptidoglycan, hence, their resistance to different antibiotics including streptomycin, and vancomycin is considered to be intrinsic. Consequently, LAB probiotic strains can be used safely as pills alongside or after antibiotic treatment to restore the gut microbiota homeostasis [
61]. In this case, antibiotic resistance possesses a strong advantage in order for probiotics to survive under antibiotic treatment conditions. In the current study, all studied isolates were resistant to kanamycin, tetracycline, neomycin, streptomycin, and vancomycin except,
L. lactis ASO26, which was sensitive to streptomycin and vancomycin. The literature evidenced that most LAB species are resistant to kanamycin [
62,
63]. Remarkably, all isolates were resistant to streptomycin and vancomycin, in harmony with previous reports [
64,
65].
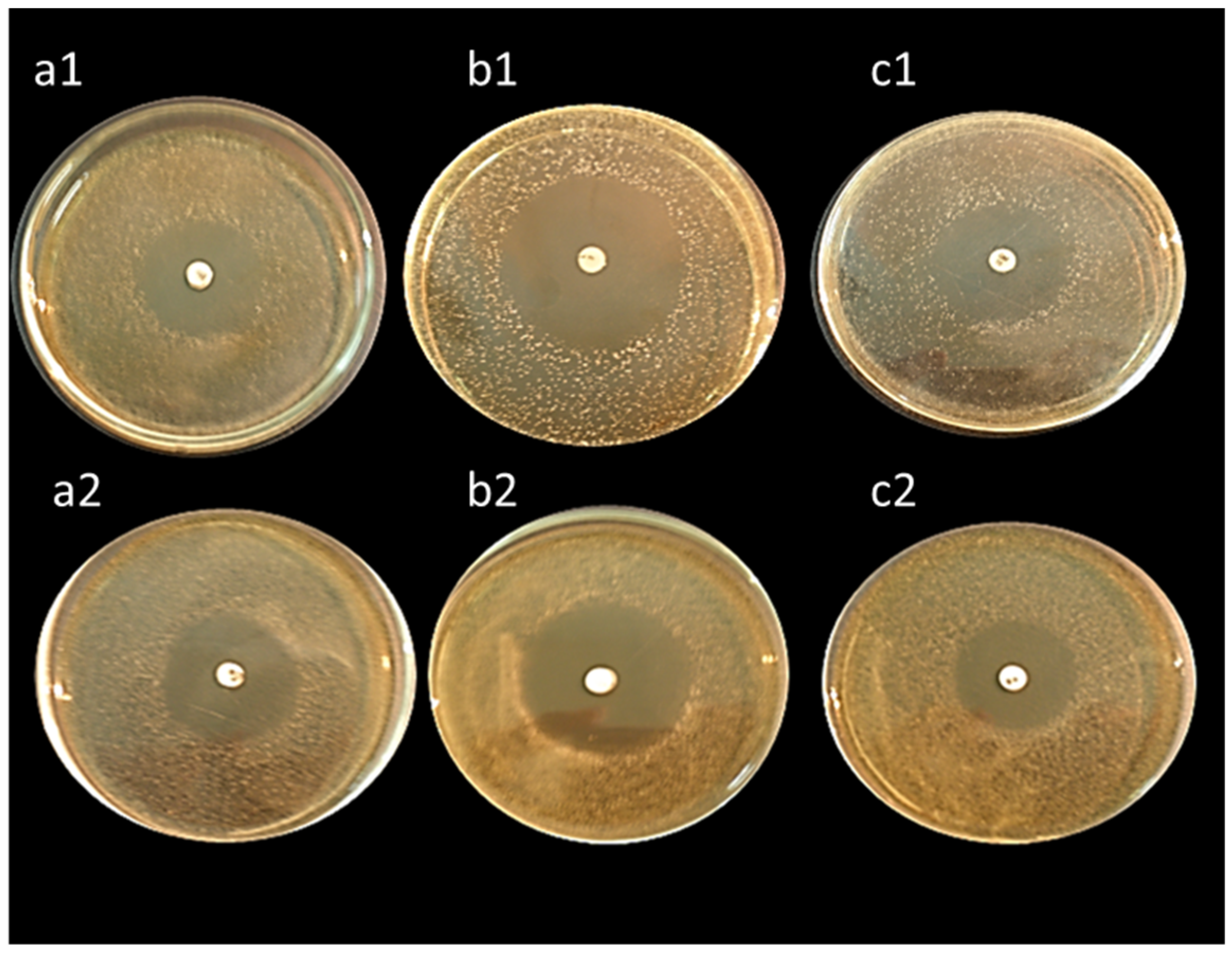
All tested isolates exhibited high antibacterial activity against
Bacillus subtilis,
Staphylococcus aureus, and
Escherichia coli except,
L. lactis ASO26 which did not reveal any antibacterial activity. Our results were in coincidence with Klayraung et al. [
66], who investigated the antibacterial activity of lactobacilli isolated from four kinds of traditional fermented foods on
Staphylococcus aureus,
Salmonella typhi, and
Escherichia coli, reporting the high antibacterial potency of LAB against
S. aureus,
S. typhi, and
E. coli. The antibacterial activity of lactic acid bacteria could be explained by their production of a wide variety of different inhibitory substances that prolong the time scale of preservation of the fermented products. The preservative action of LAB in foods results from the formation of metabolites with antimicrobial activity, e.g., organic acids (lactic, acetic, formic, etc.), hydrogen peroxide (in the presence of oxygen), diacetyl, aldehydes (e.g., β-hydroxy-propionaldehyde) and bacteriocins or bactericidal proteins during lactic fermentation, which make them useful in food bio-preservation.
As the newly identified LAB isolates exhibited high acid and bile salt tolerance, antibiotic resistance, and antibacterial activity, they could be applied as effective and competing probiotic pills for modulating intestinal pathogens and human diseases. Interestingly, pairwise sequence alignment results evidenced a low identity ratio of six isolates (less than 95%) with a high probability to be new species. Further research will be assessed to go through whole genomic sequencing of these isolates, especially the isolate L. delbrueckii, ASO 100 that will be subject to complete proteomic analysis to stand for its probiotic determinants, as it revealed the most brilliant probiotic and antibacterial features, along with another in vivo experiment that will be conducted to test the prophylactic and therapeutic ability of this isolate to modulate gut–brain axis microbiota in an Alzheimer’s disease animal model.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}