
Latitude-Induced and Behaviorally Thermoregulated Variations in Upper Thermal Tolerance of Two Anuran Species



Abstract
:Simple Summary
Abstract
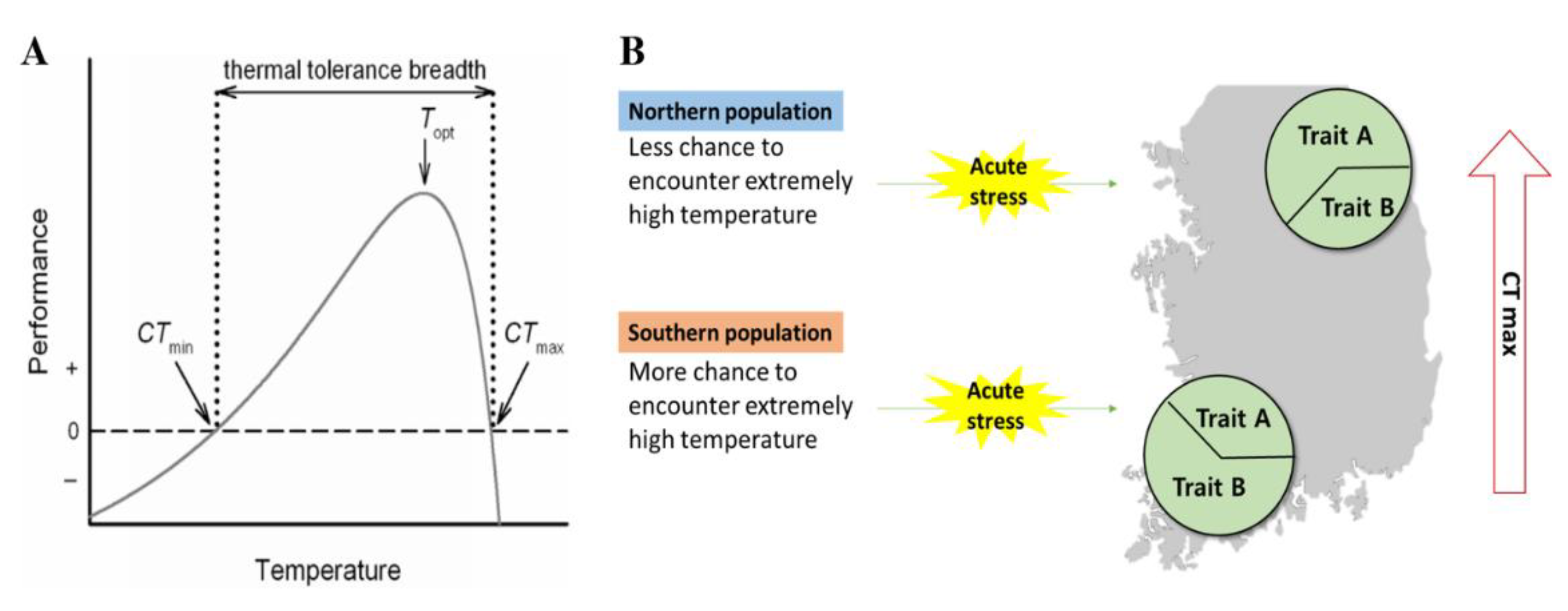
1. Introduction
2. Materials and Methods
2.1. Species Introduction
2.2. Tadpole Collection
2.3. Estimation of Critical Thermal Maximum
2.4. Behavioral Measurements
2.5. Statistical Analysis
3. Results
3.1. Effect of Abiotic Factors on Critical Thermal Maximum
3.2. Temperature-Induced Behavior Changes in the Two Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seebacher, F.; Franklin, C.E. Determining environmental causes of biological effects: The need for a mechanistic physiological dimension in conservation biology. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1607–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, J.M.; Storey, K.B. Environmental Stressors and Gene Responses; Elsevier: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Huey, R.B.; Stevenson, R. Integrating thermal physiology and ecology of ectotherms: A discussion of approaches. Am. Zool. 1979, 19, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Little, A.G.; Seebacher, F. Physiological Performance Curves: When Are They Useful? Front. Physiol. 2021, 12, 805102. [Google Scholar] [CrossRef] [PubMed]
- Rezende, E.L.; Castañeda, L.E.; Santos, M. Tolerance landscapes in thermal ecology. Funct. Ecol. 2014, 28, 799–809. [Google Scholar] [CrossRef]
- Cowles, R.B.; Bogert, C.M. A preliminary study of the thermal requirements of desert reptiles. Bull. Am. Mus. Nat. Hist. 1944, 83, 261–296. [Google Scholar]
- Bennett, J.M.; Calosi, P.; Clusella-Trullas, S.; Martínez, B.; Sunday, J.; Algar, A.C.; Araújo, M.B.; Hawkins, B.A.; Keith, S.; Kühn, I.; et al. GlobTherm, a global database on thermal tolerances for aquatic and terrestrial organisms. Sci. Data 2018, 5, 180022. [Google Scholar] [CrossRef]
- Duarte, H.; Tejedo, M.; Katzenberger, M.; Marangoni, F.; Baldo, D.; Beltrán, J.F.; Martí, D.A.; Richter-Boix, A.; Gonzalez-Voyer, A. Can amphibians take the heat? Vulnerability to climate warming in subtropical and temperate larval amphibian communities. Glob. Chang. Biol. 2012, 18, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Simon, M.N.; Ribeiro, P.L.; Navas, C.A. Upper thermal tolerance plasticity in tropical amphibian species from contrasting habitats: Implications for warming impact prediction. J. Therm. Biol. 2015, 48, 36–44. [Google Scholar] [CrossRef]
- Gutiérrez-Pesquera, L.M.; Tejedo, M.; Olalla-Tárraga, M.; Duarte, H.; Nicieza, A.; Solé, M. Testing the climate variability hypothesis in thermal tolerance limits of tropical and temperate tadpoles. J. Biogeogr. 2016, 43, 1166–1178. [Google Scholar] [CrossRef]
- Terblanche, J.S.; Klok, C.J.; Krafsur, E.S.; Chown, S.L. Phenotypic plasticity and geographic variation in thermal tolerance and water loss of the tsetse Glossina pallidipes (Diptera: Glossinidae): Implications for distribution modelling. Am. J. Trop. Med. Hyg. 2006, 74, 786–794. [Google Scholar] [CrossRef] [Green Version]
- Diamond, S.E.; Nichols, L.M.; McCoy, N.; Hirsch, C.; Pelini, S.L.; Sanders, N.J.; Ellison, A.M.; Gotelli, N.J.; Dunn, R.R. A physiological trait-based approach to predicting the responses of species to experimental climate warming. Ecology 2012, 93, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Enriquez-Urzelai, U.; Kearney, M.; Nicieza, A.; Tingley, R. Integrating mechanistic and correlative niche models to unravel range-limiting processes in a temperate amphibian. Glob. Chang. Biol. 2019, 25, 2633–2647. [Google Scholar] [CrossRef] [PubMed]
- Kellermann, V.; Sgrò, C.M. Evidence for lower plasticity in CTMAX at warmer developmental temperatures. J. Evol. Biol. 2018, 31, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- Bozinovic, F.; Calosi, P.; Spicer, J.I. Physiological correlates of geographic range in animals. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 155–179. [Google Scholar] [CrossRef] [Green Version]
- Richter-Boix, A.; Katzenberger, M.; Duarte, H.; Quintela, M.; Tejedo, M.; Laurila, A. Local divergence of thermal reaction norms among amphibian populations is affected by pond temperature variation. Evolution 2015, 69, 2210–2226. [Google Scholar] [CrossRef]
- Johansson, M.P.; Laurila, A. Maximum thermal tolerance trades off with chronic tolerance of high temperature in contrasting thermal populations of Radix balthica. Ecol. Evol. 2017, 7, 3149–3156. [Google Scholar] [CrossRef]
- Ober, G.T.; Thornber, C.; Grear, J.; Kolbe, J.J. Ecological differences influence the thermal sensitivity of swimming performance in two co-occurring mysid shrimp species with climate change implications. J. Therm. Biol. 2017, 64, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.-J.; Yen, C.-F.; Kam, Y.-C. Metabolic compensation and behavioral thermoregulation of subtropical rhacophorid (Polypedates megacephalus) tadpoles in container habitats. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2007, 146, 101–106. [Google Scholar] [CrossRef]
- Cupp, P.V., Jr. Thermal tolerance of five salientian amphibians during development and metamorphosis. Herpetologica 1980, 36, 234–244. [Google Scholar]
- Tracy, C.R. A model of the dynamic exchanges of water and energy between a terrestrial amphibian and its environment. Ecol. Monogr. 1976, 46, 293–326. [Google Scholar] [CrossRef]
- Haramura, T. Microhabitat selection by tadpoles of Buergeria japonica inhabiting the coastal area. J. Ethol. 2007, 25, 3–7. [Google Scholar] [CrossRef]
- Beiswenger, R.E. Diel patterns of aggregative behavior in tadpoles of Bufo americanus, in relation to light and temperature. Ecology 1977, 58, 98–108. [Google Scholar] [CrossRef]
- Roman, A.; Griffiths, R.A.; Schley, L. Activity patterns and microhabitat selection of Mallorcan midwife toad (Alytes muletensis) tadpoles in natural torrent pools. Amphib. Reptil. 1998, 19, 143–151. [Google Scholar] [CrossRef]
- Beiswenger, R.E. Responses of Bufo tadpoles (Amphibia, Anura, Bufonidae) to laboratory gradients of temperature. J. Herpetol. 1978, 12, 499–504. [Google Scholar] [CrossRef]
- Feder, M.E. The relation of air breathing and locomotion to predation on tadpoles, Rana berlandieri, by turtles. Physiol. Zool. 1983, 56, 522–531. [Google Scholar] [CrossRef]
- Brunelli, E.; Bernabò, I.; Berg, C.; Lundstedt-Enkel, K.; Bonacci, A.; Tripepi, S. Environmentally relevant concen-trations of endosulfan impair development, metamorphosis and behaviour in Bufo bufo tadpoles. Aquat. Toxicol. 2009, 91, 135–142. [Google Scholar] [CrossRef]
- Jeon, J.Y.; Cho, S.; Suk, H.Y.; Lee, C.H.; Song, J.-Y.; Lee, H.; Min, M.-S.; BORZgE, A. Resolving the taxonomic equivocacy and the population genetic structure of Rana uenoi–insights into dispersal and demographic history. Salamandra 2021, 57, 529–540. [Google Scholar]
- Groffen, J.; Andersen, D.; Borzée, A. Breeding phenology and landscape use in all amphibian species from the Republic of Korea based on citizen science data and governmental biodiversity surveys. Front. Environ. Sci. 2022; in press. [Google Scholar] [CrossRef]
- Kwon, S.; Borzée, A.; Koo, K.S.; Seo, J.; Kim, H.; Jang, Y. Shift in spawning date over a five-year period in the climate-sensitive biological indicator species, Rana uenoi. In Proceedings of the Korean Society of Environmental Ecology and Conservation, Online, Korea, 9 September 2020; p. 58. [Google Scholar]
- Othman, S.N.; Litvinchuk, S.N.; Maslova, I.; Dahn, H.; Messenger, K.R.; Andersen, D.; Jowers, M.J.; Kojima, Y.; Skorinov, D.V.; Yasumiba, K.; et al. From Gondwana to the Yellow Sea, evolutionary diversifications of true toads Bufo sp. in the Eastern Palearctic and a revisit of species boundaries for Asian lineages. eLife 2022, 11, e70494. [Google Scholar] [CrossRef]
- Sung, H.-C.; Park, O.-H.; Kim, S.-K.; Park, D.-S.; Park, S.-R. Abundance and breeding migration of the Asian Toad (Bufo gargarizans). J. Ecol. Field Biol. 2007, 30, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.I.; Chuang, M.-F.; Borzée, A. Latitude Induced and Behaviorally Thermoregulated Variations in Upper Thermal Tolerance of Rana uenoi and Bufo sachalinensis, version 1. Mendeley Data 2022. [Google Scholar] [CrossRef]
- Frenne, P.D.; Graae, B.J.; Rodríguez-Sánchez, F.; Kolb, A.; Chabrerie, O.; Decocq, G.; Kort, H.D.; Schrijver, A.D.; Diekmann, M.; Eriksson, O.; et al. Latitudinal gradients as natural laboratories to infer species’ responses to temperature. J. Ecol. 2013, 101, 784–795. [Google Scholar] [CrossRef]
- Frenne, P.D.; Brunet, J.; Shevtsova, A.; Kolb, A.; Graae, B.J.; Chabrerie, O.; Cousins, S.A.; Decocq, G.; Schrijver, A.D.; Diekmann, M.; et al. Temperature effects on forest herbs assessed by warming and transplant experiments along a latitudinal gradient. Glob. Chang. Biol. 2011, 17, 3240–3253. [Google Scholar] [CrossRef]
- Trip, E.D.L.; Clements, K.D.; Raubenheimer, D.; Choat, J.H. Temperature-related variation in growth rate, size, maturation and life span in a marine herbivorous fish over a latitudinal gradient. J. Anim. Ecol. 2014, 83, 866–875. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.S.; Kim, S.S.; Chun, J.H. Pattern of ant diversity in Korea: An empirical test of Rapoport’s altitudinal rule. J. Asia-Pac. Entomol. 2014, 17, 161–167. [Google Scholar] [CrossRef]
- Kim, J.Y.; Shin, M.S.; Seo, C.; Eo, S.H.; Hong, S. Testing the causal mechanism of the peninsular effect in passerine birds from South Korea. PLoS ONE 2021, 16, e0245958. [Google Scholar] [CrossRef] [PubMed]
- Andersen, D.; Litvinchuk, S.N.; Jang, H.J.; Jiang, J.; Koo, K.S.; Maslova, I.; Kim, D.; Jang, Y.; Borzée, A. Incorporation of latitude-adjusted bioclimatic variables increases accuracy in species distribution models. Ecol. Model. 2022, 469, 109986. [Google Scholar] [CrossRef]
- Gosner, K.L. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 1960, 16, 183–190. [Google Scholar]
- Hutchison, V.H.; Maness, J.D. The role of behavior in temperature acclimation and tolerance in ectotherms. Am. Zool. 1979, 19, 367–384. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-S.; Kam, Y.-C. Thermal tolerance and thermoregulation by Taiwanese rhacophorid tadpoles (Buergeria japonica) living in geothermal hot springs and streams. Herpetologica 2005, 61, 35–46. [Google Scholar] [CrossRef]
- Brooks, G.; Sassaman, J.F. Critical thermal maxima of larval and adult Eurycea bislineata. Copeia 1965, 1965, 251–252. [Google Scholar] [CrossRef]
- Spieler, M.; Linsenmair, K.E. Aggregation behaviour of Bufo maculatus tadpoles as an antipredator mechanism. Ethol. Former. Z. Tierpsychol. 1999, 105, 665–686. [Google Scholar] [CrossRef]
- Sillar, K.T.; Robertson, R.M. Thermal activation of escape swimming in post-hatching Xenopus laevis frog larvae. J. Exp. Biol. 2009, 212, 2356–2364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Heisterkamp, S.; van Willigen, B.; Maintainer, R. Package ‘nlme’. Linear and Nonlinear Mixed Effects Models R Package, version 3.1-160; CLAN: Sydney, Australia, 2017. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Barton, K. Package ‘MuMIn’ Multi-Model Inference, version 1.47.1; CLAN: Sydney, Australia, 2019. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Addo-Bediako, A.; Chown, S.L.; Gaston, K.J. Thermal tolerance, climatic variability and latitude. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2000, 267, 739–745. [Google Scholar] [CrossRef] [Green Version]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Global analysis of thermal tolerance and latitude in ectotherms. Proc. R. Soc. B Biol. Sci. 2010, 278, 1823–1830. [Google Scholar] [CrossRef] [Green Version]
- Kern, P.; Cramp, R.L.; Franklin, C.E. Physiological responses of ectotherms to daily temperature variation. J. Exp. Biol. 2015, 218, 3068–3076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in fluctuating thermal environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruel, J.J.; Ayres, M.P. Jensen’s inequality predicts effects of environmental variation. Trends Ecol. Evol. 1999, 14, 361–366. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Huey, R.B. Size, temperature, and fitness: Three rules. Evol. Ecol. Res. 2008, 10, 251–268. [Google Scholar]
- Wilson, R.S.; Franklin, C.E.; James, R.S. Allometric scaling relationships of jumping performance in the striped marsh frog Limnodynastes peronii. J. Exp. Biol. 2000, 203, 1937–1946. [Google Scholar] [CrossRef]
- Ambu, J.; Andersen, D.; Borzée, A. Spawning site selection and segregation at the landscape, habitat and microhabitat scales for five syntopic Asian amphibians. Amphib. Reptil. 2022, 43, 169–180. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Sørensen, J.G.; Loeschcke, V. Adaptation of Drosophila to temperature extremes: Bringing together quantitative and molecular approaches. J. Therm. Biol. 2003, 28, 175–216. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Kristensen, T.N.; Loeschcke, V. The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 2003, 6, 1025–1037. [Google Scholar] [CrossRef]
- Bozinovic, F.; Naya, D.E. Linking Physiology, Climate, and Species Distributional Ranges; Martin, L.B., Ghalambor, C.K., Woods, A., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 277–290. [Google Scholar]
- Brattstrom, B.H. Thermal acclimation in Australian amphibians. Comp. Biochem. Physiol. 1970, 35, 69–103. [Google Scholar] [CrossRef]
- Wassersug, R.J.; Seibert, E.A. Behavioral responses of amphibian larvae to variation in dissolved oxygen. Copeia 1975, 1975, 86–103. [Google Scholar] [CrossRef]
- Angilletta, M.J., Jr.; Angilletta, M.J. Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Kim, S.-H.; Kang, M.-H.; Chang, J.-H. Climate Change Communication in South Korea. In Oxford Research Encyclopedia of Climate Science; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Park, S.U.; Koo, K.A.; Kong, W.-S. Potential impact of climate change on distribution of warm temperate evergreen broad-leaved trees in the Korean Peninsula. J. Korean Geogr. Soc. 2016, 51, 201–217. [Google Scholar]
- Kim, J.-H.; Lee, E.-J.; Yun, J.I. Prediction of blooming dates of spring flowers by using digital temperature forecasts and phenology models. Korean J. Agric. For. Meteorol. 2013, 15, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.-H.; Kim, J.-H.; Oh, K.-H.; Lee, B.-Y. Distributional change and climate condition of warm-temperate evergreen broad-leaved trees in Korea. Korean J. Environ. Ecol. 2011, 25, 47–56. [Google Scholar]
- Rivadeneira, M.M.; Fernández, M. Shifts in southern endpoints of distribution in rocky intertidal species along the south-eastern Pacific coast. J. Biogeogr. 2005, 32, 203–209. [Google Scholar] [CrossRef]
- Sears, M.W.; Raskin, E.; Angilletta, M.J., Jr. The world is not flat: Defining relevant thermal landscapes in the context of climate change. Integr. Comp. Biol. 2011, 51, 666–675. [Google Scholar] [CrossRef]
- Williams, S.E.; Shoo, L.P.; Isaac, J.L.; Hoffmann, A.A.; Langham, G. Towards an integrated framework for assessing the vulnerability of species to climate change. PLoS Biol. 2008, 6, 2621–2626. [Google Scholar] [CrossRef]
- Kearney, M.R.; Isaac, A.P.; Porter, W.P. microclim: Global estimates of hourly microclimate based on long-term monthly climate averages. Sci. Data 2014, 1, 140006. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Site ID | Sample Size | Latitude | Longitude | Alt (m) | Temperature (°C) | Precipitation (mm) | DaylightHours (h) | CTmax (°C) |
|---|---|---|---|---|---|---|---|---|---|
| Rana uenoi | RU-1 | 30 | 37.53342 | 127.3807414 | 100 | 10.35 | 97.25 | 6.47 | 37.0 ± 0.5 |
| RU-2 | 29 | 37.64184 | 128.0233030 | 289 | 9.4 | 93.3 | 6.96 | 37.4 ± 0.4 | |
| RU-3 | 24 | 37.66629 | 128.8459008 | 190 | 10.8 | 112.05 | 7.02 | 37.5 ± 0.2 | |
| RU-4 | 20 | 36.40426 | 126.8289935 | 33 | 8.2 | 127.9 | 6.55 | 37.1 ± 0.4 | |
| RU-5 | 24 | 36.48659 | 127.7382435 | 216 | 9.8 | 116.35 | 6.61 | 37.2 ± 0.1 | |
| RU-6 | 20 | 36.90107 | 128.5241611 | 267 | 10.05 | 142.55 | 7.38 | 36.7 ± 0.1 | |
| RU-7 | 28 | 35.17369 | 126.5375166 | 134 | 10.2 | 119.65 | 7.28 | 35.9 ± 0.4 | |
| RU-8 | 24 | 35.10432 | 127.7560522 | 137 | 12.05 | 139.35 | 6.98 | 36.5 ± 0.3 | |
| RU-9 | 24 | 35.31685 | 128.6274404 | 40 | 11.95 | 154.25 | 7.09 | 36.6 ± 0.4 | |
| RU-10 | 23 | 34.44065 | 126.6373606 | 77 | 10.5 | 133.85 | 6.49 | 35.4 ± 0.2 | |
| RU-11 | 24 | 34.79935 | 127.3308623 | 62 | 11 | 162.35 | 6.79 | 36.1 ± 0.3 | |
| Bufo sachalinensis | BG-1 | 24 | 37.86783 | 126.8645720 | 88 | 8.2 | 79.35 | 7.03 | 38.1 ± 0.2 |
| BG-2 | 24 | 37.51784 | 126.8813251 | 13 | 10.55 | 89.9 | 7.09 | 38.0 ± 0.1 | |
| BG-3 | 24 | 36.90107 | 128.5241611 | 344 | 10.8 | 112.05 | 7.02 | 38.5 ± 0.1 | |
| BG-4 | 24 | 36.20582 | 126.9009314 | 44 | 10.2 | 127.9 | 6.55 | 38.5 ± 0.2 | |
| BG-5 | 24 | 36.48607 | 127.7377301 | 216 | 9.8 | 116.35 | 6.61 | 38.1 ± 0.1 | |
| BG-6 | 24 | 35.82585 | 128.7048109 | 81 | 10.05 | 142.55 | 7.38 | 38.5 ± 0.2 | |
| BG-7 | 24 | 35.18138 | 126.5409766 | 157 | 10.2 | 119.65 | 7.28 | 38.2 ± 0.1 | |
| BG-8 | 24 | 35.14597 | 127.6870155 | 23 | 12.05 | 139.35 | 6.98 | 38.1 ± 0.2 | |
| BG-9 | 24 | 35.29143 | 128.6729614 | 44 | 11.95 | 154.25 | 7.09 | 38.4 ± 0.3 | |
| BG-10 | 24 | 34.76735 | 126.4103310 | 188 | 8.2 | 160.85 | 6.81 | 38.0 ± 0.1 |
| Species | Fixed Factors | Estimate ± SE | DF | t-Value | p-Value | VIF |
|---|---|---|---|---|---|---|
| Rana uenoi | (Intercept) | 15.93 ± 7.35 | 259 | 2.17 | 0.0312 | |
| Latitude | 0.59 ± 0.17 | 5 | 3.46 | 0.018 * | 2.65 | |
| Altitude | 0 ± 0 | 5 | −0.03 | 0.979 | 2.4 | |
| Sunset | −0.24 ± 0.55 | 5 | −0.44 | 0.680 | 1.88 | |
| Average temperature | 0.03 ± 0.14 | 5 | 0.22 | 0.832 | 1.43 | |
| Precipitation | 0.01 ± 0.01 | 5 | 0.75 | 0.407 | 2.8 | |
| Bufo sachalinensis | (Intercept) | 35.19 ± 6.62 | 227 | 5.32 | <0.001 | |
| Latitude | 0.09 ± 0.16 | 7 | 0.53 | 0.474 | 4.95 | |
| Altitude | 0 ± 0 | 7 | 0.51 | 0.687 | 1.08 | |
| Sunset | −0.15 ± 0.21 | 7 | −0.75 | 0.273 | 1.04 | |
| Average temperature | 0.07 ± 0.06 | 7 | 1.05 | 0.251 | 1.15 | |
| Precipitation | 0 ± 0.01 | 7 | 0.44 | 0.66 | 4.95 |
| Species | Fixed Factors | B | SE | z Value | p Value |
|---|---|---|---|---|---|
| Rana uenoi | Intercept | −1.73 | 0.14 | −11.5 | <0.001 * |
| Time | 0.06 | 0.03 | 1.59 | 0.11 | |
| Bufo sachalinensis | Intercept | 0.96 | 0.2 | 4.43 | <0.001 * |
| Time | −0.34 | 0.02 | −11.86 | <0.001 * |
| Species | Fixed Factors | B | SE | z Value | p Value |
|---|---|---|---|---|---|
| Rana uenoi | Intercept | −3.28 | 0.25 | −13.11 | <0.001 * |
| Time | 0.2 | 0.05 | 3.63 | <0.001 * | |
| Bufo sachalinensis | Intercept | −4 | 0.39 | −10.18 | <0.001 * |
| Time | 0.37 | 0.04 | 8.05 | <0.001 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.I.; Chuang, M.-F.; Borzée, A.; Kwon, S.; Jang, Y. Latitude-Induced and Behaviorally Thermoregulated Variations in Upper Thermal Tolerance of Two Anuran Species. Biology 2022, 11, 1506. https://doi.org/10.3390/biology11101506
Kim YI, Chuang M-F, Borzée A, Kwon S, Jang Y. Latitude-Induced and Behaviorally Thermoregulated Variations in Upper Thermal Tolerance of Two Anuran Species. Biology. 2022; 11(10):1506. https://doi.org/10.3390/biology11101506
Chicago/Turabian StyleKim, Ye Inn, Ming-Feng Chuang, Amaël Borzée, Sera Kwon, and Yikweon Jang. 2022. "Latitude-Induced and Behaviorally Thermoregulated Variations in Upper Thermal Tolerance of Two Anuran Species" Biology 11, no. 10: 1506. https://doi.org/10.3390/biology11101506