Enhancement of Antiviral T-Cell Responses by Vitamin C Suggests New Strategies to Improve Manufacturing of Virus-Specific T Cells for Adoptive Immunotherapy

 , ,
, ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of PBMCs and T Cells
2.2. Cell Culture
2.3. Vitamin C and Vitamin D
2.4. Expansion of CMV-Specific T Cells with Artificial Antigen Presenting Cell Beads (aAPCs) and Vitamins
2.5. Enrichment of IFN-γ Secreting CMV-Specific T Cells by Cytokine Secretion Assay
2.6. Antigen-Specific T-Cell Response Determined by IFN-γ ELISpot Assay
2.7. Flow Cytometry
2.8. Intracellular Cytokine Staining
2.9. Statistics
3. Results
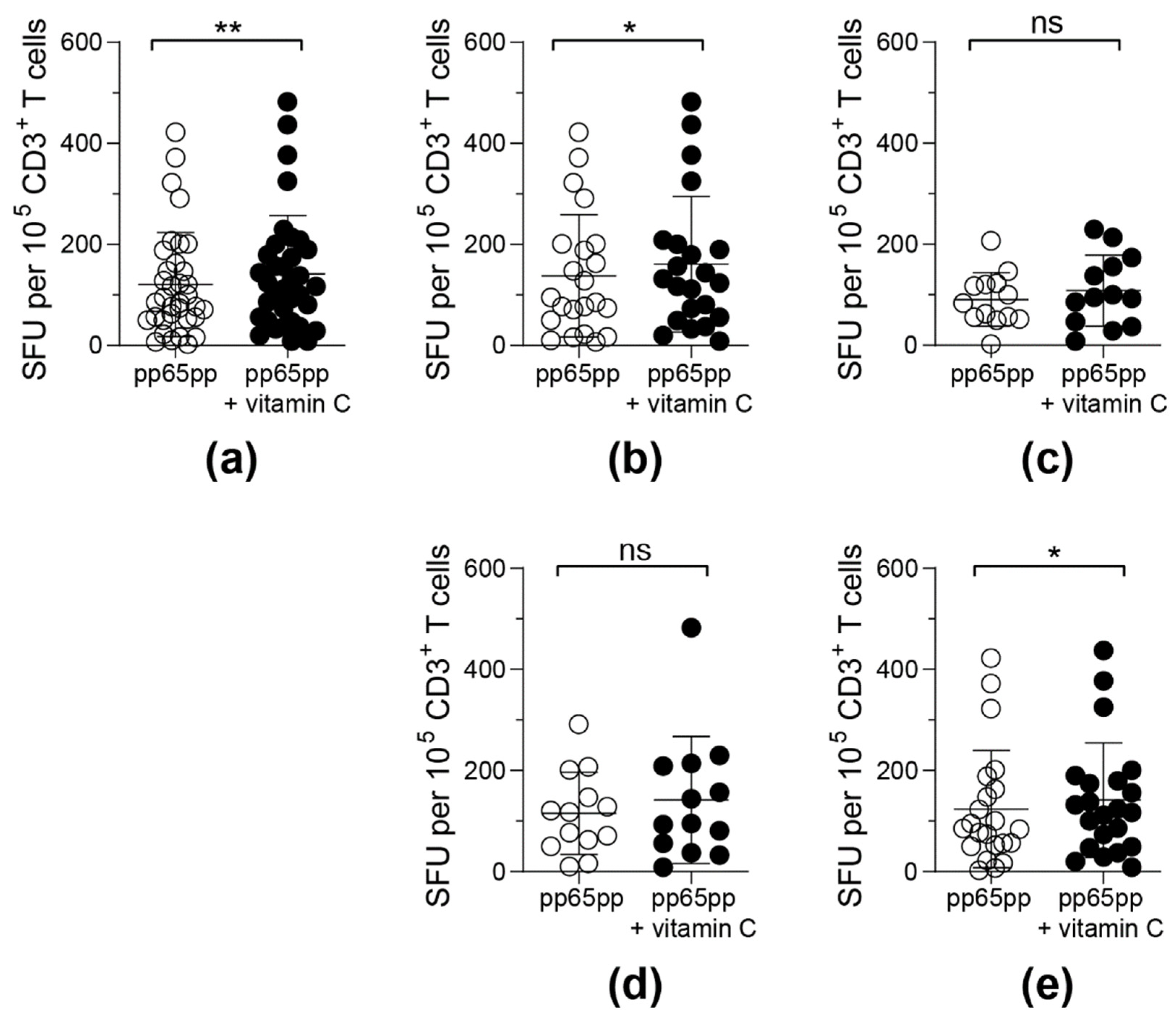
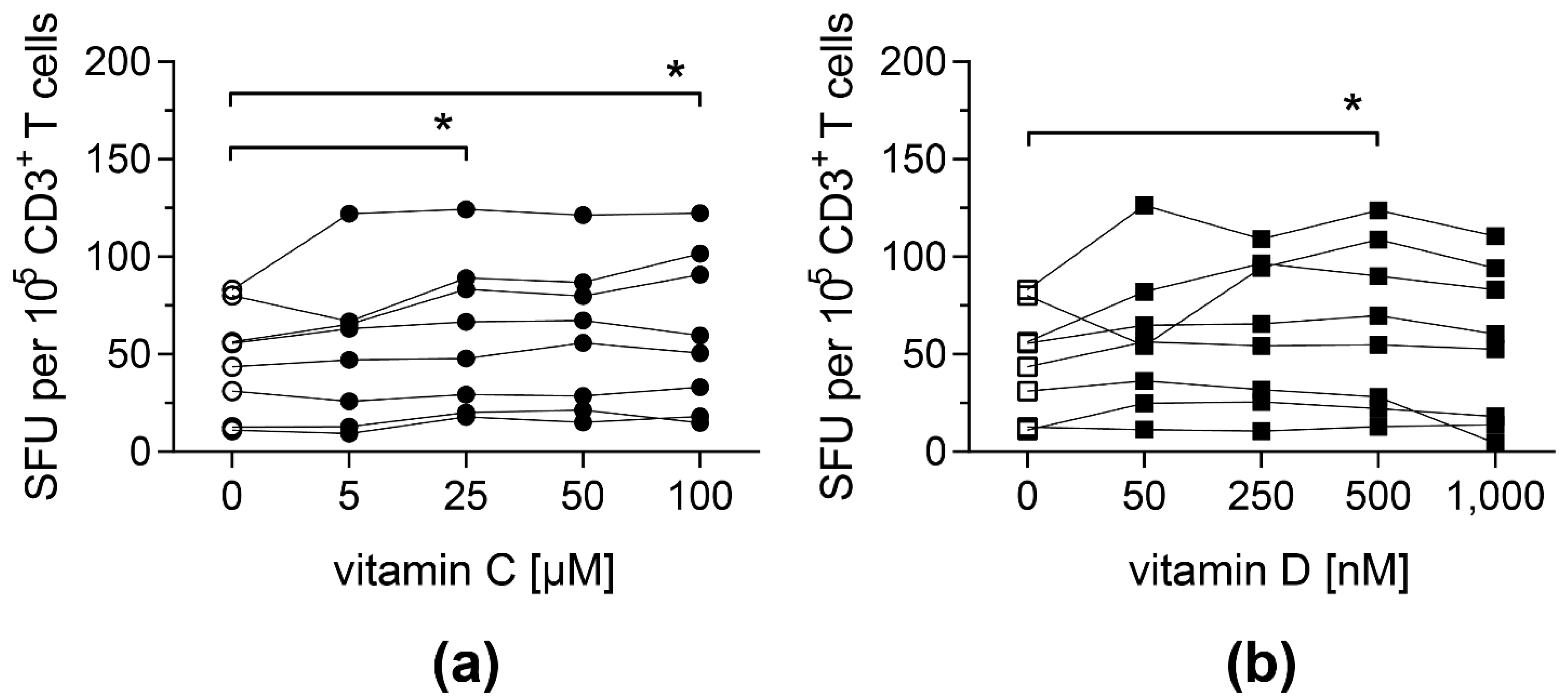
3.1. Antigen-Specific T-Cell Response against pp65 Is Enhanced upon Vitamin C and Vitamin D Supplementation
3.2. Vitamin C, but Not Vitamin D, Promotes the Expansion of CMV-Specific Cytotoxic T Cells
3.3. The Yield of CMV-Specific T Cells Using a Cytokine Secretion Assay Is Slightly Increased in the Presence of Vitamin C
3.4. Antigen-Specific T-Cell Functionality Can Be Improved by Pre-Treatment with Vitamin C for 3 Days
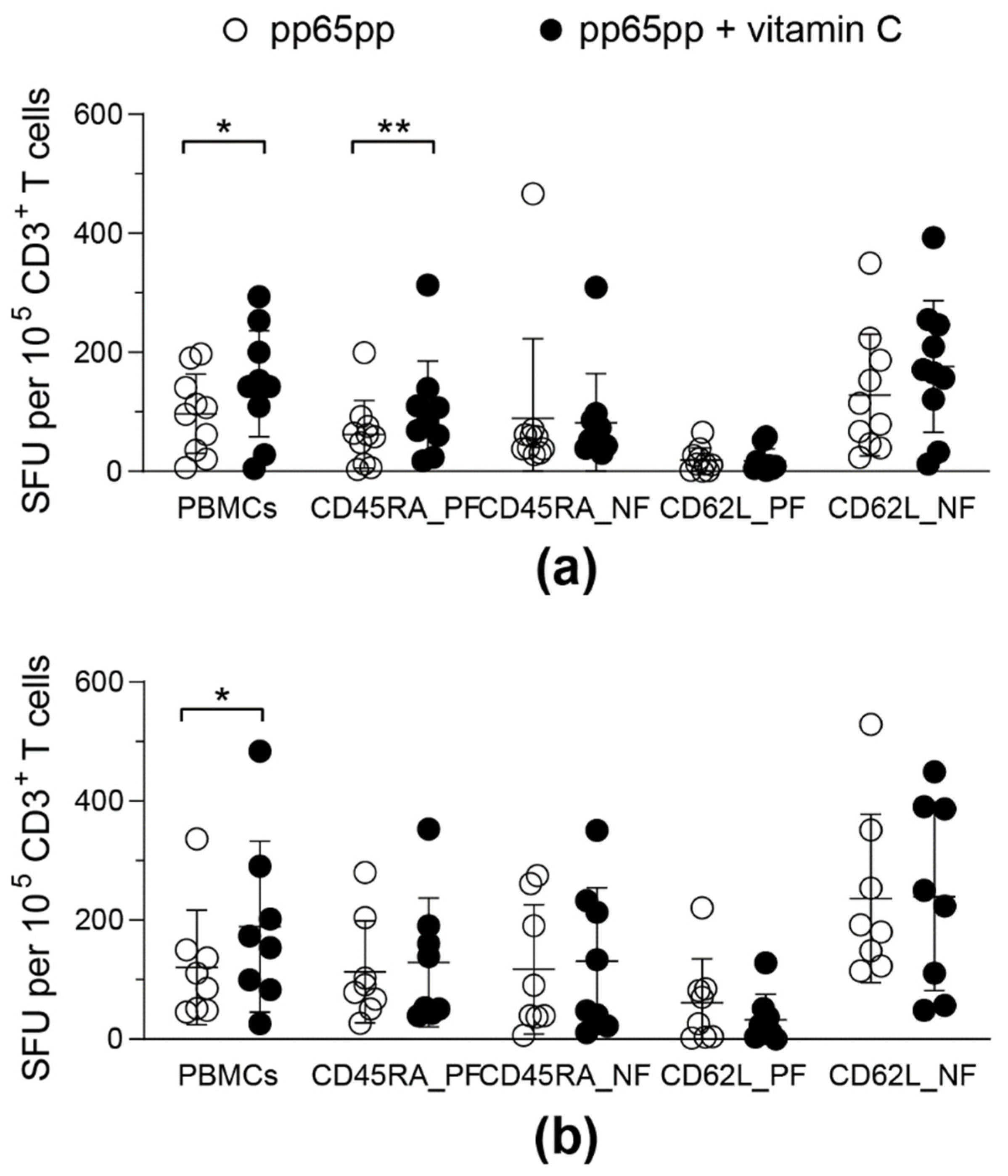
3.5. Vitamin C Does Not Differently Affect Distinct T-Cell Memory Subsets
3.6. Vitamin C Enhances IFN-γ Secretion by CD45RA+ T Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Narinx, J.; Servais, S.; Baron, F.; Beguin, Y.; Willems, E. Allogeneic hematopoietic stem cell transplantation: General principles and recent progress. Rev. Med. Liege. 2021, 76, 464–469. [Google Scholar] [PubMed]
- Wingard, J.R.; Hsu, J.; Hiemenz, J.W. Hematopoietic stem cell transplantation: An overview of infection risks and epidemiology. Hematol. Oncol. Clin. N. Am. 2011, 25, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Meyers, J.D.; Flournoy, N.; Thomas, E.D. Risk factors for cytomegalovirus infection after human marrow transplantation. J. Infect. Dis. 1986, 153, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Ljungman, P. Molecular monitoring of viral infections after hematopoietic stem cell transplantation. Int. J. Hematol. 2010, 91, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Seo, S. Respiratory viral infections occurring after hematopoietic stem cell transplantation. Rinsho Ketsueki 2017, 58, 818–826. [Google Scholar]
- Alexandersson, A.; Koskenvuo, M.; Tiderman, A.; Laaperi, M.; Huttunen, P.; Saarinen-Pihkala, U.; Anttila, V.J.; Lautenschlager, I.; Taskinen, M. Viral infections and immune reconstitution interaction after pediatric allogenic hematopoietic stem cell transplantation. Infect. Dis. 2019, 51, 772–778. [Google Scholar] [CrossRef]
- Mori, T.; Okamoto, S.; Matsuoka, S.; Yajima, T.; Wakui, M.; Watanabe, R.; Ishida, A.; Iwao, Y.; Mukai, M.; Hibi, T.; et al. Risk-adapted pre-emptive therapy for cytomegalovirus disease in patients undergoing allogeneic bone marrow transplantation. Bone Marrow Transpl. 2000, 25, 765–769. [Google Scholar] [CrossRef]
- Reed, E.C.; Bowden, R.A.; Dandliker, P.S.; Lilleby, K.E.; Meyers, J.D. Treatment of cytomegalovirus pneumonia with ganciclovir and intravenous cytomegalovirus immunoglobulin in patients with bone marrow transplants. Ann. Intern. Med. 1988, 109, 783–788. [Google Scholar] [CrossRef]
- Salzberger, B.; Bowden, R.A.; Hackman, R.C.; Davis, C.; Boeckh, M. Neutropenia in allogeneic marrow transplant recipients receiving ganciclovir for prevention of cytomegalovirus disease: Risk factors and outcome. Blood 1997, 90, 2502–2508. [Google Scholar] [CrossRef] [Green Version]
- Saglio, F.; Hanley, P.J.; Bollard, C.M. The time is now: Moving toward virus-specific t cells after allogeneic hematopoietic stem cell transplantation as the standard of care. Cytotherapy 2014, 16, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulou, A.; Gerdemann, U.; Katari, U.L.; Tzannou, I.; Liu, H.; Martinez, C.; Leung, K.; Carrum, G.; Gee, A.P.; Vera, J.F.; et al. Activity of broad-spectrum t cells as treatment for adv, ebv, cmv, bkv, and hhv6 infections after hsct. Sci. Transl. Med. 2014, 6, 242ra283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobbold, M.; Khan, N.; Pourgheysari, B.; Tauro, S.; McDonald, D.; Osman, H.; Assenmacher, M.; Billingham, L.; Steward, C.; Crawley, C.; et al. Adoptive transfer of cytomegalovirus-specific ctl to stem cell transplant patients after selection by hla-peptide tetramers. J. Exp. Med. 2005, 202, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Gerdemann, U.; Keirnan, J.M.; Katari, U.L.; Yanagisawa, R.; Christin, A.S.; Huye, L.E.; Perna, S.K.; Ennamuri, S.; Gottschalk, S.; Brenner, M.K.; et al. Rapidly generated multivirus-specific cytotoxic t lymphocytes for the prophylaxis and treatment of viral infections. Mol. Ther. 2012, 20, 1622–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bono, M.R.; Tejon, G.; Flores-Santibanez, F.; Fernandez, D.; Rosemblatt, M.; Sauma, D. Retinoic acid as a modulator of t cell immunity. Nutrients 2016, 8, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaible, U.E.; Kaufmann, S.H. Malnutrition and infection: Complex mechanisms and global impacts. PLoS Med. 2007, 4, e115. [Google Scholar] [CrossRef] [Green Version]
- Yoshii, K.; Hosomi, K.; Sawane, K.; Kunisawa, J. Metabolism of dietary and microbial vitamin b family in the regulation of host immunity. Front Nutr. 2019, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Carr, A.C.; Maggini, S. Vitamin c and immune function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef] [Green Version]
- Steiling, H.; Longet, K.; Moodycliffe, A.; Mansourian, R.; Bertschy, E.; Smola, H.; Mauch, C.; Williamson, G. Sodium-dependent vitamin c transporter isoforms in skin: Distribution, kinetics, and effect of uvb-induced oxidative stress. Free Radic. Biol. Med. 2007, 43, 752–762. [Google Scholar] [CrossRef]
- Washko, P.; Rotrosen, D.; Levine, M. Ascorbic acid transport and accumulation in human neutrophils. J. Biol. Chem. 1989, 264, 18996–19002. [Google Scholar] [CrossRef]
- Tan, P.H.; Sagoo, P.; Chan, C.; Yates, J.B.; Campbell, J.; Beutelspacher, S.C.; Foxwell, B.M.; Lombardi, G.; George, A.J. Inhibition of nf-kappa b and oxidative pathways in human dendritic cells by antioxidative vitamins generates regulatory t cells. J. Immunol. 2005, 174, 7633–7644. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.M.; Kim, J.H.; Kang, J.S.; Lee, W.J.; Hwang, Y.I. Vitamin c is taken up by human t cells via sodium-dependent vitamin c transporter 2 (svct2) and exerts inhibitory effects on the activation of these cells in vitro. Anat. Cell Biol. 2016, 49, 88–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huijskens, M.J.; Walczak, M.; Koller, N.; Briede, J.J.; Senden-Gijsbers, B.L.; Schnijderberg, M.C.; Bos, G.M.; Germeraad, W.T. Technical advance: Ascorbic acid induces development of double-positive t cells from human hematopoietic stem cells in the absence of stromal cells. J. Leukoc. Biol. 2014, 96, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Manning, J.; Mitchell, B.; Appadurai, D.A.; Shakya, A.; Pierce, L.J.; Wang, H.; Nganga, V.; Swanson, P.C.; May, J.M.; Tantin, D.; et al. Vitamin c promotes maturation of t-cells. Antioxid. Redox Signal. 2013, 19, 2054–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, J.D.; Cole, M.; Bunditrutavorn, B.; Vella, A.T. Ascorbic acid is a potent inhibitor of various forms of t cell apoptosis. Cell Immunol. 1999, 194, 1–5. [Google Scholar] [CrossRef]
- Noh, K.; Lim, H.; Moon, S.K.; Kang, J.S.; Lee, W.J.; Lee, D.; Hwang, Y.I. Mega-dose vitamin c modulates t cell functions in balb/c mice only when administered during t cell activation. Immunol. Lett. 2005, 98, 63–72. [Google Scholar] [CrossRef]
- van Gorkom, G.N.Y.; Klein Wolterink, R.G.J.; Van Elssen, C.; Wieten, L.; Germeraad, W.T.V.; Bos, G.M.J. Influence of vitamin c on lymphocytes: An overview. Antioxidants 2018, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Song, M.H.; Nair, V.S.; Oh, K.I. Vitamin c enhances the expression of il17 in a jmjd2-dependent manner. BMB Rep. 2017, 50, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Nikolouli, E.; Hardtke-Wolenski, M.; Hapke, M.; Beckstette, M.; Geffers, R.; Floess, S.; Jaeckel, E.; Huehn, J. Alloantigen-induced regulatory t cells generated in presence of vitamin c display enhanced stability of foxp3 expression and promote skin allograft acceptance. Front Immunol. 2017, 8, 748. [Google Scholar] [CrossRef] [Green Version]
- Sasidharan Nair, V.; Song, M.H.; Oh, K.I. Vitamin c facilitates demethylation of the foxp3 enhancer in a tet-dependent manner. J. Immunol. 2016, 196, 2119–2131. [Google Scholar] [CrossRef] [Green Version]
- Kasahara, H.; Kondo, T.; Nakatsukasa, H.; Chikuma, S.; Ito, M.; Ando, M.; Kurebayashi, Y.; Sekiya, T.; Yamada, T.; Okamoto, S.; et al. Generation of allo-antigen-specific induced treg stabilized by vitamin c treatment and its application for prevention of acute graft versus host disease model. Int. Immunol. 2017, 29, 457–469. [Google Scholar] [CrossRef] [Green Version]
- Yue, X.; Trifari, S.; Aijo, T.; Tsagaratou, A.; Pastor, W.A.; Zepeda-Martinez, J.A.; Lio, C.W.; Li, X.; Huang, Y.; Vijayanand, P.; et al. Control of foxp3 stability through modulation of tet activity. J. Exp. Med. 2016, 213, 377–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, J.I.; Zuchner, S.; Wang, G. Regulation of the epigenome by vitamin c. Annu. Rev. Nutr. 2015, 35, 545–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, M.T.; Lee, Y.K.; Maynard, C.L.; Oliver, J.R.; Bikle, D.D.; Jetten, A.M.; Weaver, C.T. Lineage-specific effects of 1,25-dihydroxyvitamin d(3) on the development of effector cd4 t cells. J. Biol. Chem. 2011, 286, 997–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staeva-Vieira, T.P.; Freedman, L.P. 1,25-dihydroxyvitamin d3 inhibits ifn-gamma and il-4 levels during in vitro polarization of primary murine cd4+ t cells. J. Immunol. 2002, 168, 1181–1189. [Google Scholar] [CrossRef] [Green Version]
- Cantorna, M.T.; Snyder, L.; Lin, Y.D.; Yang, L. Vitamin d and 1,25(oh)2d regulation of t cells. Nutrients 2015, 7, 3011–3021. [Google Scholar] [CrossRef] [Green Version]
- Korf, H.; Wenes, M.; Stijlemans, B.; Takiishi, T.; Robert, S.; Miani, M.; Eizirik, D.L.; Gysemans, C.; Mathieu, C. 1,25-dihydroxyvitamin d3 curtails the inflammatory and t cell stimulatory capacity of macrophages through an il-10-dependent mechanism. Immunobiology 2012, 217, 1292–1300. [Google Scholar] [CrossRef]
- von Essen, M.R.; Kongsbak, M.; Schjerling, P.; Olgaard, K.; Odum, N.; Geisler, C. Vitamin d controls t cell antigen receptor signaling and activation of human t cells. Nat. Immunol. 2010, 11, 344–349. [Google Scholar] [CrossRef]
- Mangare, C.; Tischer-Zimmermann, S.; Riese, S.B.; Dragon, A.C.; Prinz, I.; Blasczyk, R.; Maecker-Kolhoff, B.; Eiz-Vesper, B. Robust identification of suitable t-cell subsets for personalized cmv-specific t-cell immunotherapy using cd45ra and cd62l microbeads. Int. J. Mol. Sci. 2019, 20, 1415. [Google Scholar] [CrossRef] [Green Version]
- Oelke, M.; Maus, M.V.; Didiano, D.; June, C.H.; Mackensen, A.; Schneck, J.P. Ex vivo induction and expansion of antigen-specific cytotoxic t cells by hla-ig-coated artificial antigen-presenting cells. Nat. Med. 2003, 9, 619–624. [Google Scholar] [CrossRef]
- Levine, M.; Dhariwal, K.R.; Welch, R.W.; Wang, Y.; Park, J.B. Determination of optimal vitamin c requirements in humans. Am. J. Clin. Nutr. 1995, 62, 1347S–1356S. [Google Scholar] [CrossRef]
- Carr, A.C.; Frei, B. Toward a new recommended dietary allowance for vitamin c based on antioxidant and health effects in humans. Am. J. Clin. Nutr. 1999, 69, 1086–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lykkesfeldt, J.; Tveden-Nyborg, P. The pharmacokinetics of vitamin c. Nutrients 2019, 11, 2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassoy, E.Y.; Walch, M.; Martinvalet, D. Reactive oxygen species: Do they play a role in adaptive immunity? Front. Immunol. 2021, 12, 755856. [Google Scholar] [CrossRef] [PubMed]
- Iamsawat, S.; Tian, L.; Daenthanasanmak, A.; Wu, Y.; Nguyen, H.D.; Bastian, D.; Yu, X.Z. Vitamin c stabilizes cd8+ itregs and enhances their therapeutic potential in controlling murine gvhd and leukemia relapse. Blood Adv. 2019, 3, 4187–4201. [Google Scholar] [CrossRef] [Green Version]
- Constantini, N.W.; Dubnov-Raz, G.; Eyal, B.B.; Berry, E.M.; Cohen, A.H.; Hemila, H. The effect of vitamin c on upper respiratory infections in adolescent swimmers: A randomized trial. Eur. J. Pediatr. 2011, 170, 59–63. [Google Scholar] [CrossRef]
- Kim, J.; Kim, A.R.; Shin, E.C. Cytomegalovirus infection and memory t cell inflation. Immune. Netw. 2015, 15, 186–190. [Google Scholar] [CrossRef] [Green Version]
- Appay, V.; van Lier, R.A.; Sallusto, F.; Roederer, M. Phenotype and function of human t lymphocyte subsets: Consensus and issues. Cytometry A 2008, 73, 975–983. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laubert, M.; Bonifacius, A.; Dragon, A.C.; Mangare, C.; Blasczyk, R.; Huehn, J.; Eiz-Vesper, B. Enhancement of Antiviral T-Cell Responses by Vitamin C Suggests New Strategies to Improve Manufacturing of Virus-Specific T Cells for Adoptive Immunotherapy. Biology 2022, 11, 536. https://doi.org/10.3390/biology11040536
Laubert M, Bonifacius A, Dragon AC, Mangare C, Blasczyk R, Huehn J, Eiz-Vesper B. Enhancement of Antiviral T-Cell Responses by Vitamin C Suggests New Strategies to Improve Manufacturing of Virus-Specific T Cells for Adoptive Immunotherapy. Biology. 2022; 11(4):536. https://doi.org/10.3390/biology11040536
Chicago/Turabian StyleLaubert, Miriam, Agnes Bonifacius, Anna Christina Dragon, Caroline Mangare, Rainer Blasczyk, Jochen Huehn, and Britta Eiz-Vesper. 2022. "Enhancement of Antiviral T-Cell Responses by Vitamin C Suggests New Strategies to Improve Manufacturing of Virus-Specific T Cells for Adoptive Immunotherapy" Biology 11, no. 4: 536. https://doi.org/10.3390/biology11040536