
Staphylococcus aureus α-Toxin Effect on Acinetobacter baumannii Behavior

 , ,
, ,  and
and 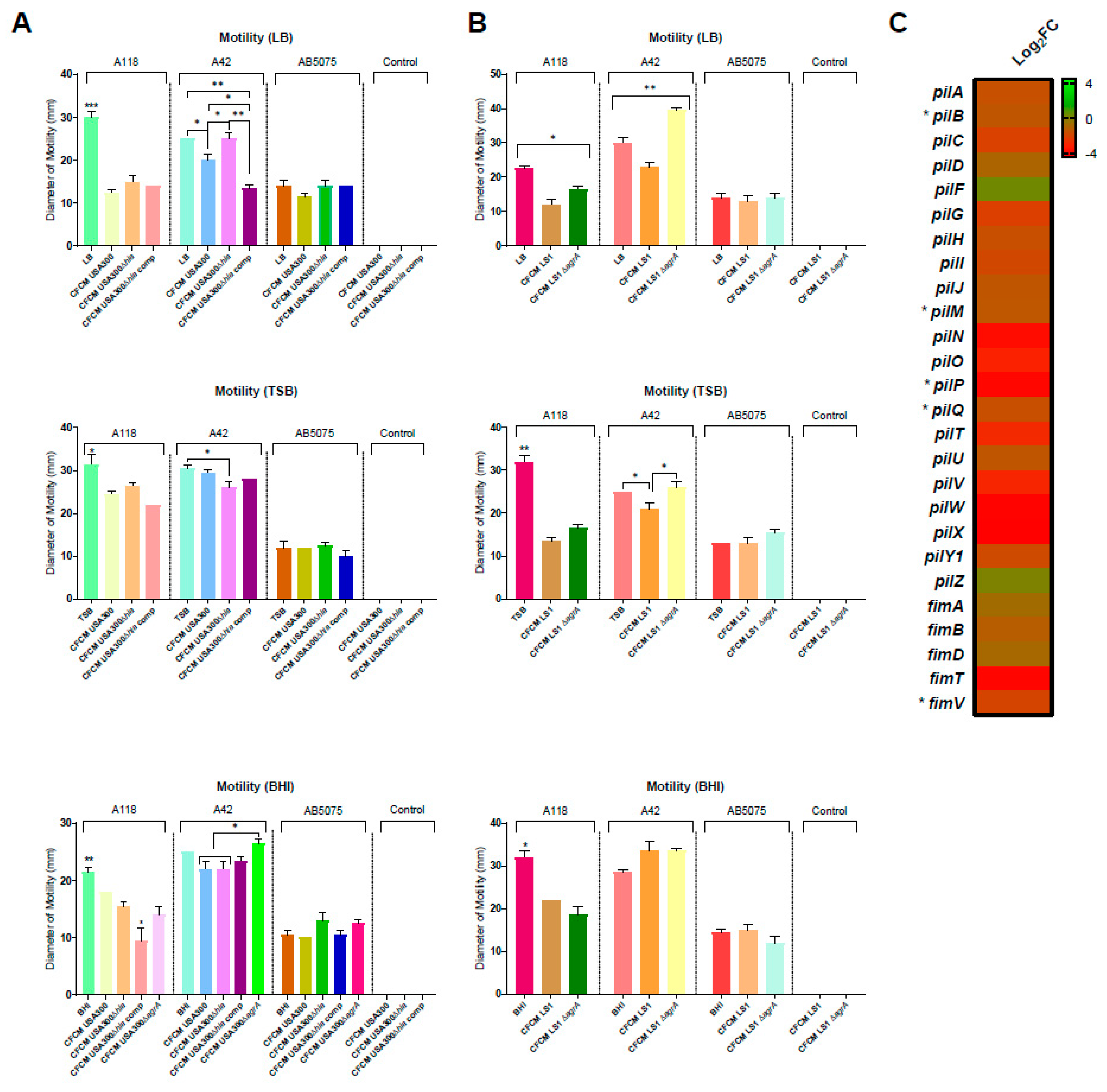{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Cell-Free Conditioned Media (CFCM)
2.3. Motility Assays
2.4. Hemolytic and Fibrinolytic Activity Assays
2.5. Biofilm Formation
2.6. Susceptibility Assays
2.7. Zone of Clearance (ZOC) Assays
2.8. RNA Extraction, Sequencing and Analysis
2.9. Statistical Analysis
3. Results
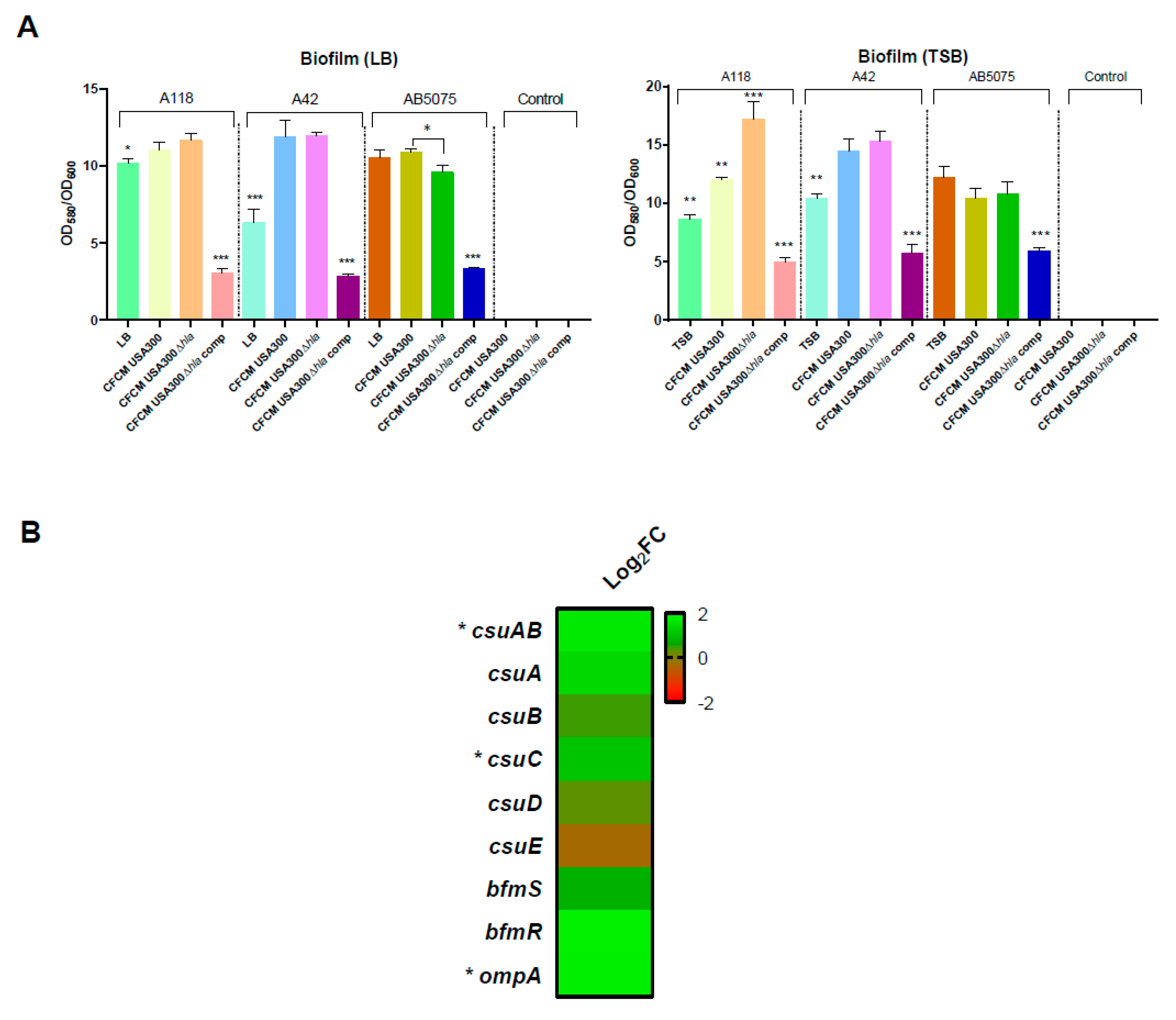
3.1. Effect on Motility and Biofilm Formation
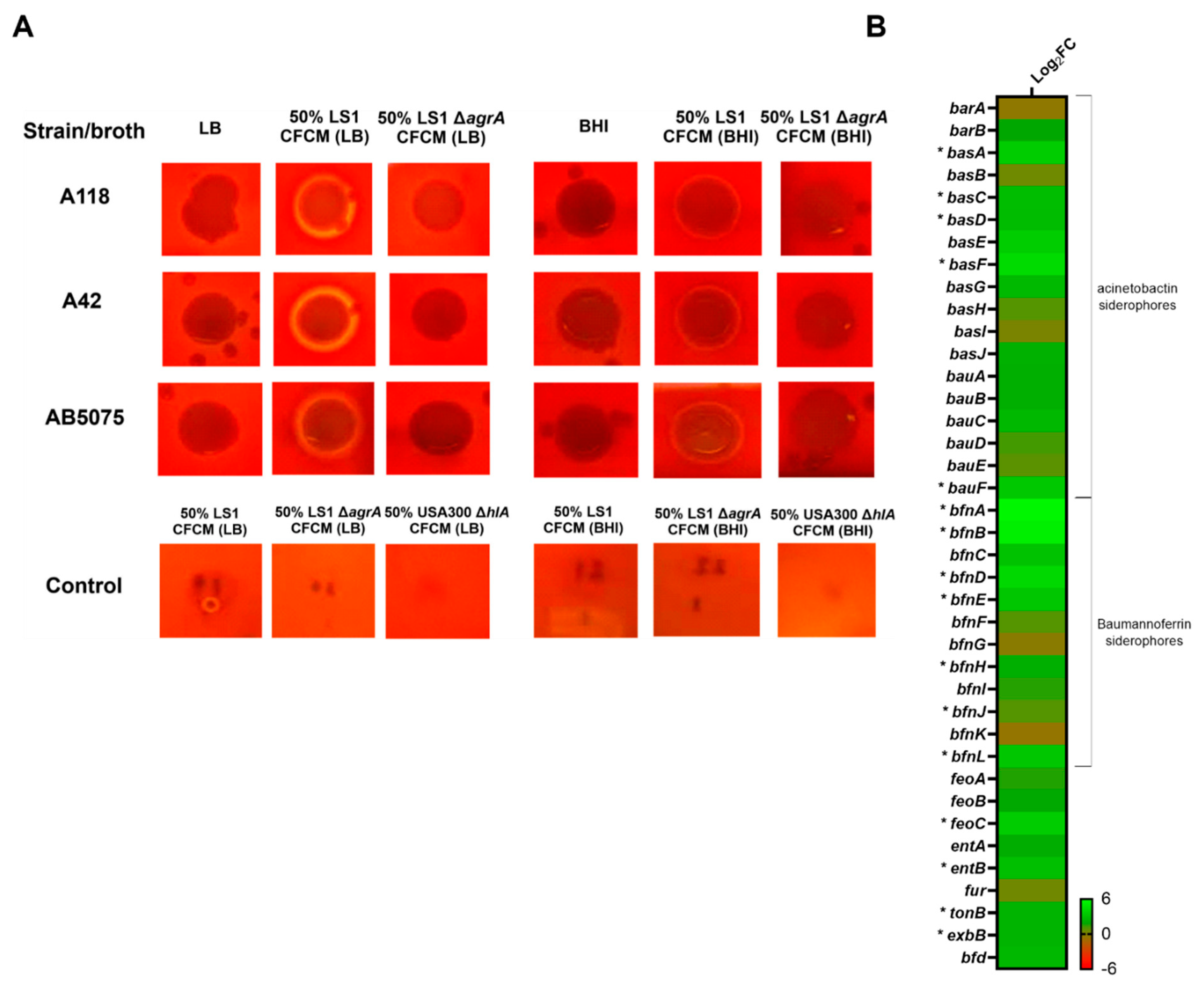
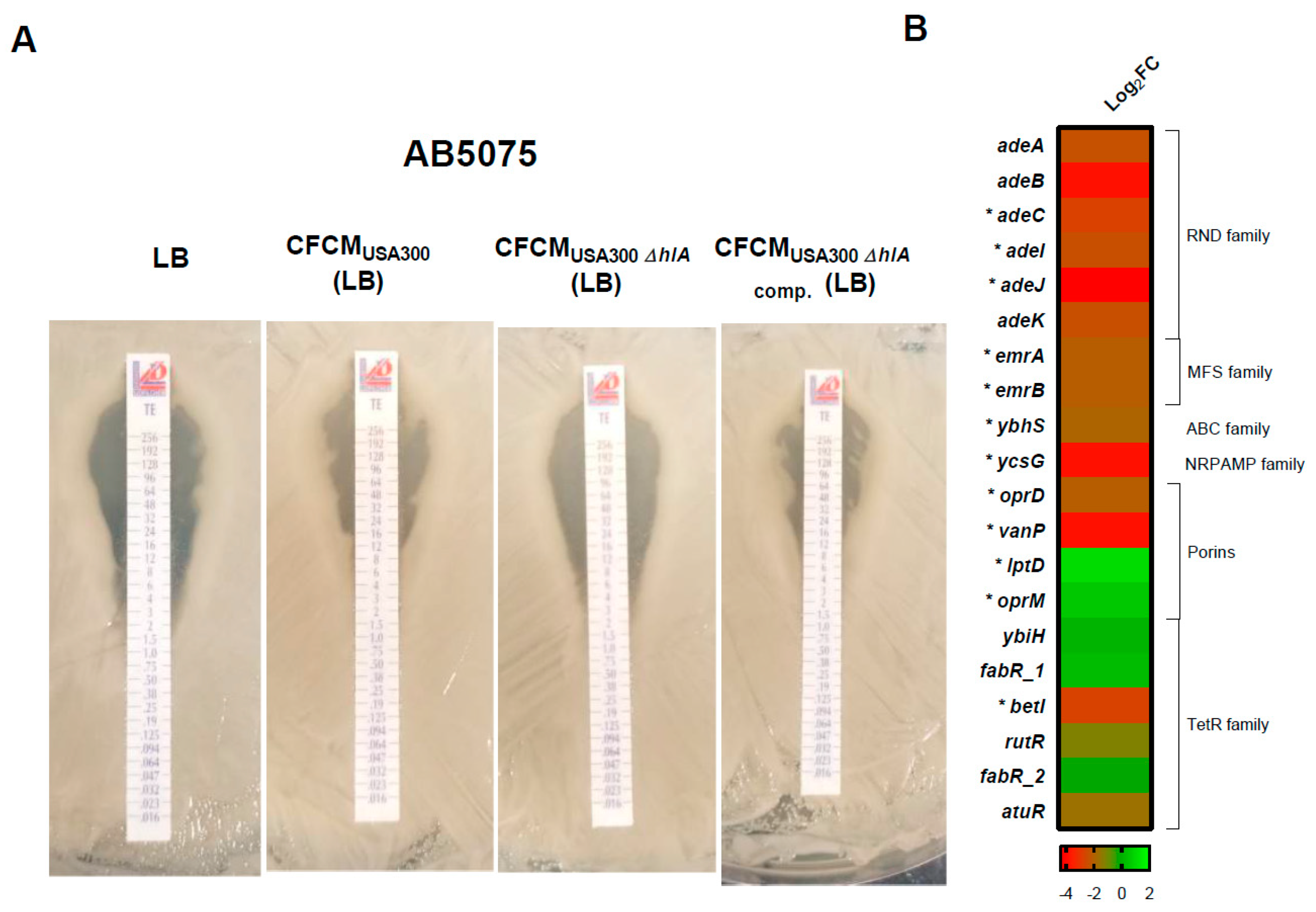
3.2. Effect of S. aureus-Released Molecules on Other Phenotypes
3.3. RNA Sequencing Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, A.P.; Choi, Y.; Brinkac, L.M.; Krishnakumar, R.; DePew, J.; Kim, M.; Hinkle, M.K.; Lesho, E.P.; Fouts, D.E. Multidrug resistant pathogens respond differently to the presence of co-pathogen, commensal, probiotic and host cells. Sci. Rep. 2018, 8, 8656. [Google Scholar] [CrossRef]
- Filkins, L.M.; O’Toole, G.A. Cystic Fibrosis Lung Infections: Polymicrobial, Complex, and Hard to Treat. PLoS Pathog. 2015, 11, e1005258. [Google Scholar] [CrossRef] [Green Version]
- Yo, C.H.; Hsein, Y.C.; Wu, Y.L.; Hsu, W.T.; Ma, M.H.; Tsai, C.H.; Chen, S.C.; Lee, C.C. Clinical predictors and outcome impact of community-onset polymicrobial bloodstream infection. Int. J. Antimicrob. Agents 2019, 54, 716–722. [Google Scholar] [CrossRef]
- Löwik, C.A.M.; Zijlstra, W.P.; Knobben, B.A.S.; Ploegmakers, J.J.W.; Dijkstra, B.; de Vries, A.J.; Kampinga, G.A.; Mithoe, G.; Al Moujahid, A.; Jutte, P.C.; et al. Obese patients have higher rates of polymicrobial and Gram-negative early periprosthetic joint infections of the hip than non-obese patients. PLoS ONE 2019, 14, e0215035. [Google Scholar] [CrossRef]
- Stacy, A.; McNally, L.; Darch, S.E.; Brown, S.P.; Whiteley, M. The biogeography of polymicrobial infection. Nat. Rev. Microbiol. 2016, 14, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Baldan, R.; Cigana, C.; Testa, F.; Bianconi, I.; De Simone, M.; Pellin, D.; Di Serio, C.; Bragonzi, A.; Cirillo, D.M. Adaptation of Pseudomonas aeruginosa in Cystic Fibrosis airways influences virulence of Staphylococcus aureus in vitro and murine models of co-infection. PLoS ONE 2014, 9, e89614. [Google Scholar] [CrossRef]
- Davies, J.C.; Rubin, B.K. Emerging and unusual gram-negative infections in cystic fibrosis. Semin. Respir. Crit. Care Med. 2007, 28, 312–321. [Google Scholar] [CrossRef]
- Safdar, N.; Maki, D.G. The commonality of risk factors for nosocomial colonization and infection with antimicrobial-resistant Staphylococcus aureus, enterococcus, gram-negative bacilli, Clostridium difficile, and Candida. Ann. Intern. Med. 2002, 136, 834–844. [Google Scholar] [CrossRef]
- Castellanos, N.; Nakanouchi, J.; Yuzen, D.I.; Fung, S.; Fernandez, J.S.; Barberis, C.; Tuchscherr, L.; Ramirez, M.S. A Study on Acinetobacter baumannii and Staphylococcus aureus Strains Recovered from the Same Infection Site of a Diabetic Patient. Curr. Microbiol. 2019, 76, 842–847. [Google Scholar] [CrossRef]
- Bhakdi, S.; Tranum-Jensen, J. Alpha-toxin of Staphylococcus aureus. Microbiol. Rev. 1991, 55, 733–751. [Google Scholar] [CrossRef]
- Xiong, Y.Q.; Willard, J.; Yeaman, M.R.; Cheung, A.L.; Bayer, A.S. Regulation of Staphylococcus aureus alpha-toxin gene (hla) expression by agr, sarA, and sae in vitro and in experimental infective endocarditis. J. Infect. Dis. 2006, 194, 1267–1275. [Google Scholar] [CrossRef] [Green Version]
- Cohen, T.S.; Hilliard, J.J.; Jones-Nelson, O.; Keller, A.E.; O’Day, T.; Tkaczyk, C.; DiGiandomenico, A.; Hamilton, M.; Pelletier, M.; Wang, Q.; et al. Staphylococcus aureus alpha toxin potentiates opportunistic bacterial lung infections. Sci. Transl. Med. 2016, 8, 329ra31. [Google Scholar] [CrossRef] [Green Version]
- Vilacoba, E.; Almuzara, M.; Gulone, L.; Traglia, G.M.; Montana, S.; Rodriguez, H.; Pasteran, F.; Pennini, M.; Sucari, A.; Gomez, N.; et al. Widespread dispersion of the resistance element tet(B)::ISCR2 in XDR Acinetobacter baumannii isolates. Epidemiol. Infect. 2016, 144, 1574–1578. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, C.; Le, C.; Tuttobene, M.R.; Subils, T.; Martinez, J.; Sieira, R.; Papp-Wallace, K.M.; Keppetipola, N.; Bonomo, R.A.; Actis, L.A.; et al. Human Pleural Fluid and Human Serum Albumin Modulate the Behavior of a Hypervirulent and Multidrug-Resistant (MDR) Acinetobacter baumannii Representative Strain. Pathogens 2021, 10, 471. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Penwell, W.F.; Traglia, G.M.; Zimbler, D.L.; Gaddy, J.A.; Nikolaidis, N.; Arivett, B.A.; Adams, M.D.; Bonomo, R.A.; Actis, L.A.; et al. Identification of potential virulence factors in the model strain Acinetobacter baumannii A118. Front. Microbiol. 2019, 10, 1599. [Google Scholar] [CrossRef] [Green Version]
- Rodman Nyah, M.J.; Fung, S.; Nakanouchi, J.; Myers, A.L.; Harris, C.M.; Dang, E.; Fernandez, J.S.; Liu, C.; Mendoza, A.M.; Jimenez, V.; et al. Human Pleural Fluid Elicits Pyruvate and Phenylalanine Metabolism in Acinetobacter baumannii to Enhance Cytotoxicity and Immune Evasion. Front. Microbiol. 2019, 10, 1581. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Don, M.; Merkier, A.K.; Bistue, A.J.; Zorreguieta, A.; Centron, D.; Tolmasky, M.E. Naturally competent Acinetobacter baumannii clinical isolate as a convenient model for genetic studies. J. Clin. Microbiol. 2010, 48, 1488–1490. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, L.A.; Ramage, E.; Weiss, E.J.; Radey, M.; Hayden, H.S.; Held, K.G.; Huse, H.K.; Zurawski, D.V.; Brittnacher, M.J.; Manoil, C. Resources for Genetic and Genomic Analysis of Emerging Pathogen Acinetobacter baumannii. J. Bacteriol. 2015, 197, 2027–2035. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, A.C.; Thompson, M.G.; Black, C.C.; Kessler, J.L.; Clark, L.P.; McQueary, C.N.; Gancz, H.Y.; Corey, B.W.; Moon, J.K.; Si, Y.; et al. AB5075, a Highly Virulent Isolate of Acinetobacter baumannii, as a Model Strain for the Evaluation of Pathogenesis and Antimicrobial Treatments. MBio 2014, 5, e01076-14. [Google Scholar] [CrossRef] [Green Version]
- Bremell, T.; Lange, S.; Svensson, L.; Jennische, E.; Grondahl, K.; Carlsten, H.; Tarkowski, A. Outbreak of spontaneous staphylococcal arthritis and osteitis in mice. Arthritis Rheum. 1990, 33, 1739–1744. [Google Scholar] [CrossRef]
- Diekema, D.J.; Richter, S.S.; Heilmann, K.P.; Dohrn, C.L.; Riahi, F.; Tendolkar, S.; McDanel, J.S.; Doern, G.V. Continued emergence of USA300 methicillin-resistant Staphylococcus aureus in the United States: Results from a nationwide surveillance study. Infect. Control. Hosp. Epidemiol. 2014, 35, 285–292. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Bischoff, M.; Lattar, S.M.; Noto Llana, M.; Pfortner, H.; Niemann, S.; Geraci, J.; Van de Vyver, H.; Fraunholz, M.J.; Cheung, A.L.; et al. Sigma factor SigB Is crucial to mediate Staphylococcus aureus adaptation during chronic infections. PLoS Pathog. 2015, 11, e1004870. [Google Scholar] [CrossRef]
- Nygaard, T.K.; Pallister, K.B.; DuMont, A.L.; DeWald, M.; Watkins, R.L.; Pallister, E.Q.; Malone, C.; Griffith, S.; Horswill, A.R.; Torres, V.J.; et al. Alpha-toxin induces programmed cell death of human T cells, B cells, and monocytes during USA300 infection. PLoS ONE 2012, 7, e36532. [Google Scholar] [CrossRef]
- Moriano, A.; Serra, D.O.; Hoard, A.; Montana, S.; Degrossi, J.; Bonomo, R.A.; Papp-Wallace, K.M.; Ramirez, M.S. Staphylococcus aureus Potentiates the Hemolytic Activity of Burkholderia cepacia Complex (Bcc) Bacteria. Curr. Microbiol. 2021, 78, 1864–1870. [Google Scholar] [CrossRef]
- Quinn, B.; Rodman, N.; Jara, E.; Fernandez, J.S.; Martinez, J.; Traglia, G.M.; Montana, S.; Cantera, V.; Place, K.; Bonomo, R.A.; et al. Human serum albumin alters specific genes that can play a role in survival and persistence in Acinetobacter baumannii. Sci. Rep. 2018, 8, 14741. [Google Scholar] [CrossRef]
- Kondo, I.; Fujise, K. Serotype B staphylococcal bacteriophage singly converting staphylokinase. Infect. Immun. 1977, 18, 266–272. [Google Scholar] [CrossRef] [Green Version]
- Tayabali, A.F.; Nguyen, K.C.; Shwed, P.S.; Crosthwait, J.; Coleman, G.; Seligy, V.L. Comparison of the virulence potential of Acinetobacter strains from clinical and environmental sources. PLoS ONE 2012, 7, e37024. [Google Scholar] [CrossRef] [Green Version]
- CLSI. M100-S30:2020; Performance Standards for Antimicrobial Susceptibility Testing: Thirty Edition Informational Supplement. CLSI Document; Clinical Lab Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Ramirez, M.S.; Traglia, G.M.; Perez, J.F.; Muller, G.L.; Martinez, F.; Golic, A.E.; Mussi, M.A. White and blue light induce reduction in susceptibility to minocycline and tigecycline in Acinetobacter sp. and other bacteria of clinical importance. J. Med. Microbiol. 2015, 64, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Joshi, N.A.; Fass, J.N. Sickle: A Sliding-Window, Adaptive, Quality-Based Trimming Tool for FastQ Files, Version 1.33. 2011. Available online: https://github.com/najoshi/sickle (accessed on 18 February 2022).
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome. Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Novick, R.P.; Jiang, D. The staphylococcal SaeRS system coordinates environmental signals with Agr quorum sensing. Microbiology 2003, 149, 2709–2717. [Google Scholar] [CrossRef] [Green Version]
- Roy, R.; You, R.I.; Lin, M.D.; Lin, N.T. Mutation of the Carboxy-Terminal Processing Protease in Acinetobacter baumannii Affects Motility, Leads to Loss of Membrane Integrity, and Reduces Virulence. Pathogens 2020, 9, 322. [Google Scholar] [CrossRef]
- Clemmer, K.M.; Bonomo, R.A.; Rather, P.N. Genetic analysis of surface motility in Acinetobacter baumannii. Microbiology 2011, 157, 2534–2544. [Google Scholar] [CrossRef] [Green Version]
- Woods, P.W.; Haynes, Z.M.; Mina, E.G.; Marques, C.N.H. Maintenance of S. aureus in Co-culture With P. aeruginosa While Growing as Biofilms. Front. Microbiol. 2018, 9, 3291. [Google Scholar] [CrossRef] [Green Version]
- Goerke, C.; Fluckiger, U.; Steinhuber, A.; Zimmerli, W.; Wolz, C. Impact of the regulatory loci agr, sarA and sae of Staphylococcus aureus on the induction of alpha-toxin during device-related infection resolved by direct quantitative transcript analysis. Mol. Microbiol. 2001, 40, 1439–1447. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, R. Expression and enzymatic properties of a unique recombinant anticoagulant and fibrinolytic enzyme from Acinetobacter baumannii. Int. J. Pharm. Pharm. Sci. 2015, 7, 88–94. [Google Scholar]
- Gaddy, J.A.; Arivett, B.A.; McConnell, M.J.; Lopez-Rojas, R.; Pachon, J.; Actis, L.A. Role of acinetobactin-mediated iron acquisition functions in the interaction of Acinetobacter baumannii strain ATCC 19606T with human lung epithelial cells, Galleria mellonella caterpillars, and mice. Infect. Immun. 2012, 80, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Granchelli, A.M.; Adler, F.R.; Keogh, R.H.; Kartsonaki, C.; Cox, D.R.; Liou, T.G. Microbial interactions in the cystic fibrosis airway. J. Clin. Microbiol. 2018, 56, e00354-18. [Google Scholar] [CrossRef] [Green Version]
- Bansal, E.; Garg, A.; Bhatia, S.; Attri, A.K.; Chander, J. Spectrum of microbial flora in diabetic foot ulcers. Indian J. Pathol. Microbiol. 2008, 51, 204–208. [Google Scholar]
- Bauer, T.T.; Ferrer, R.; Angrill, J.; Schultze-Werninghaus, G.; Torres, A. Ventilator-associated pneumonia: Incidence, risk factors, and microbiology. Semin. Respir. Infec.t 2000, 15, 272–279. [Google Scholar] [CrossRef]
- Stulik, L.; Malafa, S.; Hudcova, J.; Rouha, H.; Henics, B.Z.; Craven, D.E.; Sonnevend, A.M.; Nagy, E. α-Hemolysin activity of methicillin-susceptible Staphylococcus aureus predicts ventilator-associated pneumonia. Am. J. Respir. Crit. Care. Med. 2014, 190, 1139–1148. [Google Scholar] [CrossRef] [Green Version]
- Kahl, B.C. Impact of Staphylococcus aureus on the pathogenesis of chronic cystic fibrosis lung disease. Int. J. Med. Microbiol. 2010, 300, 514–519. [Google Scholar] [CrossRef]
- Rahim, F.; Ullah, F.; Ishfaq, M.; Afridi, A.K.; Rahman, S.U.; Rahman, H. Frequency Of Common Bacteria And Their Antibiotic Sensitivity Pattern In Diabetics Presenting With Foot Ulcer. J. Ayub Med. Coll. Abbottabad 2016, 28, 528–533. [Google Scholar]
- Saipriya, K.; Swathi, C.H.; Ratnakar, K.S.; Sritharan, V. Quorum-sensing system in Acinetobacter baumannii: A potential target for new drug development. J. Appl. Microbiol. 2020, 128, 15–27. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez, J.S.; Tuttobene, M.R.; Montaña, S.; Subils, T.; Cantera, V.; Iriarte, A.; Tuchscherr, L.; Ramirez, M.S. Staphylococcus aureus α-Toxin Effect on Acinetobacter baumannii Behavior. Biology 2022, 11, 570. https://doi.org/10.3390/biology11040570
Fernandez JS, Tuttobene MR, Montaña S, Subils T, Cantera V, Iriarte A, Tuchscherr L, Ramirez MS. Staphylococcus aureus α-Toxin Effect on Acinetobacter baumannii Behavior. Biology. 2022; 11(4):570. https://doi.org/10.3390/biology11040570
Chicago/Turabian StyleFernandez, Jennifer S., Marisel R. Tuttobene, Sabrina Montaña, Tomás Subils, Virginia Cantera, Andrés Iriarte, Lorena Tuchscherr, and Maria Soledad Ramirez. 2022. "Staphylococcus aureus α-Toxin Effect on Acinetobacter baumannii Behavior" Biology 11, no. 4: 570. https://doi.org/10.3390/biology11040570