
Optimisation of Biomass Production and Nutritional Value of Two Marine Diatoms (Bacillariophyceae), Skeletonema costatum and Chaetoceros calcitrans

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
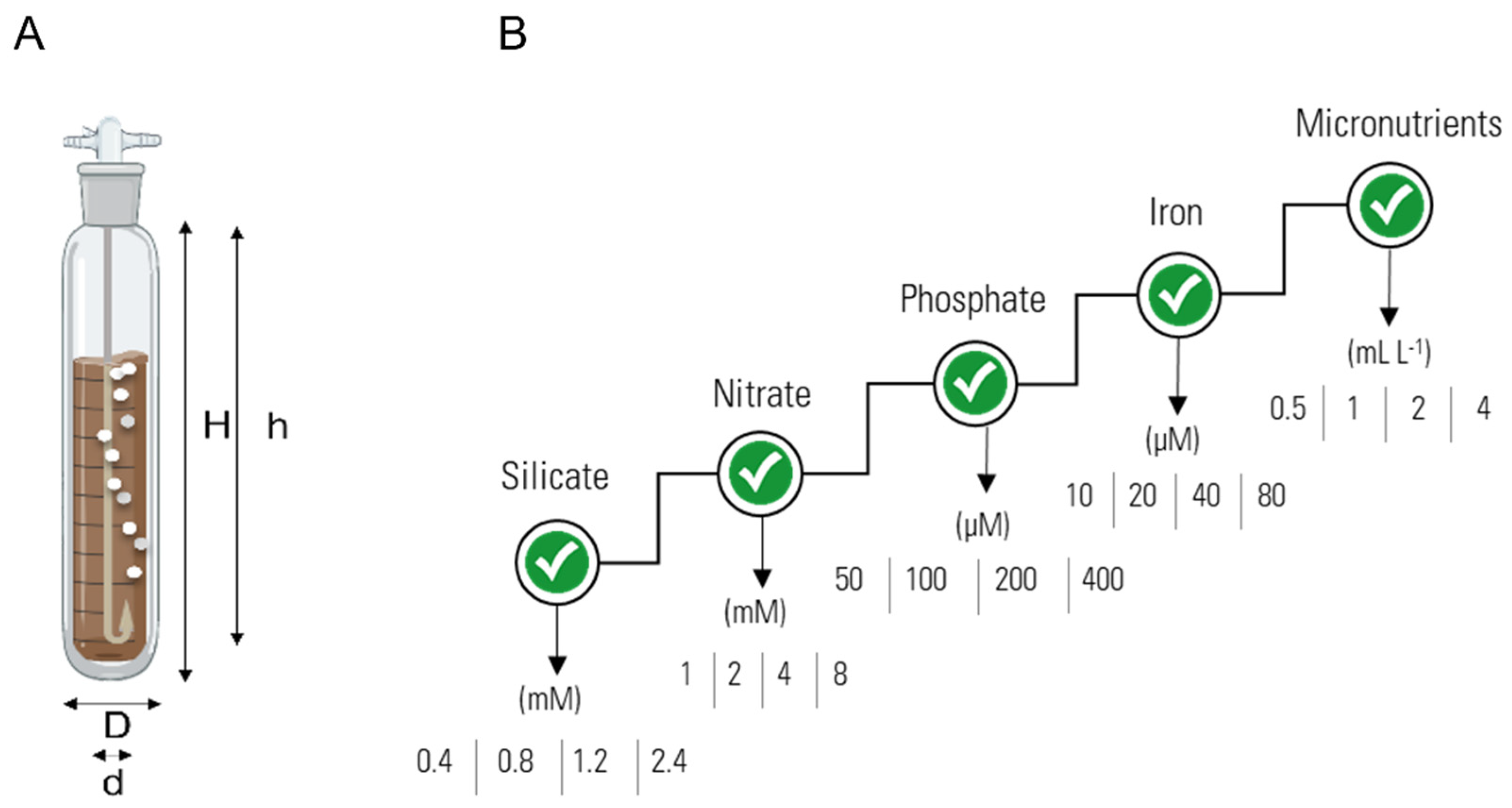
2.1. Culture Medium Optimisation
2.2. Culture Monitoring
2.3. Biochemical Analysis
2.4. Statistical Analysis
3. Results and Discussion
3.1. Culture Optimisation
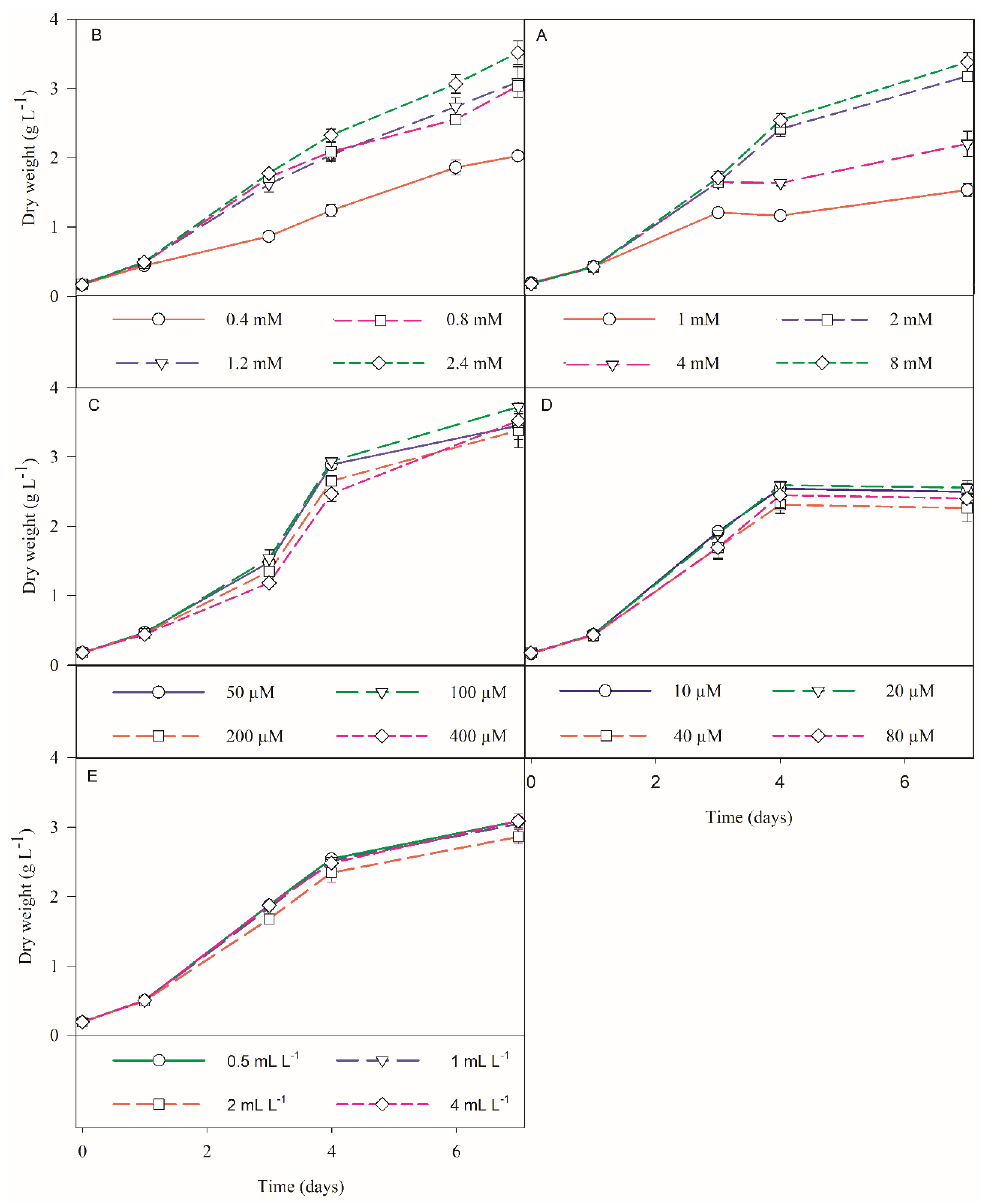
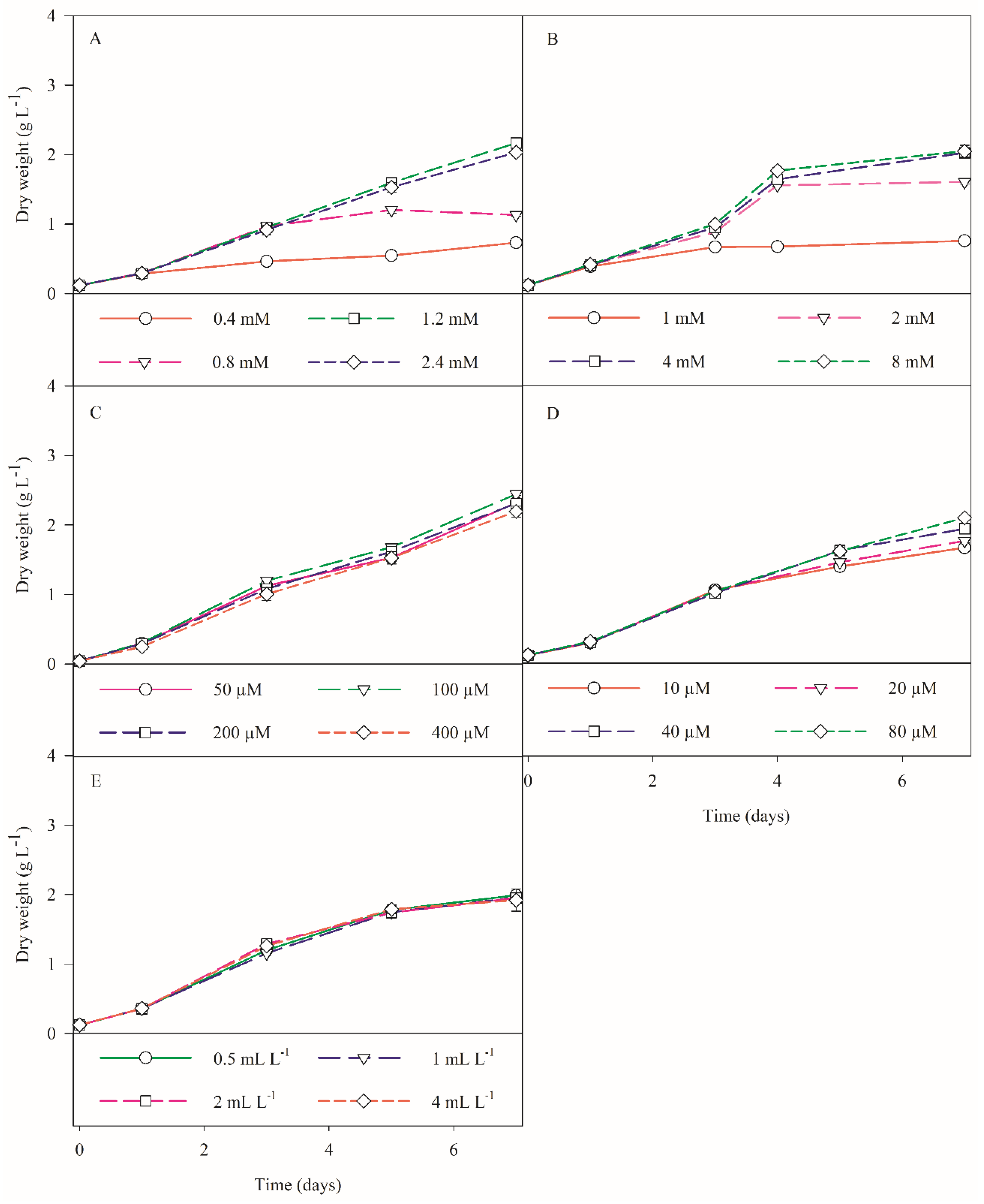
3.1.1. Silicate Trials
3.1.2. Nitrate Trials
3.1.3. Phosphate Trials
3.1.4. Iron Trials
3.1.5. Micronutrients Trials
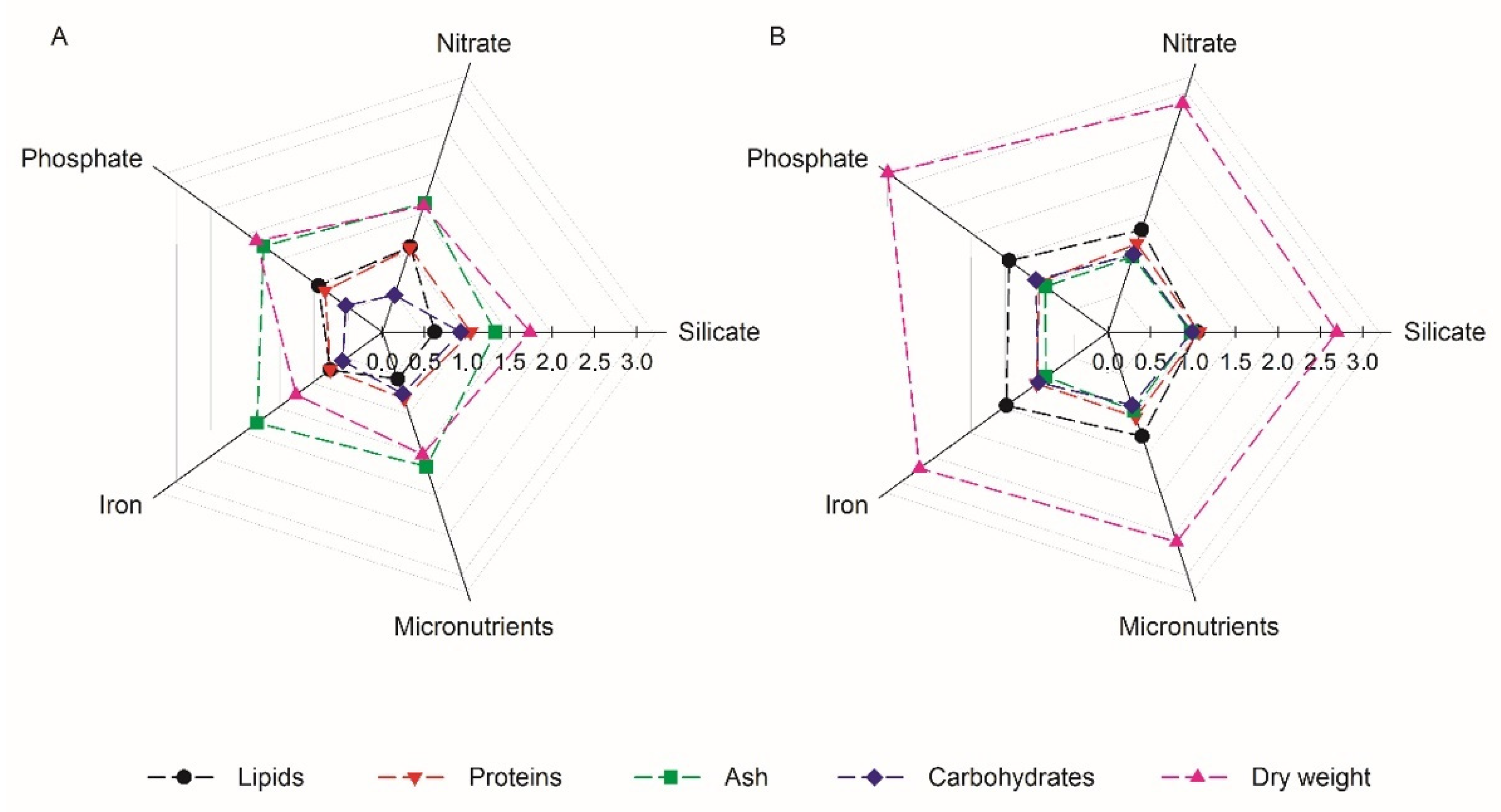
3.2. Biochemical Analysis
3.2.1. Proximal Composition
3.2.2. Fatty Acid Profile of Control and Optimised Conditions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ye, Y.; Huang, Y.; Xia, A.; Fu, Q.; Liao, Q.; Zeng, W.; Zheng, Y.; Zhu, X. Optimizing Culture Conditions for Heterotrophic-Assisted Photoautotrophic Biofilm Growth of Chlorella vulgaris to Simultaneously Improve Microalgae Biomass and Lipid Productivity. Bioresour. Technol. 2018, 270, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Ran, W.; Wang, H.; Liu, Y.; Qi, M.; Xiang, Q.; Yao, C.; Zhang, Y.; Lan, X. Storage of Starch and Lipids in Microalgae: Biosynthesis and Manipulation by Nutrients. Bioresour. Technol. 2019, 291, 121894. [Google Scholar] [CrossRef] [PubMed]
- Kiran, T.; Tiwari, A.; Bhaskar, M.V. A New Novel Solution to Grow Diatom Algae in Large Natural Water Bodies and Its Impact on CO2 Capture and Nutrient Removal. J. Algal Biomass Util. 2015, 6, 2227. [Google Scholar]
- Lebeau, T.; Robert, J.M. Diatom Cultivation and Biotechnologically Relevant Products. Part I: Cultivation at Various Scales. Appl. Microbiol. Biotechnol. 2003, 60, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Bozarth, A.; Maier, U.G.; Zauner, S. Diatoms in Biotechnology: Modern Tools and Applications. Appl. Microbiol. Biotechnol. 2009, 82, 195–201. [Google Scholar] [CrossRef]
- Wang, J.; Seibert, M. Prospects for Commercial Production of Diatoms. Biotechnol. Biofuels 2017, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Zhao, Y.; Sun, Y.; Wu, P.; Zhou, S.; Ren, L. Diatom DNA Barcodes for Forensic Discrimination of Drowning Incidents. FEMS Microbiol. Lett. 2020, 367, fnaa145. [Google Scholar] [CrossRef]
- Mishra, M.; Arukha, A.P.; Bashir, T.; Yadav, D.; Prasad, G.B.K.S. All New Faces of Diatoms: Potential Source of Nanomaterials and Beyond. Front. Microbiol. 2017, 8, 1239. [Google Scholar] [CrossRef] [Green Version]
- Lebeau, T.; Robert, J.M. Diatom Cultivation and Biotechnologically Relevant Products. Part II: Current and Putative Products. Appl. Microbiol. Biotechnol. 2003, 60, 624–632. [Google Scholar] [CrossRef]
- Bhattacharjya, R.; Kiran Marella, T.; Tiwari, A.; Saxena, A.; Kumar Singh, P.; Mishra, B. Bioprospecting of Marine Diatoms Thalassiosira, Skeletonema and Chaetoceros for Lipids and Other Value-Added Products. Bioresour. Technol. 2020, 318, 124073. [Google Scholar] [CrossRef]
- Azmi, K.A.; Arsad, S.; Sari, L.A. The Effect of Commercial Nutrients to Increase the Population of Skeletonema Costatum on Laboratory and Mass Scales. In IOP Conference Series: Earth and Environmental Science; Institute of Physics Publishing: Bristol, UK, 2020; Volume 441. [Google Scholar] [CrossRef]
- Couteau, C.; Coiffard, L. Microalgal Application in Cosmetics. In Microalgae in Health and Disease Prevention; Levine, I.A., Fleurence, J., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 317–323. [Google Scholar] [CrossRef]
- Brown, M.R. Nutritional Value and Uses of Microalgae in Aquaculture. In Avances en Nutrición Acuícola VI. Memorias del VI Simposium Internacional de Nutrición Acuícola; Cruz-Suárez, L.E., Ricque-Marie, D., Tapia-Salazar, M., Gaxiola-Cortés, M.G., Simoes, N., Eds.; CSIRO Marine Research: Hobart, Australia, 2002. [Google Scholar]
- Foo, S.C.; Yusoff, F.M.; Imam, M.U.; Foo, J.B.; Ismail, N.; Azmi, N.H.; Tor, Y.S.; Khong, N.M.H.; Ismail, M. Increased Fucoxanthin in Chaetoceros calcitrans Extract Exacerbates Apoptosis in Liver Cancer Cells via Multiple Targeted Cellular Pathways. Biotechnol. Rep. 2019, 21, e00296. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Wu, M.; Fu, Q.; Li, X.; Xu, J. A Two-Stage Model with Nitrogen and Silicon Limitation Enhances Lipid Productivity and Biodiesel Features of the Marine Bloom-Forming Diatom Skeletonema costatum. Bioresour. Technol. 2019, 289, 121717. [Google Scholar] [CrossRef] [PubMed]
- Cupp, E.E. Marine Plankton Diatoms of the West Coast of North America. In Marine Plankton Diatoms of the West Coast of North America; Sverdrup, H.U., Fleming, R.H., Miller, L.H., ZoBELL, C.E., Eds.; University of California Press: Berkeley, CA, USA, 1962; pp. 1–238. [Google Scholar]
- Kumar, C.S.; Prabu, V.A. Original Research Article Culture of the Phytoplankton Skeletonema costatum, Cleve, 1873. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 129–136. [Google Scholar]
- Li, C.; Zhu, B.; Chen, H.; Liu, Z.; Cui, B.; Wu, J.; Li, B.; Yu, H.; Peng, M. The Relationship between the Skeletonema costatum Red Tide and Environmental Factors in Hongsha Bay of Sanya, South China Sea. J. Coast. Res. 2009, 25, 651–658. [Google Scholar] [CrossRef]
- Gao, G.; Xia, J.; Yu, J.; Zeng, X. Physiological Response of a Red Tide Alga Skeletonema costatum. to Nitrate Enrichment, with Special Reference to Inorganic Carbon Acquisition. Mar. Environ. Res. 2018, 133, 15–23. [Google Scholar] [CrossRef]
- Naviner, M.; Bergé, J.P.; Durand, P.; le Bris, H. Antibacterial Activity of the Marine Diatom Skeletonema costatum against Aquacultural Pathogens. Aquaculture 1999, 174, 15–24. [Google Scholar] [CrossRef]
- Lauritano, C.; Ianora, A. Grand Challenges in Marine Biotechnology: Overview of Recent EU-Funded Projects. In Grand Challenges in Biology and Biotechnology; Springer Science and Business Media B.V.: Berlin/Heidelberg, Germany, 2018; pp. 425–449. [Google Scholar] [CrossRef]
- Brown, M.R. The Amino-Acid and Sugar Composition of 16 Species of Microalgae Used in Mariculture. J. Exp. Mar. Biol. Ecol. 1991, 145, 79–99. [Google Scholar] [CrossRef]
- Monkonsit, S.; Powtongsook, S.; Pavasant, P. Comparison between Airlift Photobioreactor and Bubble Column for Skeletonema costatum Cultivation. Eng. J. 2011, 15, 53–64. [Google Scholar] [CrossRef]
- Putri Lestari, D.; Ekawati, A.W. Dried S. costatum Diatom in Feed of Vaname Shrimp. Life Sci. 2014, 4, 45–49. [Google Scholar]
- van Houcke, J.; Medina, I.; Maehre, H.K.; Cornet, J.; Cardinal, M.; Linssen, J.; Luten, J. The Effect of Algae Diets Skeletonema costatum and Rhodomonas Baltica. on the Biochemical Composition and Sensory Characteristics of Pacific Cupped Oysters Crassostrea gigas. during Land-Based Refinement. Food Res. Int. 2017, 100, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Volkman, J.K.; Jeffrey, S.W.; Nichols, P.D.; Rogers, G.I.; Garland, C.D.; Volkman, J.K. Fatty Acid and Lipid Composition of 10 Species of Microalgae Used in Mariculture; Elsevier: Amsterdam, The Netherlands, 1989; Volume 128. [Google Scholar] [CrossRef]
- Pennarun, A.L.; Prost, C.; Haure, J.; Demaimay, M. Comparison of Two Microalgal Diets. 1. Influence on the Biochemical and Fatty Acid Compositions of Raw Oysters Crassostrea gigas. J. Agric. Food Chem. 2003, 51, 2006–2010. [Google Scholar] [CrossRef] [PubMed]
- Guihéneuf, F.; Mimouni, V.; Ulmann, L.; Tremblin, G. Environmental Factors Affecting Growth and Omega 3 Fatty Acid Composition in Skeletonema costatum. The Influences of Irradiance and Carbon Source. Diatom Res. 2008, 23, 93–103. [Google Scholar] [CrossRef]
- Şirin, S.; Clavero, E.; Salvadó, J. Efficient Harvesting of Chaetoceros calcitrans for Biodiesel Production. Environ. Technol. UK 2015, 36, 1902–1912. [Google Scholar] [CrossRef] [PubMed]
- Krichnavaruk, S.; Loataweesup, W.; Powtongsook, S.; Pavasant, P. Optimal Growth Conditions and the Cultivation of Chaetoceros calcitrans in Airlift Photobioreactor. Chem. Eng. J. 2005, 105, 91–98. [Google Scholar] [CrossRef]
- Kaspar, H.F.; Keys, E.F.; King, N.; Smith, K.F.; Kesarcodi-Watson, A.; Miller, M.R. Continuous Production of Chaetoceros calcitrans in a System Suitable for Commercial Hatcheries. Aquaculture 2014, 420–421, 1–9. [Google Scholar] [CrossRef]
- Pérez, L.; Salgueiro, J.L.; González, J.; Parralejo, A.I.; Maceiras, R.; Cancela, Á. Scaled up from Indoor to Outdoor Cultures of Chaetoceros gracilis and Skeletonema costatum Microalgae for Biomass and Oil Production. Biochem. Eng. J. 2017, 127, 180–187. [Google Scholar] [CrossRef]
- Armstrong, F.A.J. Determination of Nitrate in Water Ultraviolet Spectrophotometry. Anal. Chem. 1963, 35, 1292–1294. [Google Scholar] [CrossRef]
- Smith, J.D.; Milne, J.P. Spectrophotometric Determination of Silicate in Natural Waters by Formation of α-Molybdosilicic Acid and Reduction with a TinIV.-Ascorbic Acid-Oxalic Acid Mixture. Anal. Chim. Acta 1981, 123, 263–270. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; Lavín, P.L.; Lanfer Marquez, U.M.; Aidar, E. Distribution of Intracellular Nitrogen in Marine Microalgae: Calculation of New Nitrogen-to-Protein Conversion Factors. Eur. J. Phycol. 2004, 39, 17–32. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Mozes, A.; Florindo, C.; Polo, C.; Duarte, C.V.; Custádio, L.; Varela, J. Microplate-Based High Throughput Screening Procedure for the Isolation of Lipid-Rich Marine Microalgae. Biotechnol. Biofuels 2011, 4, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepage, G.; Roy, C.C. Improved Recovery of Fatty Acid through Direct Transesterification without Prior Extraction or Purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custódio, L.; Vizetto-Duarte, C.; Polo, C.; Rešek, E.; Aschwin, E.; Varela, J. Polyunsaturated Fatty Acids of Marine Macroalgae: Potential for Nutritional and Pharmaceutical Applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimi, E.; Salarzadeh, A. The Effect of Temperature and Salinity on the Growth of Skeletonema costatum and Chlorella capsulata in Vitro. Int. J. Life Sci. 2016, 10, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Ee Hew, W.; Khatoon, H.; Shariff, M.; Yusoff, F.M. Chaetoceros calcitrans and Nannochloropsis oculata Cultured Outdoors and under Laboratory Conditions. Afr. J. Biotechnol. 2011, 10, 1375–1383. [Google Scholar] [CrossRef]
- Vaulot, D.; Olson, R.J.; Merkel, S.; Chisholm, S.W. Cell-Cycle Response to Nutrient Starvation in Two Phytoplankton Species, Thalassiosira weissflogii and Hymenomonas arterae. Mar. Biol. 1987, 95, 625–630. [Google Scholar] [CrossRef]
- Brzezinski, M.A.; Villareal, T.A.; Lipschultz, F. Silica Production and the Contribution of Diatoms to New and Primary Production in the Central North Pacific. Mar. Ecol. Prog. Ser. 1998, 167, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Heintze, C.; Formanek, P.; Pohl, D.; Hauptstein, J.; Rellinghaus, B.; Kröger, N. An Intimate View into the Silica Deposition Vesicles of Diatoms. BMC Mater. 2020, 2, 11. [Google Scholar] [CrossRef]
- Abate, R.; Song, S.; Patil, V.; Chen, C.; Liang, J.; Sun, L.; Li, X.; Huang, B.; Gao, Y. Enhancing the Production of a Marine Diatom Skeletonema costatum. with Low-Frequency Ultrasonic Irradiation. J. Appl. Phycol. 2020, 32, 3711–3722. [Google Scholar] [CrossRef]
- Gleich, S.J.; Plough, L.V.; Glibert, P.M. Photosynthetic Efficiency and Nutrient Physiology of the Diatom Thalassiosira pseudonana at Three Growth Temperatures. Mar. Biol. 2020, 167, 124. [Google Scholar] [CrossRef]
- Corzo, A.; Morillo, J.A.; Rodríguez, S. Production of Transparent Exopolymer Particles TEP. in Cultures of Chaetoceros calcitrans under Nitrogen Limitation. Aquat. Microb. Ecol. 2000, 23, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Tantanasarit, C.; Englande, A.J.; Babel, S. Nitrogen, Phosphorus and Silicon Uptake Kinetics by Marine Diatom Chaetoceros calcitrans under High Nutrient Concentrations. J. Exp. Mar. Biol. Ecol. 2013, 446, 67–75. [Google Scholar] [CrossRef]
- Diaz, J.; Ingall, E.; Benitez-Nelson, C.; Paterson, D.; de Jonge, M.D.; McNulty, I.; Brandes, J.A. Marine Polyphosphate: A Key Player in Geologic Phosphorus Sequestration. Science 2008, 320, 652–655. [Google Scholar] [CrossRef]
- Sanz-Luque, E.; Bhaya, D.; Grossman, A.R. Polyphosphate: A Multifunctional Metabolite in Cyanobacteria and Algae. Front. Plant Sci. 2020, 11, 938. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Hahn, H.H.; Hoffmann, E. Effects of Solution Conditions on the Precipitation of Phosphate for Recovery: A Thermodynamic Evaluation. Chemosphere 2002, 48, 1029–1034. [Google Scholar] [CrossRef]
- Solovchenko, A.E.; Ismagulova, T.T.; Lukyanov, A.A.; Vasilieva, S.G.; Konyukhov, I.V.; Pogosyan, S.I.; Lobakova, E.S.; Gorelova, O.A. Luxury Phosphorus Uptake in Microalgae. J. Appl. Phycol. 2019, 31, 2755–2770. [Google Scholar] [CrossRef]
- Rivero-Rodríguez, S.; Beaumont, A.R.; Lora-Vilchis, M.C. The Effect of Microalgal Diets on Growth, Biochemical Composition, and Fatty Acid Profile of Crassostrea Corteziensis Hertlein. Juveniles. Aquaculture 2007, 263, 199–210. [Google Scholar] [CrossRef]
- Lampe, R.H.; Mann, E.L.; Cohen, N.R.; Till, C.P.; Thamatrakoln, K.; Brzezinski, M.A.; Bruland, K.W.; Twining, B.S.; Marchetti, A. Different Iron Storage Strategies among Bloom-Forming Diatoms. Proc. Natl. Acad. Sci. USA 2018, 115, E12275–E12284. [Google Scholar] [CrossRef] [Green Version]
- Anu, P.R.; Bijoy Nandan, S.; Jayachandran, P.R.; Don Xavier, N.D. Toxicity Effects of Copper on the Marine Diatom, Chaetoceros calcitrans. Reg. Stud. Mar. Sci. 2016, 8, 498–504. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, J.; Tan, L.; Chen, X. Toxic Effects of Nano-ZnO on Marine Microalgae Skeletonema costatum: Attention to the Accumulation of Intracellular Zn. Aquat. Toxicol. 2016, 178, 158–164. [Google Scholar] [CrossRef]
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar] [CrossRef]
- Kudaibergenov, A.K.; Khajiyeva, L.A. Modelling of Resonance and Stability of Drill String Nonlinear Dynamics. Aquaculture 1987, 60, 231–241. [Google Scholar]
- Raven, B.Y.J.A. The Role of Vacuoles. New Phytol. 1987, 106, 357–422. [Google Scholar] [CrossRef]
- Hildebrand, M.; Davis, A.K.; Smith, S.R.; Traller, J.C.; Abbriano, R. The Place of Diatoms in the Biofuels Industry. Biofuels 2012, 3, 221–240. [Google Scholar] [CrossRef] [Green Version]
- Maia, I.B.; Carneiro, M.; Magina, T.; Malcata, F.X.; Otero, A.; Navalho, J.; Varela, J.; Pereira, H. Diel Biochemical and Photosynthetic Monitorization of Skeletonema costatum and Phaeodactylum tricornutum Grown in Outdoor Pilot-Scale Flat Panel Photobioreactors. J. Biotechnol. 2022, 343, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, X.; Cao, X.; Yu, Z. Responses of Photosynthetic Characters of Skeletonema costatum to Different Nutrient Conditions. J. Plankton Res. 2013, 35, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Nedbal, L.; Ebenhöh, O. Modelling Phosphorus Uptake in Microalgae. Biochem. Soc. Trans. 2018, 46, 483–490. [Google Scholar] [CrossRef]
- Velasco, L.A.; Carrera, S.; Barros, J. Isolation, Culture and Evaluation of Chaetoceros muelleri from the Caribbean as Food for the Native Scallops, Argopecten nucleus and Nodipecten nodosus. Lat. Am. J. Aquat. Res. 2016, 44, 557–568. [Google Scholar] [CrossRef]
- Granum, E.; Kirkvold, S.; Myklestad, S.M. Cellular and Extracellular Production of Carbohydrates and Amino Acids by the Marine Diatom Skeletonema costatum: Diel Variations and Effects of N Depletion. Mar. Ecol. Prog. Ser. 2002, 242, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Schmid, A.-M.M.; Borowitzka, M.A.; Volcani, B.E. Morphogenesis and Biochemistry of Diatom Cell Walls. In Cytomorphogenesis in Plants; Kiermayer, O., Ed.; Springer: Berlin/Heidelberg, Germany, 1981; pp. 63–97. [Google Scholar]
- Gautam, S.; Kaur, M.; Rohilla, A.K. The Role of Physical Chemical Parameters on Diatoms Growth—A Review. Plant Arch. 2019, 19, 570–573. [Google Scholar]
- Fernández-Reiriz, M.J.; Perez-Camacho, A.; Ferreiro, M.J.; Blanco, J.; Planas, M.; Campos, M.J.; Labarta, U. Biomass Production and Variation in the Biochemical Profile Total Protein, Carbohydrates, RNA, Lipids and Fatty Acids of Seven Species of Marine Microalgae. Aquaculture 1989, 83, 17–37. [Google Scholar] [CrossRef]
- Delaunay, F.; Marty, Y.; Moal, J.; Samain, J.F. The Effect of Monospecific Algal Diets on Growth and Fatty Acid Composition of Pecten maximus L. Larvae. J. Exp. Mar. Biol. Ecol. 1993, 173, 163–179. [Google Scholar] [CrossRef]
- Prartono, T.; Kawaroe, M.; Katili, V. Fatty Acid Composition of Three Diatom Species Skeletonema costatum, Thalassiosira sp. and Chaetoceros Gracilis. Int. J. Environ. Bioener. 2013, 6, 28–43. [Google Scholar]
- Rohit, M.V.; Mohan, S.V. Quantum Yield and Fatty Acid Profile Variations with Nutritional Mode during Microalgae Cultivation. Front. Bioeng. Biotechnol. 2018, 6, 111. [Google Scholar] [CrossRef]
- Méndez-Martínez, Y.; García-Guerrero, M.U.; Lora-Vilchis, M.C.; Martínez-Córdova, L.R.; Arcos-Ortega, F.G.; Alpuche, J.J.; Cortés-Jacinto, E. Nutritional Effect of Artemia nauplii Enriched with Tetraselmis suecica and Chaetoceros calcitrans Microalgae on Growth and Survival on the River Prawn Macrobrachium americanum Larvae. Aquac. Int. 2018, 26, 1001–1015. [Google Scholar] [CrossRef]
- Remize, M.; Brunel, Y.; Silva, J.L.; Berthon, J.Y.; Filaire, E. Microalgae n − 3 PUFAs Production and Use in Food and Feed Industries. Mar. Drugs 2021, 19, 113. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.K.; Jiménez, C.; Wulff, A. Nutraceutical Fatty Acid Production in Marine Microalgae and Cyanobacteria. In Nutraceutical Fatty Acids from Oleaginous Microalgae; Patel, A.K., Matsakas, L., Eds.; Scrivener Publishing LLC: Berlin/Heidelberg, Germany, 2020; pp. 23–76. [Google Scholar] [CrossRef]
- Pratiwy, F.M.; Pratiwi, D.Y. The Potentiality of Microalgae as a Source of DHA and EPA for Aquaculture Feed: A Review. Int. J. Fish. Aquat. Stud. 2020, 8, 39–41. [Google Scholar]
- Srinuanpan, S.; Cheirsilp, B.; Prasertsan, P.; Asano, Y.; Kato, Y. Strategies to Increase the Potential Use of Oleaginous Microalgae as Biodiesel Feedstocks: Nutrient Starvations and Cost-Effective Harvesting Process. Renew. Energy 2018, 122, 507–516. [Google Scholar] [CrossRef]
- Deshmukh, S.; Kumar, R.; Bala, K. Microalgae Biodiesel: A Review on Oil Extraction, Fatty Acid Composition, Properties and Effect on Engine Performance and Emissions. Fuel Process. Technol. 2019, 191, 232–247. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macronutrients | Metals | Vitamins |
|---|---|---|
| 2 M N | 20 Fe (mM) | 10.4 Thiamine (µM) |
| 100 mM P | 1 Zn (mM) | 2 Biotin (µM) |
| 1 Mo (mM) | 2 B12 (µM) | |
| 1 Mn (mM) | ||
| 2 Mg (mM) | ||
| 0.1 Cu (mM) | ||
| 0.1 Co (mM) |
| Trial | Treatments | Silicate (mM) | Nitrate (mM) | Phosphate (µM) | Iron (µM) | Micronutrients (mL L−1) |
|---|---|---|---|---|---|---|
| Silicate | Tested nutrients | 0.4|0.8|1.2|2.4 | 4 | 200 | 40 | 2 |
| Optimised combination | 2.4 | 4 | 200 | 40 | 2 | |
| Nitrate | Tested nutrients | 2.4 | 1|2|4|8 | 200 | 40 | 2 |
| Optimised combination | 2.4 | 4 | 200 | 40 | 2 | |
| Phosphate | Tested nutrients | 2.4 | 4 | 50|100|200|400 | 40 | 2 |
| Optimised combination | 2.4 | 4 | 100 | 40 | 2 | |
| Iron | Tested nutrients | 2.4 | 4 | 100 | 10|20|40|80 | 2 |
| Optimised combination | 2.4 | 4 | 100 | 20 | 2 | |
| Micronutrients | Tested nutrients | 2.4 | 4 | 100 | 20 | 0.5|1|2|4 |
| Final optimised combination | 2.4 | 4 | 100 | 20 | 0.5 |
| Trial | Treatments | Silicate (mM) | Nitrate (mM) | Phosphate (µM) | Iron (µM) | Micronutrients (mL L−1) |
|---|---|---|---|---|---|---|
| Silicate | Tested nutrients | 0.4|0.8|1.2|2.4 | 4 | 200 | 40 | 2 |
| Optimised combination | 1.2 | 4 | 200 | 40 | 2 | |
| Nitrate | Tested nutrients | 1.2 | 1|2|4|8 | 200 | 40 | 2 |
| Optimised combination | 1.2 | 4 | 200 | 40 | 2 | |
| Phosphate | Tested nutrients | 1.2 | 4 | 50|100|200|400 | 40 | 2 |
| Optimised combination | 1.2 | 4 | 100 | 40 | 2 | |
| Iron | Tested nutrients | 1.2 | 4 | 100 | 10|20|40|80 | 2 |
| Optimised combination | 1.2 | 4 | 100 | 80 | 2 | |
| Micronutrients | Tested nutrients | 1.2 | 4 | 100 | 80 | 0.5|1|2|4 |
| Final optimised combination | 1.2 | 4 | 100 | 80 | 0.5 |
| Fatty Acids (%) | Skeletonema costatum (C) | Skeletonema costatum (O) | Chaetoceros calcitrans (C) | Chaetoceros calcitrans (O) |
|---|---|---|---|---|
| (C14:0) | 31.4 ± 3.8 | 17.3 ± 0.3 | 15.8 ± 1.2 | 13.1 ± 1.1 |
| (C16:0) | 7.3 ± 1.4 | 4.9 ± 0.0 | 32.9 ± 1.1 | 20.6 ± 1.6 |
| (C18:0) | n.d. | n.d. | 1.6 ± 0.4 | 1.4 ± 0.1 |
| ∑SFA | 36.3 ± 7.4 a | 18.9 ± 2.8 b | 50.3 ± 0.5 a | 34.4 ± 0.8 b |
| (C16:1) | 40.9 ± 2.9 | 23.6 ± 1.4 | 27.2 ± 1.0 | 32.9 ± 0.7 |
| (C18:1) | 2.6 ± 0.5 | 1.3 ± 0.0 | 4.6 ± 0.6 | 2.8 ± 0.5 |
| ∑MUFA | 42.2 ± 4.3 a | 24.9 ± 1.4 b | 31.8 ± 0.6 b | 35.7 ± 0.6 a |
| (C18:4) | 0.6 ± 0.3 | 2.2 ± 0.1 | n.d. | n.d. |
| (C16:4) | 6.6 ± 2.4 | 20.1 ± 0.5 | n.d. | n.d. |
| (C16:3) | 3.2 ± 1.3 | 6.5 ± 0.1 | 6.1 ± 0.5 | 9.2± 2.6 |
| (C20:5) | 8.9 ± 5.6 | 22.2 ± 1.3 | 10.5 ± 0.6 | 18.8 ± 2.3 |
| (C22:6) | 2.0 ± 1.2 | 5.2 ± 0.3 | 1.3 ± 0.2 | 1.9 ± 0.2 |
| ∑PUFA | 21.6 ± 11.1 b | 56.2 ± 1.4 a | 17.7 ± 0.3 b | 29.9 ± 0.5 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bastos, C.R.V.; Maia, I.B.; Pereira, H.; Navalho, J.; Varela, J.C.S. Optimisation of Biomass Production and Nutritional Value of Two Marine Diatoms (Bacillariophyceae), Skeletonema costatum and Chaetoceros calcitrans. Biology 2022, 11, 594. https://doi.org/10.3390/biology11040594
Bastos CRV, Maia IB, Pereira H, Navalho J, Varela JCS. Optimisation of Biomass Production and Nutritional Value of Two Marine Diatoms (Bacillariophyceae), Skeletonema costatum and Chaetoceros calcitrans. Biology. 2022; 11(4):594. https://doi.org/10.3390/biology11040594
Chicago/Turabian StyleBastos, Carolina R. V., Inês B. Maia, Hugo Pereira, João Navalho, and João C. S. Varela. 2022. "Optimisation of Biomass Production and Nutritional Value of Two Marine Diatoms (Bacillariophyceae), Skeletonema costatum and Chaetoceros calcitrans" Biology 11, no. 4: 594. https://doi.org/10.3390/biology11040594
APA StyleBastos, C. R. V., Maia, I. B., Pereira, H., Navalho, J., & Varela, J. C. S. (2022). Optimisation of Biomass Production and Nutritional Value of Two Marine Diatoms (Bacillariophyceae), Skeletonema costatum and Chaetoceros calcitrans. Biology, 11(4), 594. https://doi.org/10.3390/biology11040594