
Fetal Fascial Reinforcement Development: From “a White Tablet” to a Sculpted Precise Organization by Movement

 ,
, 
 ,
,  ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
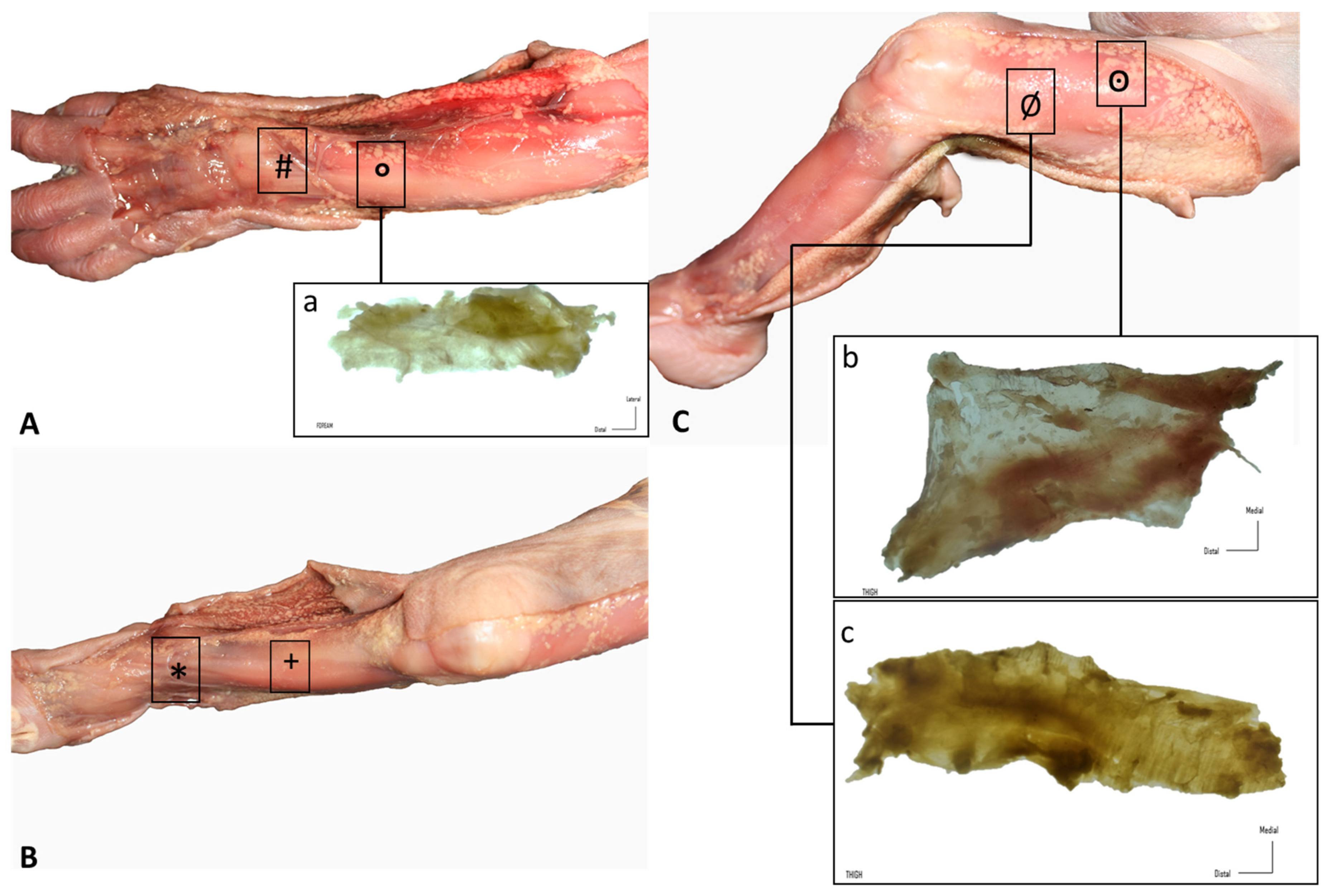
2.1. Gross Anatomy
2.2. Histological Study
2.3. Immunocytochemistry to Detect Hyaluronic Acid-Binding Protein (HABP)
2.4. Immunocytochemistry to Detect Collagen Types I and III
3. Results
3.1. Gross Anatomy
3.2. Histological Study
- −
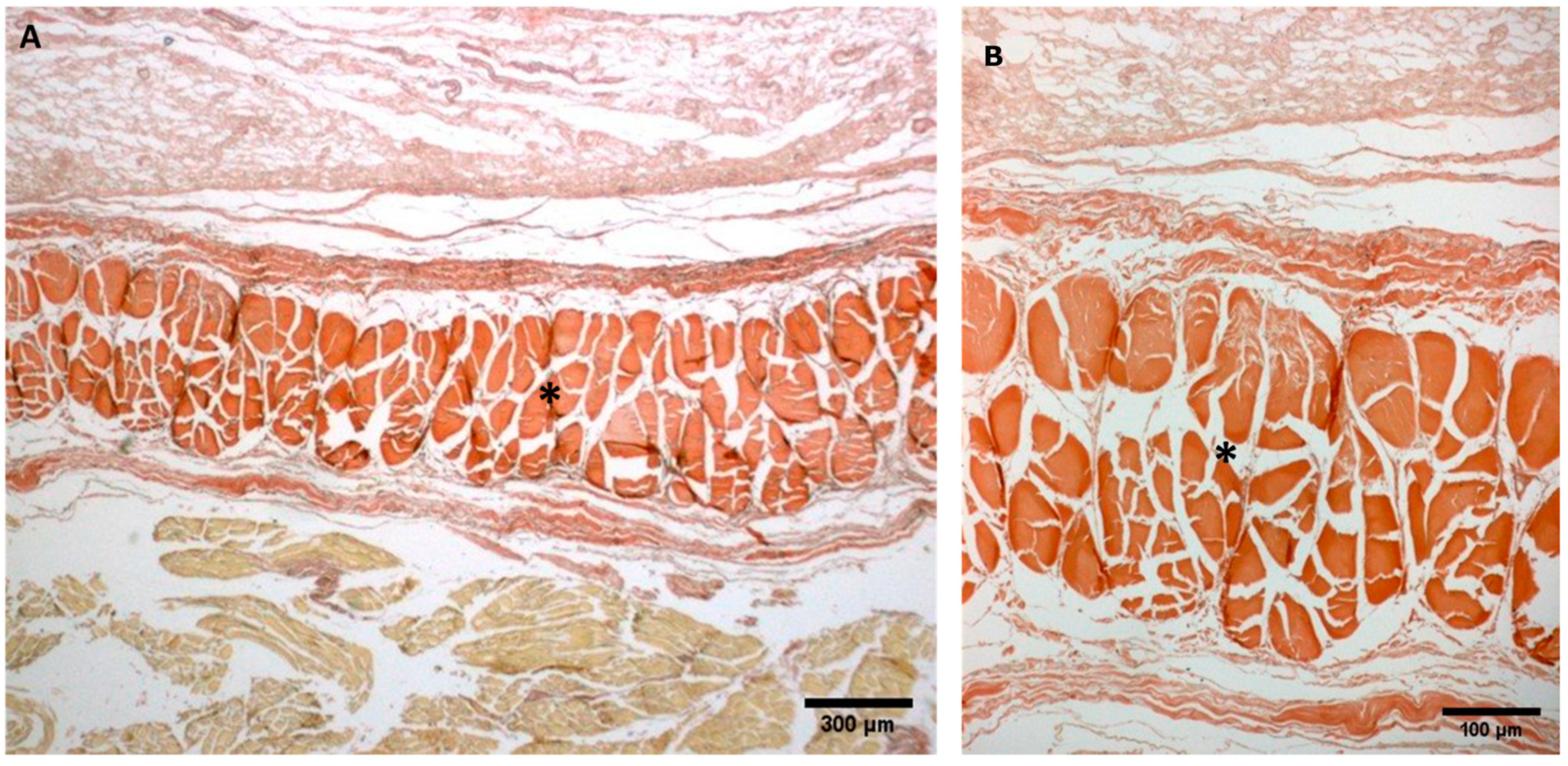
- Week 24: In all samples, a singular layer of mesenchymal irregular connective tissue was observed. The deep fascia was clearly identified (Figure 2A, Figure 3A, Figure 4A, Figure 5A and Figure 6A) but it was not organized in layers and the fascial thickness is huge in all samples, 531.6 ± 178.4 µm, except for the extensor retinaculum of the upper limb 112.40 ± 6.1 µm (Figure 7A). In the deep fascia, a huge cellularity without clear organization was evident. Hematoxylin-eosin demonstrated the presence of a high number of mesenchymal cells, isolated groups of adipocytes, blood vessels and neural structures within this layer.
- −
- Week 27: In all samples, the deep fascia was composed of a huge cellularity without clear organization in layers (Figure 2B, Figure 3B, Figure 4B and Figure 6B). No sublayers were recognized, and in some cases, small aggregates of adipose tissue were found within these structures. The mean thickness was 319.8 ± 51.83 µm for all fascial structures, except for the extensor retinaculum of the upper limb, 169.7 ± 10.30 µm (Figure 7B).
- −
- Week 29: In all samples, the deep fascia started to acquire an organization showing dense connective tissue (Figure 2C, Figure 3C, Figure 4C, Figure 5C and Figure 6C). Some of the sketches of the first layers began to show. The same histological features as those seen at 27 weeks were found, as described in samples from previous weeks. The mean thickness was 229.8 ± 87.79 µm for all fascial structures.
- −
- Week 36: All samples started to show a well-differentiated deep fascia-like structure of 2–3 layers of dense regular connective tissue with areolar tissue in between (Figure 2D, Figure 3D, Figure 4D and Figure 6D). At this age, organization was seen in sublayers but the fibers were organized in one direction. The mean thickness was 239.3 ± 59.15 µm for all fascial structures.
- −
- Week 38: As was seen at 36 weeks, the deep fascia was significantly clear, with a well-defined organization, with 2–3 layers of dense regular connective tissue with areolar tissue in between (Figure 2E, Figure 3E, Figure 4E, Figure 5C and Figure 6E). All samples showed a structure organized in different layers with different directions (transversal and longitudinal). The middle layer was the most prominent and consisted of compact bundles of collagen fibers, while the outermost layers showed a woven-like and thin pattern. This was evident in the deep fasciae of the anterior compartment of the thigh and the leg, but also in the iliotibial tract and in the extensor retinaculum of the ankle, while in the wrist the organization in layers has yet to be constituted. The mean thickness was 219.6 ± 86.1 µm for all fascial structures, except for the ex-tensor retinaculum of the upper limb, 334.6 ± 44.74 µm (Figure 7C).
- −
- Week 40: The deep fascia shows a clear stratification. All samples showed a well-differentiated deep fascia-like structure of 2–3 layers of dense regular connective tissue with areolar connective tissue in between (Figure 2F, Figure 3F, Figure 4F, Figure 5D and Figure 6F). The connective tissue of the thigh region showed the most organized pattern, with the middle layer consisting of several non-aligned dense connective tissue bundles, all in the same direction. The mean thickness was 176.8 ± 58.95 µm for all fascial structures, except for the extensor retinaculum of the upper limb 545.4 ± 22.33 µm (Figure 7D).
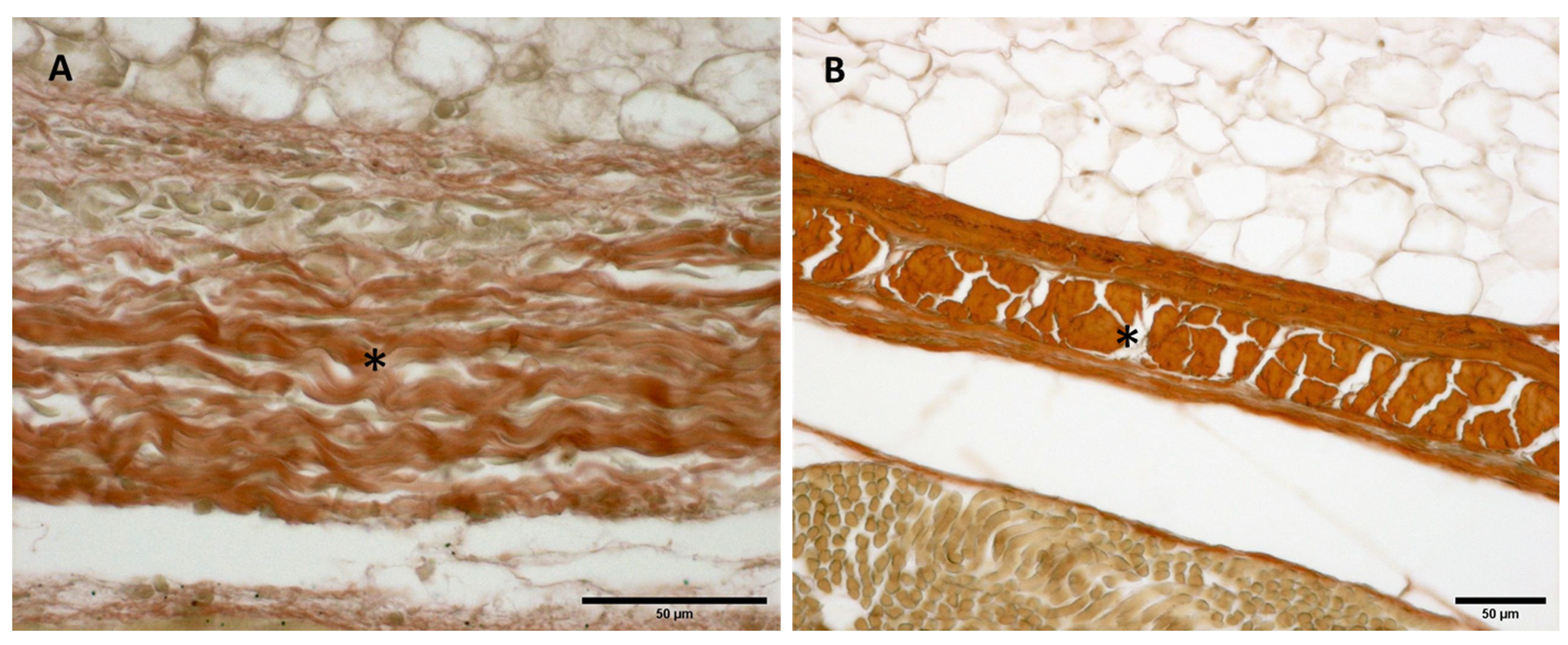
3.2.1. Hyaluronan and Elastic Fibers
3.2.2. Collagen I and III
4. Discussion
Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
Appendix A
References
- Sinhorim, L.; Amorim, M.D.S.; Ortiz, M.E.; Bittencourt, E.B.; Bianco, G.; da Silva, F.C.; Horewicz, V.V.; Schleip, R.; Reed, W.R.; Mazzardo-Martins, L.; et al. Potential Nociceptive Role of the Thoracolumbar Fascia: A Scope Review Involving In Vivo and Ex Vivo Studies. J. Clin. Med. 2021, 24, 4342. [Google Scholar] [CrossRef] [PubMed]
- Amir, A.; Kim, S.; Stecco, A.; Jankowski, M.P.; Raghavan, P. Hyaluronan homeostasis and its role in pain and muscle stiffness. PM R 2022. [Google Scholar] [CrossRef] [PubMed]
- Roch, M.; Gaudreault, N.; Cyr, M.P.; Venne, G.; Bureau, N.J.; Morin, M. The Female Pelvic Floor Fascia Anatomy: A Systematic Search and Review. Life 2021, 30, 900. [Google Scholar] [CrossRef] [PubMed]
- Vesalio, A. De Humani Corporis Fabbrica: Basel, Ex officina Joannis Oporini 1543; Octavo: Palo Alto, CA, USA, 1998. [Google Scholar]
- Leardini, A.; O’Connor, J.J. A model for lever-arm length calculation of the flexor and extensor muscles at the ankle. Gait Posture 2002, 15, 220–229. [Google Scholar] [CrossRef]
- Umidon, M. Architecture, Topography and Morphogenesis of The Peroneal Retinacula and the Lateral Annular Ligament of The Tarsus in Man. Chir Organi Mov. 1963, 52, 305–317. [Google Scholar]
- Demondion, X.; Canella, C.; Moraux, A.; Cohen, M.; Bry, R.; Cotten, A. Retinacular disorders of the ankle and foot. Semin. Musculoskelet. Radiol. 2010, 14, 281–291. [Google Scholar] [CrossRef]
- McCarthy, C.L. Ultrasound of Normal and Injured Ligaments and Retinacula of the Hand. Semin. Musculoskelet. Radiol. 2020, 24, 83–100. [Google Scholar] [CrossRef]
- Viladot, A.; Lorenzo, J.C.; Salazar, J.; Rodrıguez, A. The subtalar joint: Embryology and morphology. Foot Ankle 1984, 5, 54–66. [Google Scholar] [CrossRef]
- Klein, D.M.; Katzman, B.M.; Mesa, J.A.; Lipton, J.F.; Caligiuri, D.A. Histology of the extensor retinaculum of the wrist and the ankle. J. Hand Surg. 1999, 24, 799–802. [Google Scholar] [CrossRef]
- Stecco, A.; Stecco, C.; Macchi, V.; Porzionato, A.; Ferraro, C.; Masiero, S.; De Caro, R. RMI study and clinical correlations of ankle retinacula damage and outcomes of ankle sprain. Surg. Radiol. Anat. 2011, 33, 881–890. [Google Scholar] [CrossRef]
- Blasi, M.; Blasi, J.; Domingo, T.; Pérez-Bellmunt, A.; Miguel-Pérez, M. Anatomical and histological study of human deep fasciae development. Surg. Radiol. Anat. 2015, 37, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Suzuki, M.; Cho, K.H.; Murakami, G.; Cho, B.H.; Ide, Y. CD34-positive developing vessels and other structures in human fetuses: An immunohistochemical study. Surg. Radiol. Anat. 2011, 33, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.H.; Jang, H.S.; Abe, H.; Yamamoto, M.; Murakami, G.; Shibata, S. Fetal Development of Fasciae around the Arm and Thigh Muscles: A Study Using Late Stage Fetuses. Anat. Rec. 2018, 301, 1235–1243. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.H.; Jin, Z.W.; Abe, H.; Wilting, J.; Murakami, G.; Rodríguez-Vázquez, J.F. Tensor fasciae latae muscle in human embryos and foetuses with special reference to its contribution to the development of the iliotibial tract. Folia Morphol. (Warsz) 2018, 77, 703–710. [Google Scholar] [PubMed] [Green Version]
- Young, B.; Woodford, P.; O’Dowd, G. Wheater’s Functional Histology, A Text and Colour Atlas, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Pirri, C.; Fede, C.; Petrelli, L.; Guidolin, D.; Fan, C.; De Caro, R.; Stecco, C. An anatomical comparison of the fasciae of the thigh: A macroscopic, microscopic and ultrasound imaging study. J. Anat. 2020, 238, 999–1009. [Google Scholar] [CrossRef]
- Pirri, C.; Fede, C.; Petrelli, L.; Guidolin, D.; Fan, C.; De Caro, R.; Stecco, C. Elastic Fibres in the subcutaneous tissue: Is there a difference between superficial and muscular fascia? A cadaver study. Ski. Res. Technol. 2021, 28, 21–27. [Google Scholar] [CrossRef]
- Feldman, H.N. Developmental-Behavioural Pediatrics. In Atlas of Pediatric Physical Diagnosis, 4th ed.; Bj, Z., Hw, D., Eds.; Mosby: St. Louis, MI, USA, 2002; pp. 58–86. [Google Scholar]
- Verbruggen, S.W.; Loo, J.H.; Hayat, T.T.; Hajnal, J.V.; Rutherford, M.A.; Phillips, A.T.; Nowlan, N.C. Modeling the biomechanics of fetal movements. Biomech. Model. Mechanobiol. 2016, 15, 995–1004. [Google Scholar] [CrossRef] [Green Version]
- Katz, K.; Mashiach, R.; Bar On, A.; Merlob, P.; Soudry, M.; Meizner, I. Normal range of fetal knee movements. J. Pediatr. Orthop. 1999, 19, 739–741. [Google Scholar] [CrossRef]
- Shaw, H.M.; Vazquez, O.T.; McGonagle, D.; Bydder, G.; Santer, R.M.; Benjamin, M. Development of the human Achilles tendon enthesis organ. J. Anat. 2008, 213, 718–724. [Google Scholar] [CrossRef]
- Fan, C.; Pirri, C.; Fede, C.; Guidolin, D.; Biz, C.; Petrelli, L.; Porzionato, A.; Macchi, V.; De Caro, R.; Stecco, C. Age-Related Alterations of Hyaluronan and Collagen in Extracellular Matrix of the Muscle Spindles. J. Clin. Med. 2021, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Pratt, R.L. Hyaluronan and the Fascial Frontier. Int. J. Mol. Sci. 2021, 22, 6845. [Google Scholar] [CrossRef] [PubMed]
- Stecco, C.; Macchi, V.; Porzionato, A.; Morra, A.; Parenti, A.; Stecco, A.; Delmas, V.; De Caro, R. The ankle retinacula: Morphological evidence of the proprioceptive role of the fascial system. Cells Tissues Organs 2010, 192, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Numkarunarunrote, N.; Malik, A.; Aguiar, R.O.; Trudell, D.J.; Resnick, D. Retinacula of the foot and ankle: MRI with anatomic correlation in cadavers. Am. J. Roentgenol. 2007, 188, 348–354. [Google Scholar] [CrossRef]
- Blottner, D.; Huang, Y.; Trautmann, G.; Sun, L. The fascia: Continuum linking bone and myofascial bag for global and local body movement control on Earth and in Space. A scoping review. Reach 2019, 14, 100030. [Google Scholar] [CrossRef]
- Abu-Hijleh, M.F.; Harris, P.F. Deep fascia on the dorsum of the ankle and foot: Extensor retinacula revisited. Clin. Anat. 2007, 20, 186–195. [Google Scholar] [CrossRef]
- Sarrafian, S.K. Anatomy of the Foot and Ankle: Descriptive, Topographic, Functional, 1st ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1983; pp. 157–175. [Google Scholar]
- Moore, K.L.; Dalley, A.F., II. Clinically Oriented Anatomy, 4th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1999; pp. 594–595. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weeks | Upper Limb (Extensor Retinaculum) | Upper Limb (Antebrachial Fascia) | Lower Limb (Extensor Retinaculum) | Lower Limb (Crural Fascia) | Lower Limb (Anterior Thigh Fascia): | Lower Limb (Iliotibial Tract) |
|---|---|---|---|---|---|---|
| 24 | 112.40 ± 6.1 | 652.73 ± 187.6 | 514.1 ± 140 | 486.3 ± 116.1 | 735.3 ± 44.26 | 269.4 ± 31.80 |
| 27 | 169.7 ± 10.30 | 328.38 ± 59.38 | 340 ± 40.41 | 365.5 ± 79.1 | 334.6 ± 44.74 | 230.6 ± 10.3 |
| 29 | - | 310.1 ± 41.22 | 162 ± 23 | 311 ± 18 | 285.4 ± 16.4 | 280.59 ± 5.84 |
| 36 | - | 292.54 ± 118.4 | 195 ± 21.04 | 305.1 ± 30 | 234.3 ± 12 | 368.2 ± 51.64 |
| 38 | 334.6 ± 44.74 | 364.83 ± 93.25 | 190 ± 23.02 | 225.7 ± 45 | 169.7 ± 10.30 | 147.8 ± 33.53 |
| 40 | 545.4 ± 22.33 | 147.11 ± 49.04 | 185.3 ± 94 | 273 ± 18.44 | 160.40 ± 6.1 | 140.3 ± 13.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pirri, C.; Petrelli, L.; Pérez-Bellmunt, A.; Ortiz-Miguel, S.; Fede, C.; De Caro, R.; Miguel-Pérez, M.; Stecco, C. Fetal Fascial Reinforcement Development: From “a White Tablet” to a Sculpted Precise Organization by Movement. Biology 2022, 11, 735. https://doi.org/10.3390/biology11050735
Pirri C, Petrelli L, Pérez-Bellmunt A, Ortiz-Miguel S, Fede C, De Caro R, Miguel-Pérez M, Stecco C. Fetal Fascial Reinforcement Development: From “a White Tablet” to a Sculpted Precise Organization by Movement. Biology. 2022; 11(5):735. https://doi.org/10.3390/biology11050735
Chicago/Turabian StylePirri, Carmelo, Lucia Petrelli, Albert Pérez-Bellmunt, Sara Ortiz-Miguel, Caterina Fede, Raffaele De Caro, Maribel Miguel-Pérez, and Carla Stecco. 2022. "Fetal Fascial Reinforcement Development: From “a White Tablet” to a Sculpted Precise Organization by Movement" Biology 11, no. 5: 735. https://doi.org/10.3390/biology11050735