
A fresh look at the Hyolithid Doliutheca from the Early Cambrian (Stage 4) Shipai Formation of the Three Gorges Area, Hubei, South China



Abstract
:Simple Summary
Abstract
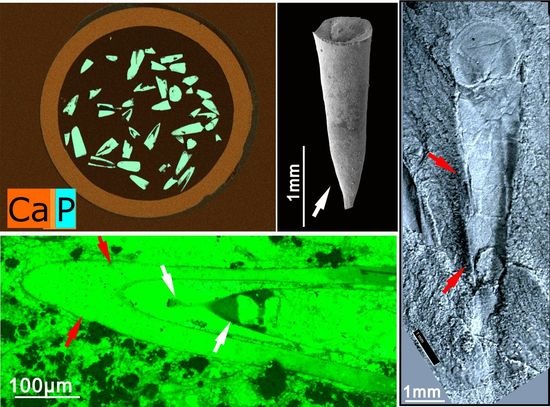
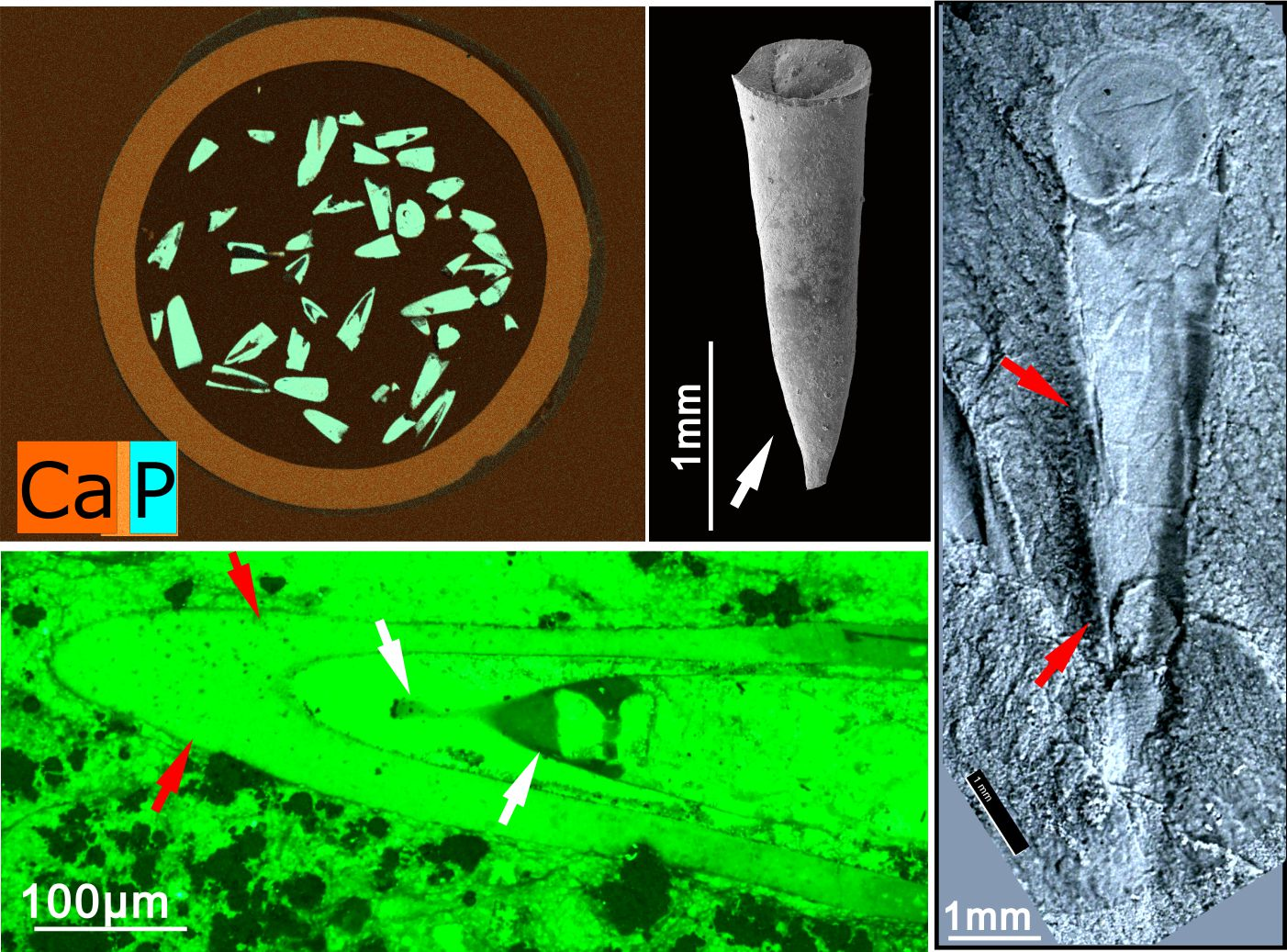{kind=link}
{kind=link}
{kind=link}
{kind=link}
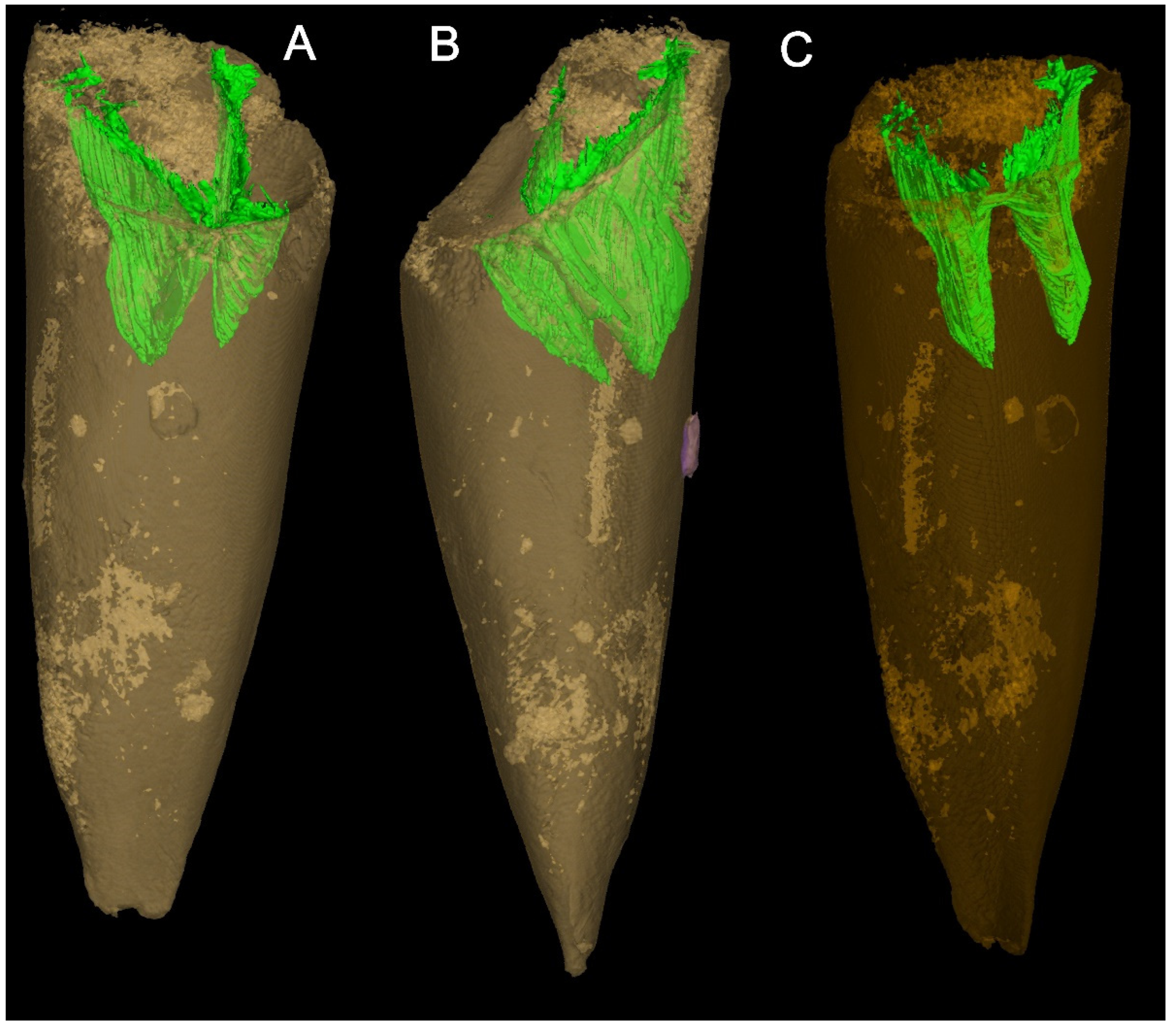{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
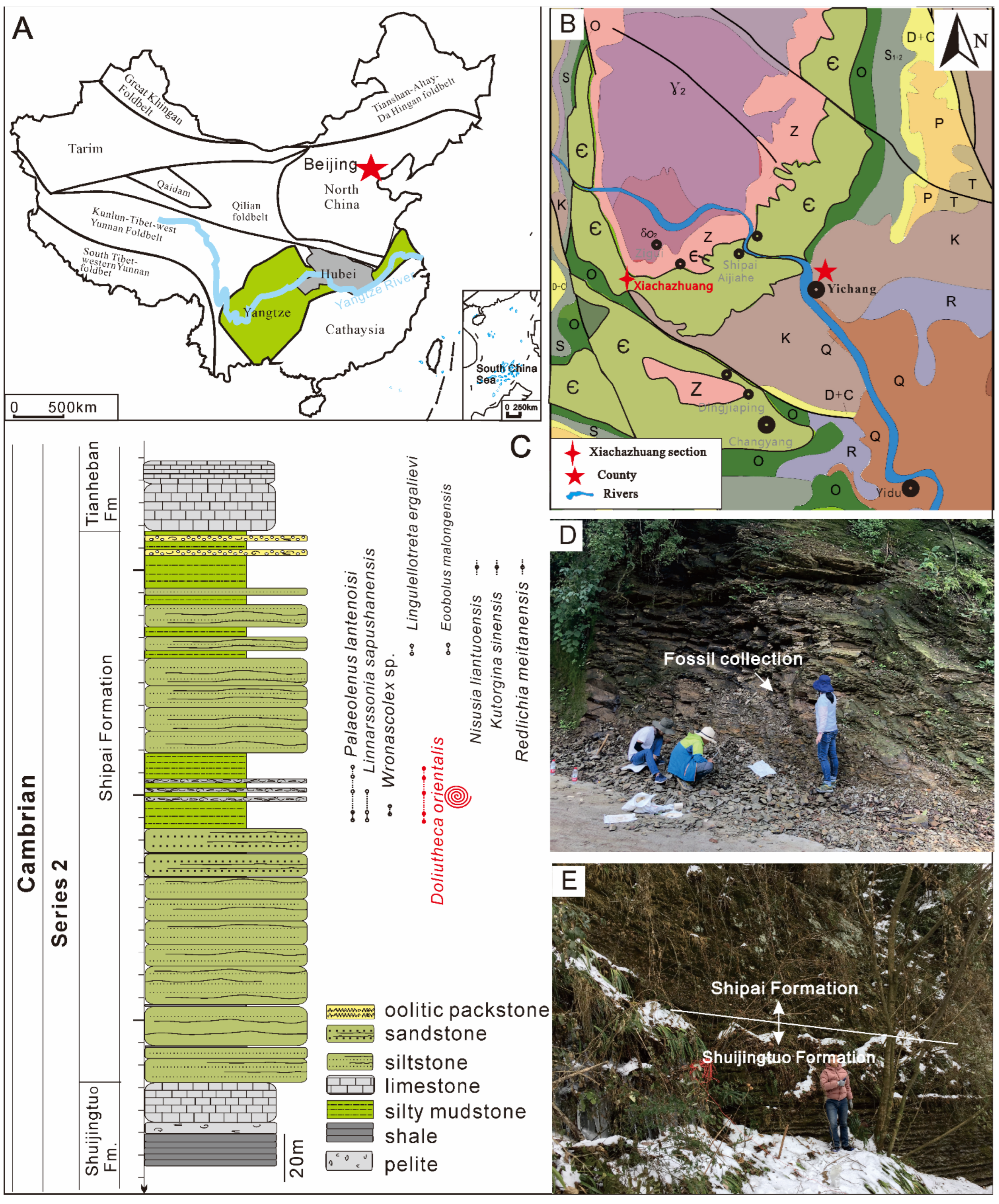
2. Geological Setting and Stratigraphy
3. Materials and Methods
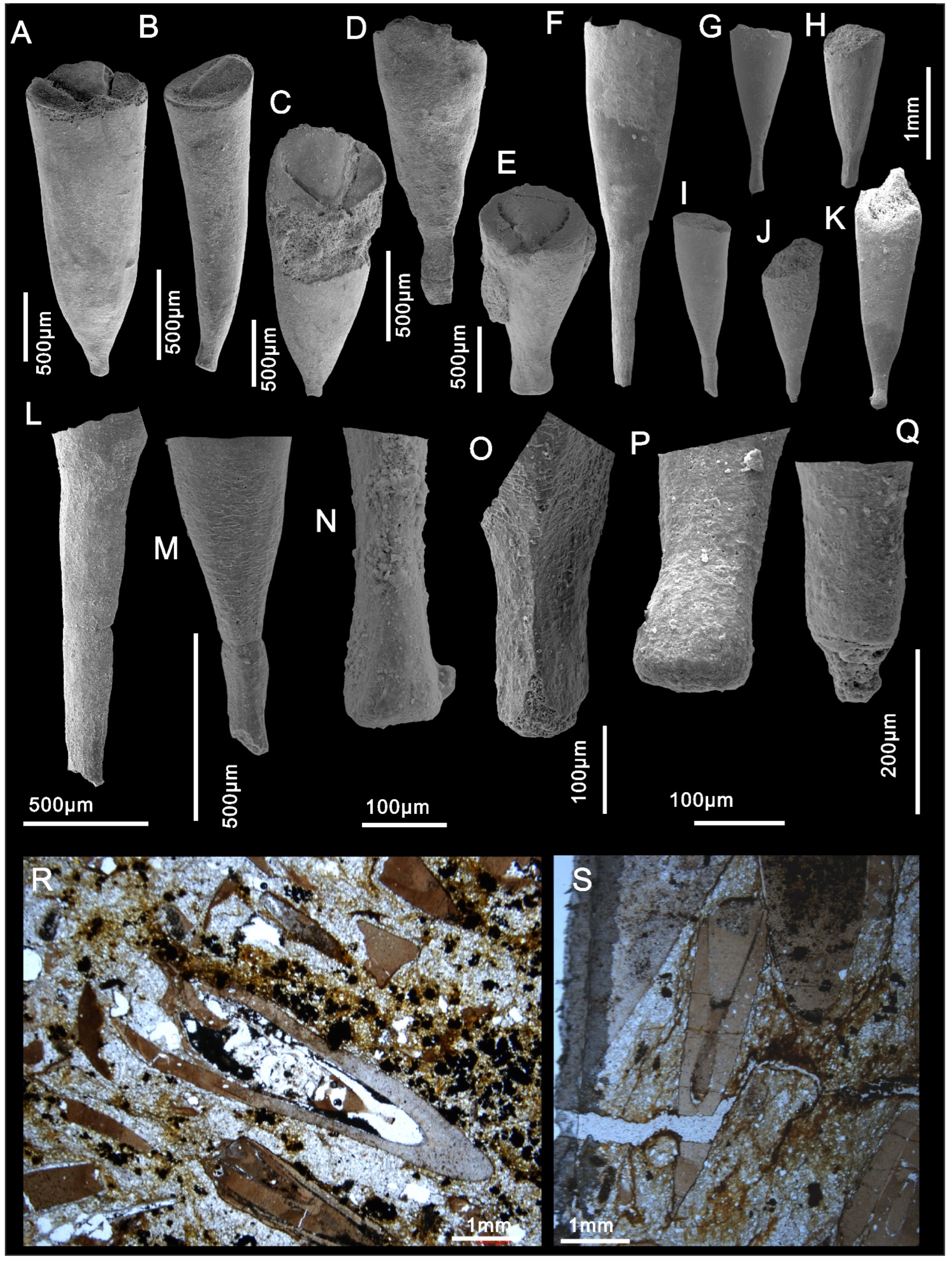
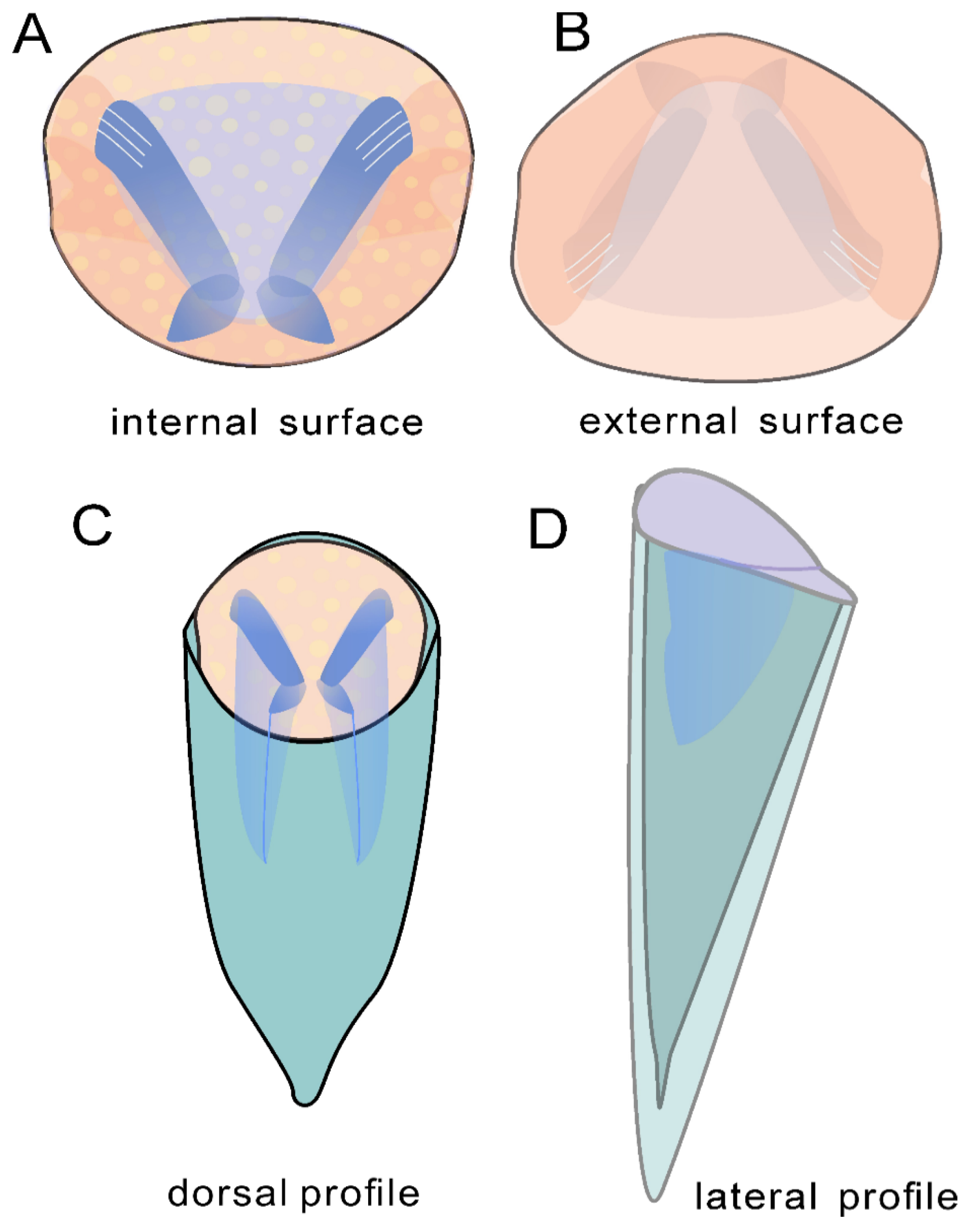
4. Systematic Paleontology
- Class Hyolitha Marek, 1963
- Family Paramicrocornidae Liu, Skovsted, Topper & Zhang, 2022
- Genus Doliutheca Qian, 1989
- Doliutheca orientalis (Qian, 1978)
5. Discussion
5.1. Hyolith Accumulations and Soft Part Preservation
5.2. Apical Variability
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Runnegar, B.; Pojeta, J.; Morris, N.J.; Taylor, J.D.; Taylor, M.E.; Mcclung, G. Biology of the Hyolitha. Lethaia 1975, 8, 181–191. [Google Scholar] [CrossRef]
- Qian, Y.; Xiao, L.G. Hyolitha; Scientific Publication: Beijing, China, 1995; p. 262. (In Chinese) [Google Scholar]
- Malinky, J.M.; Yochelson, E.L. On the systematic position of the Hyolitha (Kingdom Animalia). Mem. Assoc. Australas. Palaeontol. 2007, 34, 521–536. [Google Scholar]
- Marek, L. The class Hyolitha in the Caradoc of Bohemia. Sborník Geol. Věd Paleontol. 1967, 9, 51–112. [Google Scholar]
- Marek, L.; Yochelson, E.L. Paleozoic mollusk: Hyolithes. Science 1964, 146, 1674–1675. [Google Scholar] [CrossRef] [PubMed]
- Marek, L.; Yochelson, E.L. Aspects of the biology of Hyolitha (Mollusca). Lethaia 1976, 9, 65–82. [Google Scholar] [CrossRef]
- Dzik, J. Larval development of hyolithids. Lethaia 1978, 11, 293–299. [Google Scholar] [CrossRef]
- Mus, M.M.; Bergström, J. Skeletal microstructure of helens, lateral spines of hyolithids. Palaeontology 2007, 50, 1231–1243. [Google Scholar] [CrossRef]
- Mus, M.M.; Jeppsson, L.; Malinky, J.M. A complete reconstruction of the hyolithid skeleton. J. Paleontol. 2014, 88, 160–170. [Google Scholar]
- Malinky, J.M.; Skovsted, C.B. Hyoliths and small shelly fossils from the Lower Cambrian of North-East Greenland. Acta Palaeontol. Pol. 2004, 49, 551–578. [Google Scholar]
- Zhang, Z.L.; Skovsted, C.B.; Zhang, Z.F. A hyolithid without helens preserving the oldest hyolith muscle scars; palaeobiology of Paramicrocornus from the Shuijingtuo Formation (Cambrian Series 2) of South China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 489, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Skovsted, C.B.; Mus, M.M.; Zhang, Z.L.; Pan, B.; Li, L.Y.; Liu, F.; Li, G.X.; Zhang, Z.F. On the origin of hyolith helens. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 555, 109848. [Google Scholar] [CrossRef]
- Liu, F.; Skovsted, C.B.; Topper, T.P.; Zhang, Z. Hyolithid-like hyoliths without helens from the early Cambrian of South China, and their implications for the evolution of hyoliths. BMC Ecol. Evol. 2022, 22, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y. The Early Cambrian hyolithids in central and southwest China and their stratigraphical significance. Mem. Nanjing Inst. Geol. Palaeontol. 1978, 11, 1–43. [Google Scholar]
- Qian, Y. Early Cambrian Small Shelly Fossils of China with Special Reference to the Precambrian–Cambrian Boundary (2); Nanjing University Publishing House: Nanjing, China, 1989; p. 342. [Google Scholar]
- Qian, Y.; Xie, Y.S.; He, T.G. Hyoliths of the lower Cambrian Chiungchussuan Stage in South Shaanxi Province. Acta Palaeontol. Sin. 2001, 40, 37–43, (In Chinese with English abstract). [Google Scholar]
- Liu, F.; Chen, F.Y.; Chen, Y.L.; Zhang, Z.F. Note on the Shipai biota from the lower Cambrian (Series 2, Stage 4) of the Three Gorges Area, South China. Acta Palaeontol. Sinica. 2017, 56, 516–528, (In Chinese with English abstract). [Google Scholar]
- Ishikawa, T.; Ueno, Y.; Shu, D.; Li, Y.; Han, J.; Guo, J.; Yoshida, N.; Maruyama, S.; Komiya, T. The δ13C excursions spanning the Cambrian explosion to the Canglangpuian mass extinction in the Three Gorges area, South China. Gondwana Res. 2014, 25, 1045–1056. [Google Scholar] [CrossRef]
- Zhang, X.L.; Hua, H. Soft-bodied fossils from the Shipai Formation, lower Cambrian of the Three Gorge area, South China. Geol. Mag. 2005, 142, 699–709. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Zhang, Z.L.; Holmer, L.E.; Li, G.X. First report of linguloid brachiopods with soft parts from the lower Cambrian (Series 2, Stage 4) of the Three Gorges area, South China. Ann. De Paléontologie 2015, 101, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.L.; Betts, M.J.; Holmer, L.E.; Chen, Y.L.; Liu, F.; Liang, Y.; Zhang, Z.F. Early Cambrian (Stage 4) brachiopods from the Shipai Formation in the Three Gorges area of South China. J. Paleontol. 2021, 95, 497–526. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, W.; Zhao, Y. Cambrian Burgess Shale-type Lagerstätten in south China: Distribution and significance. Gondwana Res. 2008, 14, 255–262. [Google Scholar] [CrossRef]
- Wang, X.F.; Ni, S.Z.; Zeng, Q.L.; Xu, G.H.; Zhou, T.M.; Li, Z.H.; Lai, C. G Biostratigraphy of the Yangtze Gorge Area (2): Early Palaeozoic Era; Geological Publishing House: Beijing, China, 1987; pp. 4–38. [Google Scholar]
- Pan, B.; Skovsted, C.B.; Sun, H.; Li, G. Biostratigraphical and palaeogeographical implications of Early Cambrian hyoliths from the North China Platform. Alcheringa Australas. J. Palaeontol. 2019, 43, 351–380. [Google Scholar] [CrossRef]
- Luo, H.L.; Jiang, Z.W.; Wu, X.; Song, X.; Ouyang, L. The Sinian–Cambrian Boundary in Eastern Yunnan, China. Yunnan Inst. Geol. Sci. 1982, 265, 303, (In Chinese with English summary). [Google Scholar]
- Missarzhevsky, V.V. Descriptions of hyoliths, gastropods, hyolithelminths, camenids, and forms of an obscure systematic position [in Russian]. 105–175. In Tommotskij Ârus i Problema Nižnej Granicy Kembriâ; Rozanov, A.Û., Missarževskij, V.V., Volkova, N.A., Voronova, L.G., Krylov, I.N., Keller, B.M., Korolûk, I.K., Lendzion, K., Mihnâk, R., Pyhova, N.G., et al., Eds.; Trudy Ordena Trudovogo Krasnogo Znameni Geologičeskogo Instituta Akademii Nauk SSSR: Amerind, New Delhi, 1969; p. 359. [Google Scholar]
- Yang, B.; Steiner, M.; Keupp, H. Early Cambrian palaeobiogeography of the Zhenba–Fangxian Block (South China): Independent terrane or part of the Yangtze Platform? Gondwana Res. 2015, 28, 1543–1565. [Google Scholar] [CrossRef]
- Sun, H.J.; Zhao, F.C.; Steiner, M.; Li, G.X.; Na, L.; Pan, B.; Yin, Z.J.; Zeng, H.; Van Iten, H.; Zhu, M.Y. Skeletal faunas of the lower Cambrian Yu’anshan formation, eastern Yunnan, China: Metazoan diversity and community structure during the Cambrian age 3. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 542, 109580. [Google Scholar] [CrossRef]
- Morton, B.; Peharda, M.; Harper, E.M. Drilling and chipping patterns of bivalve prey predation by Hexaplex trunculus (Mollusca: Gastropoda: Muricidae). J. Mar. Biol. Assoc. 2007, 87, 933–940. [Google Scholar] [CrossRef]
- Klompmaker, A.A.; Kelley, P.H.; Chattopadhyay, D.; Clements, J.C.; Huntley, J.W.; Kowalewski, M. Predation in the marine fossil record: Studies, data, recognition, environmental factors, and behavior. Earth-Sci. Rev. 2019, 194, 472–520. [Google Scholar] [CrossRef]
- Robson, S.P.; Pratt, B.R. Predation of late Marjuman (Cambrian) linguliformean brachiopods from the Deadwood Formation of South Dakota, USA. Lethaia 2007, 40, 19–32. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Strotz, L.C.; Topper, T.P.; Chen, F.Y.; Chen, Y.; Liang, Y.; Zhang, Z.L.; Skovsted, C.B.; Brock, G.A. An encrusting kleptoparasite-host interaction from the early Cambrian. Nat. Commun. 2020, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Liu, F.; Liang, Y.; Hu, Y.Z.; Chen, F.Y.; Zhang, Z.L.; Chen, Y.L.; Ren, X.Y.; Yao, J.L.; Li, G.X.; et al. The Cambrian explosion of animals and the evolution of ecosystems on earth. J. Northwest. Univ. 2021, 51, 1065–1106. [Google Scholar]
- Chen, F.Y.; Topper, T.P.; Skovsted, C.B.; Strotz, L.C.; Shen, J.; Zhang, Z.F. Cambrian ecological complexities: Perspectives from the earliest brachiopod–supported benthic communities in the early Cambrian Guanshan Lagerstätte. Gondwana Res. 2022, 107, 30–41. [Google Scholar] [CrossRef]
- Vannier, J.; Chen, J. Early Cambrian food chain: New evidence from fossil aggregates in the Maotianshan Shale biota, SW China. Palaios 2005, 20, 3–26. [Google Scholar] [CrossRef]
- Sun, H.J.; Zhao, F.C.; Wen, R.Q.; Zeng, H.; Peng, J. Feeding strategy and locomotion of Cambrian hyolithides. Palaeoworld 2018, 27, 334–342. [Google Scholar] [CrossRef]
- Liu, F.; Skovsted, C.B.; Topper, T.P.; Zhang, Z.F. Soft part preservation in hyolithids from the lower Cambrian (Stage 4) Guanshan Biota of South China and its implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 562, 110079. [Google Scholar] [CrossRef]
- Liu, F.; Skovsted, C.B.; Topper, T.P.; Zhang, Z.F. Are hyoliths Palaeozoic lophophorates? Natl. Sci. Rev. 2020, 7, 453–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg-Madsen, V.; Valent, M.; Ebbestad, J.O.R. An orthothecid hyolith with a digestive tract from the early Cambrian of Bornholm, Denmark. GFF 2018, 140, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Li, L.Y.; Skovsted, C.B.; Yun, H.; Marissa, J.B.; Zhang, X.L. New insight into the soft anatomy and shell microstructures of early Cambrian orthothecids (Hyolitha). Proc. R. Soc. B 2020, 287, 20201467. [Google Scholar] [CrossRef] [PubMed]
- Dzik, J. Ontogeny of Bactrotheca and related hyoliths. Geol. Föreningen I Stockh. Förhandlingar 1980, 102, 223–233. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Skovsted, C.B.; Topper, T.P.; Zhang, Z. A fresh look at the Hyolithid Doliutheca from the Early Cambrian (Stage 4) Shipai Formation of the Three Gorges Area, Hubei, South China. Biology 2022, 11, 875. https://doi.org/10.3390/biology11060875
Liu F, Skovsted CB, Topper TP, Zhang Z. A fresh look at the Hyolithid Doliutheca from the Early Cambrian (Stage 4) Shipai Formation of the Three Gorges Area, Hubei, South China. Biology. 2022; 11(6):875. https://doi.org/10.3390/biology11060875
Chicago/Turabian StyleLiu, Fan, Christian B. Skovsted, Timothy P. Topper, and Zhifei Zhang. 2022. "A fresh look at the Hyolithid Doliutheca from the Early Cambrian (Stage 4) Shipai Formation of the Three Gorges Area, Hubei, South China" Biology 11, no. 6: 875. https://doi.org/10.3390/biology11060875
APA StyleLiu, F., Skovsted, C. B., Topper, T. P., & Zhang, Z. (2022). A fresh look at the Hyolithid Doliutheca from the Early Cambrian (Stage 4) Shipai Formation of the Three Gorges Area, Hubei, South China. Biology, 11(6), 875. https://doi.org/10.3390/biology11060875