
A Review of Histocytological Events and Molecular Mechanisms Involved in Intestine Regeneration in Holothurians

Abstract
:Simple Summary
Abstract
1. Introduction
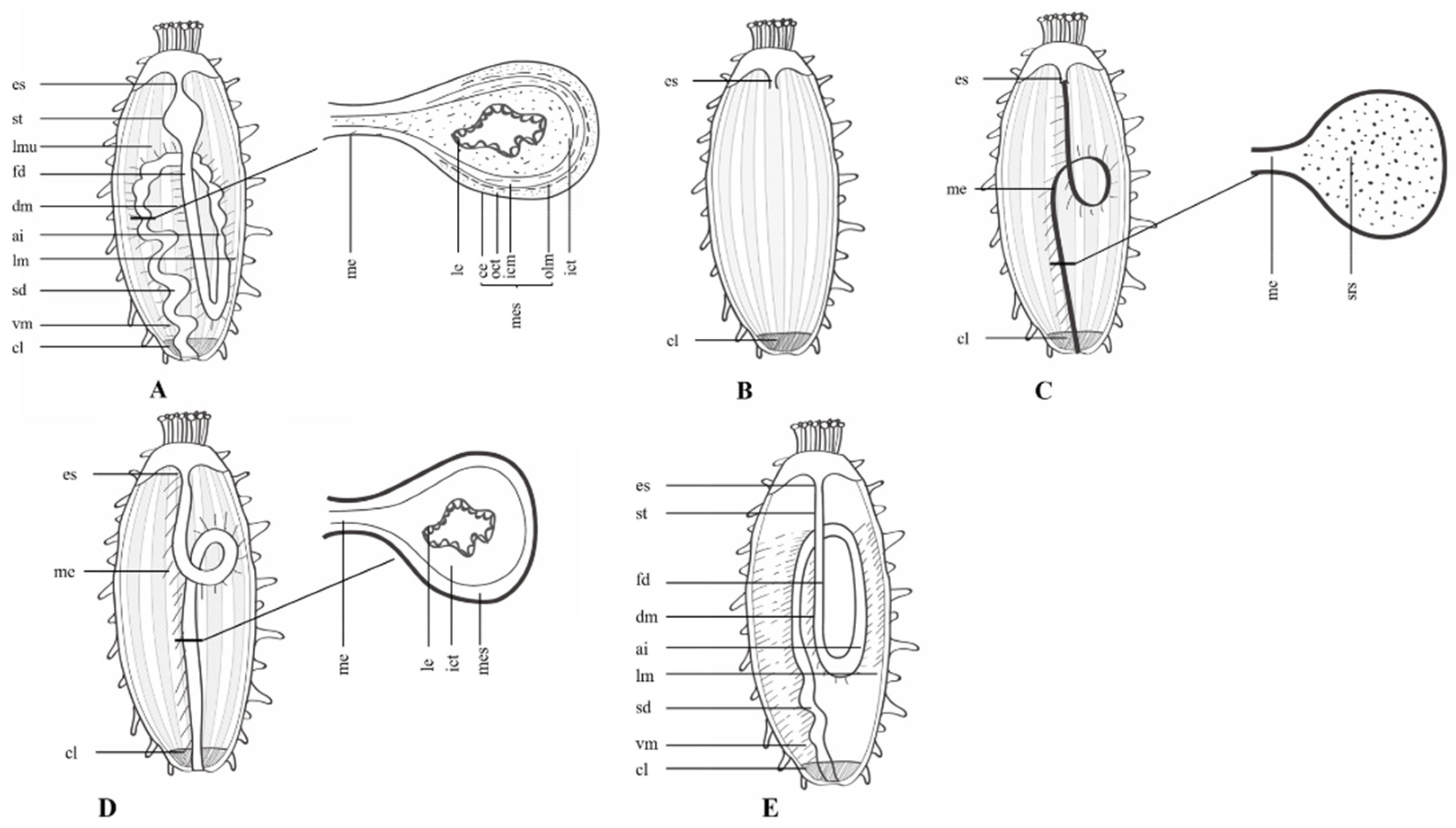
2. Autotomy and Evisceration
3. Histology and Cytology of Intestinal Regeneration
3.1. Wound Healing Stage
3.2. Anlage Formation Stage
3.3. Lumen Formation Stage
3.4. Intestinal Differentiation Stage
3.5. Intestinal Growth Stage
4. Molecular Mechanism of Intestinal Regeneration
4.1. Intestinal Regeneration
4.2. Neural Components Regeneration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alvarado, A.S. Regeneration in the metazoans: Why does it happen? BioEssays 2000, 22, 579. [Google Scholar] [CrossRef]
- Fumagalli, M.R.; Zapperi, S.; La Porta, C.A.M. Regeneration in distantly related species: Common strategies and pathways. NPJ Syst. Biol. Appl. 2018, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Zeng, A.; Li, H.; Guo, L.; Gao, X.; McKinney, S.; Wang, Y.; Yu, Z.; Park, J.; Semerad, C.; Ross, E.; et al. Prospectively Isolated Tetraspanin (+) Neoblasts Are Adult Pluripotent Stem Cells Underlying Planaria Regeneration. Cell 2018, 173, 1593–1608.e20. [Google Scholar] [CrossRef] [Green Version]
- Boehm, A.-M.; Khalturin, K.; Erxleben, F.A.; Hemmrich, G.; Klostermeier, U.C.; Lopez-Quintero, J.A.; Oberg, H.-H.; Puchert, M.; Rosenstiel, P.; Wittlieb, J.; et al. FoxO is a critical regulator of stem cell maintenance in immortal Hydra. Proc. Natl. Acad. Sci. USA 2012, 109, 19697–19702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galliot, B.; Ghila, L. Cell plasticity in homeostasis and regeneration. Mol. Reprod. Dev. 2010, 77, 837–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatani, Y.; Kawakami, A.; Kudo, A. Cellular and molecular processes of regeneration, with special emphasis on fish fins. Dev. Growth Differ. 2007, 49, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Arenas Gomez, C.M.; Gomez Molina, A.; Zapata, J.D.; Delgado, J.P. Limb regeneration in a direct-developing terrestrial salamander, Bolitoglossa ramosi (Caudata: Plethodontidae): Limb regeneration in plethodontid salamanders. Regeneration 2017, 4, 227–235. [Google Scholar] [CrossRef]
- Bhavsar, R.B.; Nakamura, K.; Tsonis, P.A. A System for Culturing Iris Pigment Epithelial Cells to Study Lens Regeneration in Newt. J. Vis. Exp. 2011, 52, e2713. [Google Scholar] [CrossRef]
- Brockes, J.P. Amphibian limb regeneration: Rebuilding a complex structure. Science 1997, 276, 81–87. [Google Scholar] [CrossRef]
- Dunoyer, L.A.; Seifert, A.W.; Van Cleve, J. Evolutionary bedfellows: Reconstructing the ancestral state of autotomy and regeneration. J. Exp. Zool. Part B Mol. Dev. Evol. 2020, 336, 94–115. [Google Scholar] [CrossRef]
- Stocum, D.L. Vertebrate regeneration. Semin. Cell Dev. Biol. 2002, 13, 325–326. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, M.D.C. Regeneration in Echinoderms: Repair, regrowth, cloning. Inf. Syst. J. 2006, 3, 64–76. [Google Scholar]
- Hernroth, B.; Farahani, F.; Brunborg, G.; Dupont, S.; Dejmek, A.; Skold, H.N. Possibility of mixed progenitor cells in sea star arm regeneration. J. Exp. Zool. B Mol. Dev. Evol. 2010, 314, 457–468. [Google Scholar] [CrossRef]
- Huet, M. Ultrastructural study and development of the neuroepithelial cells of regenerating epidermic arm cells in Asterina gibbosa Penn. Z. Zellforsch. Mikrosk. Anat. 1972, 126, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Huet, M. Role of the nervous system during the regeneration of an arm in a starfish: Asterina gibbosa Penn. (Echinodermata, Asteriidae). J. Embryol. Exp. Morphol. 1975, 33, 535–552. [Google Scholar] [PubMed]
- Vickery, M.S.; McClintock, J.B. Regeneration in metazoan larvae. Nature 1998, 394, 140. [Google Scholar] [CrossRef]
- Makra, A.; Keegan, B.F. Arm regeneration in Acrocnida brachiata (Ophiuroidea) at Little Killary, west coast of Ireland. Biol. Environ. Proc. R. Ir. Acad. 1999, 99B, 95–102. [Google Scholar]
- Reinardy, H.C.; Emerson, C.E.; Manley, J.M.; Bodnar, A.G. Tissue regeneration and biomineralization in sea urchins: Role of Notch signaling and presence of stem cell markers. PLoS ONE 2015, 10, e0133860. [Google Scholar] [CrossRef] [Green Version]
- García-Arrarás, J.E.; Greenberg, M.J. Visceral regeneration in holothurians. Microsc. Res. Tech. 2001, 55, 438–451. [Google Scholar] [CrossRef]
- Bertolini, F. Rigenerazione dell’Apparato Digerente nello Stichopus Regalis. Publ. Stn. Zool. Napoli. 1930, 10, 439–448. [Google Scholar]
- Zang, Y.Q.; Tian, X.L.; Dong, S.L.; Dong, Y.W. Growth, metabolism and immune responses to evisceration and the regeneration of viscera in sea cucumber, Apostichopus japonicus. Aquaculture 2012, 358-359, 50–60. [Google Scholar] [CrossRef]
- Bai, M.M. Regeneration in holothurian, holothuria-scabra jager. Indian J. Exp. Biol. 1971, 9, 467. [Google Scholar] [PubMed]
- García-Arrarás, J.E.; Lázaro-Peña, M.I.; Díaz-Balzac, C.A. Holothurians as a Model System to Study Regeneration. In Marine Organisms as Model Systems in Biology and Medicine; Kloc, M., Kubiak, J.Z., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 255–283. [Google Scholar]
- Byrne, M. The morphology of autotomy structures in the sea cucumber Eupentacta quinquesemita before and during evisceration. J. Exp. Biol. 2001, 204, 849–863. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, I.C. Variable tensility in echinoderm collagenous tissues—A review. Mar. Behav. Physiol. 1984, 11, 1–34. [Google Scholar] [CrossRef]
- Wilkie, I.C. Autotomy as a prelude to regeneration in echinoderms. Microsc. Res. Tech. 2001, 55, 369–396. [Google Scholar] [CrossRef]
- Mashanov, V.; García-Arrarás, J. Gut regeneration in holothurians: A snapshot of recent developments. Biol. Bull. 2011, 221, 93–109. [Google Scholar] [CrossRef]
- Miller, A.K.; Kerr, A.M.; Paulay, G.; Reich, M.; Wilson, N.G.; Carvajal, J.I.; Rouse, G.W. Molecular phylogeny of extant Holothuroidea (Echinodermata). Mol. Phylogenet. Evol. 2017, 111, 110–131. [Google Scholar] [CrossRef] [Green Version]
- Pearse, A.S. Autotomy in holothurians. Biol. Bull. 1909, 18, 42–49. [Google Scholar] [CrossRef]
- Byrne, M. The case for seasonal evisceration in the holothyroid Eupentacta quinquesemita (selenka), a reply. Can. J. Zool. 1986, 64, 2391–2392. [Google Scholar] [CrossRef]
- Swan, E.F. Seasonal evisceration in the sea cucumber, Parastichopus californicus (Stimpson). Science 1961, 133, 1078–1079. [Google Scholar] [CrossRef]
- Byrne, M. Induction of Evisceration in the Holothurian Eupentacta quinquesemita and Evidence for the Existence of an Endogenous Evisceration Factor. J. Exp. Biol. 1986, 120, 25–39. [Google Scholar] [CrossRef]
- Okada, A.; Kondo, M. Regeneration of the digestive tract of an anterior-eviscerating sea cucumber, Eupentacta quinquesemita, and the involvement of mesenchymal-epithelial transition in digestive tube formation. Zool. Lett. 2019, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashanov, V.S.; Dolmatov, I.Y.; Heinzeller, T. Transdifferentiation in holothurian gut regeneration. Biol. Bull. 2005, 209, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Kille, F.R. Regeneration in Thyone briareus Lesueur following induced autotomy. Biol. Bull. 1935, 69, 82–108. [Google Scholar] [CrossRef]
- Weigel, B.L. Sea Cucumber Intestinal Regeneration Reveals Deterministic Assembly of the Gut Microbiome. Appl. Environ. Microbiol. 2020, 86, e00489-20. [Google Scholar] [CrossRef] [PubMed]
- Shukalyuk, A.; Dolmatov, I.Y. Regeneration of the Digestive Tube in the Holothurian Apostichopus japonicusafter Evisceration. Russ. J. Mar. Biol. 2001, 27, 168–173. [Google Scholar] [CrossRef]
- Moussa Moussa, R.; El-Saied Nasef, L.; Abdel-Latif, H.A.A. Evisceration induction and gut regeneration of Holothuria polii (Delle Chiaje, 1824). Cah. Biol. Mar. 2018, 59, 309–315. [Google Scholar]
- Dolmatov, I.Y.; Khang, N.A.; Kamenev, Y.O. Asexual reproduction, evisceration, and regeneration in holothurians (Holothuroidea) from Nha Trang Bay of the South China Sea. Russ. J. Mar. Biol. 2012, 38, 243–252. [Google Scholar] [CrossRef]
- Ding, K.; Zhang, L.; Sun, L.; Lin, C.; Feng, Q.; Zhang, S.; Yang, H.; Brinkman, R.; Lin, G.; Huang, Z. Transcriptome analysis provides insights into the molecular mechanisms responsible for evisceration behavior in the sea cucumber Apostichopus japonicus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 30, 143–157. [Google Scholar] [CrossRef]
- Wilkie, I.C. Mutable collagenous tissue: Overview and biotechnological perspective. Prog. Mol. Subcell. Biol. 2005, 39, 221–250. [Google Scholar]
- Smith, G.N.; Greenberg, M.J. Chemical control of evisceration process in Thyone briareus. Biol. Bull. 1973, 144, 421–436. [Google Scholar] [CrossRef]
- Jenzri, M.; Gharred, C.; Bouraoui, Z.; Guerbej, H.; Jebali, J.; Gharred, T. Evisceration of Holothuria poli by mechanical, chemical and hypoxia stress methods and its bioremediation potentials for the pisciculture wastewater. Aquac. Res. 2022, 53, 3309–3317. [Google Scholar] [CrossRef]
- Ding, K.; Zhang, L.; Huo, D.; Guo, X.; Liu, X.; Zhang, S. Metabolomic analysis of coelomic fluids reveals the physiological mechanisms underlying evisceration behavior in the sea cucumber Apostichopus japonicus. Aquaculture 2021, 543, 736960. [Google Scholar] [CrossRef]
- Dobson, W.E.; Turner, R.L. Morphology and histology of the disk autotomy plane in Ophiophragmus filograneus (Echinodermata, Ophiurida). Zoomorphology 1989, 108, 323–332. [Google Scholar] [CrossRef]
- Dolmatov, I.Y. Asexual reproduction in holothurians. Sci. World J. 2014, 2014, 527234. [Google Scholar] [CrossRef] [Green Version]
- Dolmatov, I.Y. New data on asexual reproduction, autotomy, and regeneration in holothurians of the Order Dendrochirotida. Russ. J. Mar. Biol. 2014, 40, 228–232. [Google Scholar] [CrossRef]
- García-Arrarás, J.; Estrada-Rodgers, L.; Santiago, R.; Torres, I.I.; Díaz-Miranda, L.; Torres-Avillán, I. Cellular mechanisms of intestine regeneration in the sea cucumber, Holothuria glaberrima Selenka (Holothuroidea: Echinodermata). J. Exp. Zool. 1998, 281, 288–304. [Google Scholar] [CrossRef]
- Féral, J.-P.; Massin, C. Digestive systems: Holothurioidea. In Echinoderm Nutrition; Balkema: Rotterdam, The Netherlands, 1982; pp. 191–212. [Google Scholar]
- Foster, G.G.; Hodgson, A.N. Feeding, tentacle and gut morphology in five species of southern African intertidal holothuroids (Echinodermata). S. Afr. J. Zool. 1996, 31, 70–79. [Google Scholar] [CrossRef]
- Mashanov, V.; Frolova, L.; Dolmatov, I.Y. Structure of the digestive tube in the holothurian Eupentacta fraudatrix (Holothuroidea: Dendrochirota). Russ. J. Mar. Biol. 2004, 30, 314–322. [Google Scholar] [CrossRef]
- Ortiz-Pineda, P.A.; Ramírez-Gómez, F.; Pérez-Ortiz, J.; González-Díaz, S.; Santiago-De Jesús, F.; Hernández-Pasos, J.; Del Valle-Avila, C.; Rojas-Cartagena, C.; Suárez-Castillo, E.C.; Tossas, K.; et al. Gene expression profiling of intestinal regeneration in the sea cucumber. BMC Genom. 2009, 10, 262. [Google Scholar] [CrossRef] [Green Version]
- Filimonova, G.F.; Tokin, I.B. Structural and functional peculiarities of the digestive-system of Cucumaria frondosa (Echinodermata, Holothuroidea). Mar. Biol. 1980, 60, 9–16. [Google Scholar] [CrossRef]
- Boyko, A.V.; Girich, A.S.; Tkacheva, E.S.; Dolmatov, I.Y. The Eupentacta fraudatrix transcriptome provides insights into regulation of cell transdifferentiation. Sci. Rep. 2020, 10, 1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieves-Rios, C.; Alvarez-Falcon, S.; Malavez, S.; Rodriguez-Otero, J.; Garcia-Arraras, J.E. The nervous system component of the mesentery of the sea cucumber Holothuria glaberrima in normal and regenerating animals. Cell Tissue Res. 2019, 380, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Arraras, J.E.; Bello, S.A.; Malavez, S. The mesentery as the epicenter for intestinal regeneration. Semin. Cell Dev. Biol. 2019, 92, 45–54. [Google Scholar] [CrossRef]
- Candelaria, A.G.; Murray, G.; File, S.K.; García-Arrarás, J.E. Contribution of mesenterial muscle dedifferentiation to intestine regeneration in the sea cucumber Holothuria glaberrima. Cell Tissue Res. 2006, 325, 55–65. [Google Scholar] [CrossRef]
- Carnevali, M.D.C.; Bonasoro, F. Microscopic overview of crinoid regeneration. Microsc. Res. Tech. 2001, 55, 403–426. [Google Scholar] [CrossRef]
- Dolmatov, I.Y. Regeneration of the aquapharyngeal complex in the holothurian Eupentacta fraudatrix (Holothuroidea, Dendrochirota). Monogr. Dev. Biol. 1992, 23, 40. [Google Scholar]
- Dolmatov, I.Y.; Eliseikina, M.G.; Ginanova, T.; Lamash, N.; Korchagin, V.; Bulgakov, A. Muscle regeneration in the holothurian Stichopus japonicus. Dev. Genes Evol. 1996, 205, 486–493. [Google Scholar] [CrossRef]
- Dolmatov, I.Y.; Ginanova, T.T. Muscle regeneration in holothurians. Microsc. Res. Tech. 2001, 55, 452–463. [Google Scholar] [CrossRef]
- Dolmatov, I.Y. Molecular Aspects of Regeneration Mechanisms in Holothurians. Genes 2021, 12, 250. [Google Scholar] [CrossRef]
- Menton, D.N.; Eisen, A.Z. Cutaneous wound-healing in sea cucumber, Thyone-Briareus. J. Morphol. 1973, 141, 185–203. [Google Scholar] [CrossRef] [PubMed]
- San Miguel-Ruiz, J.E.; Garcia-Arraras, J.E. Common cellular events occur during wound healing and organ regeneration in the sea cucumber Holothuria glaberrima. BMC Dev. Biol. 2007, 7, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jose, E. Garcia-Arraras, I.Y.D. Echinoderms: Potential Model Systems for Studies on Muscle Regeneration. Curr. Pharm. Des. 2010, 16, 942–955. [Google Scholar]
- Vazzana, M.; Siragusa, T.; Arizza, V.; Buscaino, G.; Celi, M. Cellular responses and HSP70 expression during wound healing in Holothuria tubulosa (Gmelin, 1788). Fish Shellfish Immunol. 2015, 42, 306–315. [Google Scholar] [CrossRef]
- García-Arrarás, J.E.; Valentin-Tirado, G.; Flores, J.E.; Rosa, R.J.; Rivera-Cruz, A.; San Miguel-Ruiz, J.E.; Tossas, K. Cell dedifferentiation and epithelial to mesenchymal transitions during intestinal regeneration in H. glaberrima. BMC Dev. Biol. 2011, 11, 61–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girich, A.S.; Boyko, A.V. Wnt and Frizzled Genes in Echinoderms. Russ. J. Mar. Biol. 2019, 45, 302–312. [Google Scholar] [CrossRef]
- Leibson, N. Regeneration of digestive tube in holothurians Stichopus japonicus and Eupentacta fraudatrix. Monogr. Dev. Biol. 1992, 23, 51–61. [Google Scholar] [PubMed]
- Quinones, J.L.; Rosa, R.; Ruiz, D.L.; Garcia-Arraras, J.E. Extracellular matrix remodeling and metalloproteinase involvement during intestine regeneration in the sea cucumber Holothuria glaberrima. Dev. Biol. 2002, 250, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashanov, V.S.; Dolmatov, I.Y. Regeneration of digestive tract in the pentactulae of the far-eastern holothurian Eupentacta fraudatrix (Holothuroidea, Dendrochirota). Invertebr. Reprod. Dev. 2001, 39, 143–151. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Zueva, O.R.; Rojas-Catagena, C.; García-Arrarás, J.E. Visceral regeneration in a sea cucumber involves extensive expression of survivin and mortalin homologs in the mesothelium. BMC Dev. Biol. 2010, 10, 117–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawbin, W.H. Auto-evisceration and the regeneration of viscera in the holothurian Stichopus mollis (Hutton). Trans. Proc. R. Soc. N. Z. 1949, 77, 497–523. [Google Scholar]
- Chen, L.; Yao, F.; Qin, Y.; Shao, Y.; Fang, L.; Yu, X.; Wang, S.; Hou, L. The potential role of Kruppel-like factor 13 (Aj-klf13) in the intestine regeneration of sea cucumber Apostichopus japonicus. Gene 2020, 735, 144407. [Google Scholar] [CrossRef] [PubMed]
- Santiago, P.; Roig-López, J.L.; Santiago, C.; García-Arrarás, J.E. Serum amyloid A protein in an echinoderm: Its primary structure and expression during intestinal regeneration in the sea cucumber Holothuria glaberrima. J. Exp. Zool. 2000, 288, 335–344. [Google Scholar] [CrossRef]
- Mendez, A.T.; Roig-Lopez, J.L.; Santiago, P.; Santiago, C.; Garcia-Arraras, J.E. Identification of Hox gene sequences in the sea cucumber Holothuria glaberrima Selenka (Holothuroidea: Echinodermata). Mar. Biotechnol. 2000, 2, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Cardona, P.G.; Berrios, C.A.; Ramirez, F.; Garcia-Arraras, J.E. Lipopolysaccharides induce intestinal serum amyloid A expression in the sea cucumber Holothuria glaberrima. Dev. Comp. Immunol. 2003, 27, 105–110. [Google Scholar] [CrossRef]
- Suarez-Castillo, E.C.; Medina-Ortíz, W.E.; Roig-López, J.L.; E García-Arrarás, J. Ependymin, a gene involved in regeneration and neuroplasticity in vertebrates, is overexpressed during regeneration in the echinoderm Holothuria glaberrima. Gene 2004, 334, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.-X.; Sun, X.-Q.; Fang, B.-H.; Hong, X.-G.; Zhang, J.-X. Comparative analysis of genes expressed in regenerating intestine and non-eviscerated intestine of Apostichopus japonicus Selenka (Aspidochirotida: Stichopodidae) and cloning of ependymin gene. Hydrobiologia 2006, 571, 109–122. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Zueva, O.R.; Garcia-Arraras, J.E. Expression of Wnt9, TCTP, and Bmp1/Tll in sea cucumber visceral regeneration. Gene Expr. Patterns 2012, 12, 24–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.N.; Yang, H.S.; Chen, M.Y.; Xu, D.X. Cloning and expression analysis of Wnt6 and Hox6 during intestinal regeneration in the sea cucumber Apostichopus japonicus. Genet. Mol. Res. 2013, 12, 5321–5334. [Google Scholar] [CrossRef]
- Mei, Y.L.; Yao, F.; Wu, Y.; Chu, B.; Cheng, C.; Liu, Y.; Li, X.J.; Zou, X.Y.; Hou, L. Identification and expression of the elongator protein 2 (Ajelp2) gene, a novel regeneration-related gene from the sea cucumber Apostichopus japonicus. Mol. Biol. Rep. 2014, 41, 4985–4996. [Google Scholar] [CrossRef] [PubMed]
- Li, X.N.; Sun, L.N.; Yang, H.S.; Zhang, L.B.; Miao, T.; Xing, L.L.; Huo, D. Identification and expression characterization of WntA during intestinal regeneration in the sea cucumber Apostichopus japonicus. Comp. Biochem. Phys. B Biochem. Mil. Biol. 2017, 210, 55–63. [Google Scholar] [CrossRef]
- Adams, M.D.; Kelley, J.M.; Gocayne, J.D.; Dubnick, M.; Polymeropoulos, M.H.; Xiao, H.; Merril, C.R.; Wu, A.; Olde, B.; Moreno, R.F.; et al. Complementary DNA Sequencing—Expressed Sequence Tags and Human Genome Project. Science 1991, 252, 1651–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas-Cartagena, C.; Ortaz-Pineda, P.; Ramirez-Gomez, F.; Suarez-Castillo, E.C.; Matos-Cruz, V.; Rodriguez, C.; Ortiz-Zuazaga, H.; Garcia-Arraras, J.E. Distinct profiles of expressed sequence tags during intestinal regeneration in the sea cucumber Holothuria glaberrima. Physiol. Genom. 2007, 31, 203–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Chen, M.; Yang, H.; Wang, T.; Liu, B.; Shu, C.; Gardiner, D.M. Large Scale Gene Expression Profiling during Intestine and Body Wall Regeneration in the Sea Cucumber Apostichopus japonicus. Comp. Biochem. Physiol. Part. D Genom. Proteom. 2011, 6, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yang, H.; Chen, M.; Ma, D.; Lin, C. RNA-Seq Reveals Dynamic Changes of Gene Expression in Key Stages of Intestine Regeneration in the Sea Cucumber Apostichopus japonicus. PLoS ONE 2013, 8, e69441. [Google Scholar]
- Zhang, X.J.; Sun, L.N.; Yuan, J.B.; Sun, Y.M.; Gao, Y.; Zhang, L.B.; Li, S.H.; Dai, H.; Hamel, J.F.; Liu, C.Z.; et al. The sea cucumber genome provides insights into morphological evolution and visceral regeneration. PLoS Biol. 2017, 15, e2003790. [Google Scholar] [CrossRef] [Green Version]
- Medina-Feliciano, J.G.; Pirro, S.; Garcia-Arraras, J.E.; Mashanov, V.; Ryan, J.F. Draft Genome of the Sea Cucumber Holothuria glaberrima, a Model for the Study of Regeneration. Front. Mar. Sci. 2021, 8, 603410. [Google Scholar] [CrossRef]
- Yuan, J.; Gao, Y.; Sun, L.; Jin, S.; Zhang, X.; Liu, C.; Li, F.; Xiang, J. Wnt Signaling Pathway Linked to Intestinal Regeneration via Evolutionary Patterns and Gene Expression in the Sea Cucumber Apostichopus japonicus. Front. Genet. 2019, 10, 112. [Google Scholar] [CrossRef]
- Viera-Vera, J.; García-Arrarás, J.E. Retinoic Acid Signaling Is Associated with Cell Proliferation, Muscle Cell Dedifferentiation, and Overall Rudiment Size during Intestinal Regeneration in the Sea Cucumber, Holothuria glaberrima. Biomolecules 2019, 9, 873. [Google Scholar] [CrossRef] [Green Version]
- Girich, A.S.; Isaeva, M.P.; Dolmatov, I.Y. Wnt and frizzled expression during regeneration of internal organs in the holothurian Eupentacta fraudatrix. Wound Repair Regen. 2017, 25, 828–835. [Google Scholar] [CrossRef]
- Kawakami, Y.; Esteban, C.R.; Raya, M.; Kawakami, H.; Martí, M.; Dubova, I.; Belmonte, J.C.I. Wnt/β-catenin signaling regulates vertebrate limb regeneration. Genes Dev. 2006, 20, 3232–3237. [Google Scholar] [CrossRef] [Green Version]
- Novikova, E.L.; Bakalenko, N.I.; Nesterenko, A.Y.; Kulakova, M.A. Hox genes and animal regeneration. Russ. J. Dev. Biol. 2016, 47, 173–180. [Google Scholar] [CrossRef]
- Dolmatov, I.Y.; Kalacheva, N.V.; Tkacheva, E.S.; Shulga, A.P.; Zavalnaya, E.G.; Shamshurina, E.V.; Girich, A.S.; Boyko, A.V.; Eliseikina, M.G. Expression of Piwi, MMP, TIMP, and Sox during Gut Regeneration in Holothurian Eupentacta fraudatrix (Holothuroidea, Dendrochirotida). Genes 2021, 12, 1292. [Google Scholar] [CrossRef] [PubMed]
- Miao, T.; Wan, Z.; Sun, L.; Li, X.; Xing, L.; Bai, Y.; Wang, F.; Yang, H. Extracellular matrix remodeling and matrix metalloproteinases (ajMMP-2 like and ajMMP-16 like) characterization during intestine regeneration of sea cucumber Apostichopus japonicus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2017, 212, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Pasos, J.; Valentin-Tirado, G.; Garcia-Arraras, J.E. Melanotransferrin: New Homolog Genes and Their Differential Expression during Intestinal Regeneration in the Sea Cucumber Holothuria glaberrima. J. Exp. Zool. Part B Mol. Dev. Evol. 2017, 328, 259–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashanov, V.S.; Zueva, O.R.; Garcia-Arraras, J.E. Expression of pluripotency factors in echinoderm regeneration. Cell Tissue Res. 2015, 359, 521–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolmatov, I.Y.; Shulga, A.P.; Ginanova, T.T.; Eliseikina, M.G.; Lamash, N.E. Metalloproteinase inhibitor GM6001 delays regeneration in holothurians. Tissue Cell 2019, 59, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bello, S.A.; Torres-Gutierrez, V.; Rodriguez-Flores, E.J.; Toledo-Roman, E.J.; Rodriguez, N.; Diaz-Diaz, L.M.; Vazquez-Figueroa, L.D.; Cuesta, J.M.; Grillo-Alvarado, V.; Amador, A.; et al. Insights into intestinal regeneration signaling mechanisms. Dev. Biol 2019, 458, 12–31. [Google Scholar] [CrossRef]
- Bello, S.A.; Abreu-Irizarry, R.J.; Garcia-Arraras, J.E. Primary cell cultures of regenerating holothurian tissues. Methods Mol. Biol. 2015, 1189, 283–297. [Google Scholar]
- García-Arrarás, J.; Díaz-Miranda, L.; Torres, I.I.; File, S.; Jimenez, L.B.; Rivera-Bermudez, K.; Arroyo, E.J.; Cruz, W. Regeneration of the enteric nervous system in the sea cucumber Holothuria glaberrima. J. Comp. Neurol. 1999, 406, 461–475. [Google Scholar] [CrossRef]
- Tossas, K.; Qi-Huang, S.; Cuyar, E.; Garcia-Arraras, J.E. Temporal and spatial analysis of enteric nervous system regeneration in the sea cucumber Holothuria glaberrima. Regeneration 2014, 1, 10–26. [Google Scholar] [CrossRef] [PubMed]
- Siebert, S.; Farrell, J.A.; Cazet, J.F.; Abeykoon, Y.; Primack, A.S.; Schnitzler, C.E.; Juliano, C.E. Stem cell differentiation trajectories in Hydra resolved at single-cell resolution. Science 2019, 365, eaav9314. [Google Scholar] [CrossRef] [PubMed]
- Plass, M.; Solana, J.; Wolf, F.A.; Ayoub, S.; Misios, A.; Glazar, P.; Obermayer, B.; Theis, F.J.; Kocks, C.; Rajewsky, N. Cell type atlas and lineage tree of a whole complex animal by single-cell transcriptomics. Science 2018, 360, eaaq1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, T.; Murawala, P.; Knapp, D.; Masselink, W.; Schuez, M.; Hermann, S.; Gac-Santel, M.; Nowoshilow, S.; Kageyama, J.; Khattak, S.; et al. Single-cell analysis uncovers convergence of cell identities during axolotl limb regeneration. Science 2018, 362, eaaq0681. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.; Tran, N.T.T.; Diaz, D.C.; Tsai, Y.Y.; Acedo, J.N.; Lush, M.E.; Piotrowski, T. Single-cell transcriptome analysis reveals three sequential phases of gene expression during zebrafish sensory hair cell regeneration. Dev. Cell 2022, 57, 799–819.e6. [Google Scholar] [CrossRef]


{kind=link}
| Species | Order | Eviscerating | Regenerating | Reference | |||
|---|---|---|---|---|---|---|---|
| Autotomy Site | Evisceration Site | Excreted Viscera | Initial Regeneration Site | Regeneration Cycle | |||
| Eupentacta quinquesemita | Dendrochirotida | (1) the oral body wall/the oral complex (2) the longitudinal muscle/the retractor muscle (3) intestine/mesentery (4) intestine/cloaca | rupture of anterior end of the body (anterior) | the oral complex; the digestive tract; part of the gonads | the free end of the anterior mesentery | 2–3 weeks | [33] |
| Eupentacta fraudatrix | (1) rupture of the introvert (2) the pharyngeal retractor muscles/the pharyngeal retractor muscles (3) intestine/cloaca (4) intestine/mesentery | the entire digestive tube; the oral complex | free end of the dorsal mesentery at the healed oral end of the body; the free end of the ventral mesentery at the cloaca | 27 days | [34] | ||
| Sclerodactyla briareus | (1) the introvert and the muscular body wall (2) the intestine/the cloaca (3) intestine/mesentery | stomach; intestine; oral complex | the free end of the remaining mesentery | 20–37 days | [35,36] | ||
| Holothuria glaberrima | Holothuriida Synallactida | (1) the esophagus/the descending small intestine (2) the large intestine/the cloaca (3) intestine/mesentery | cloaca (posterior) | the intestinal system; the hemal system; the right respiratory tree; most of the gonads | 28 days | [23] | |
| Apostichopus japonicus | (1) the esophagus/the stomach (2) the intestine/the cloaca (3) intestine/mesentery | the intestinal system (except esophagus) | the free end of the remaining mesentery; anlage arise from the esophagus; anlage arise from the cloaca | 21 days | [37] | ||
| Holothuria polii | (1) the esophagus/the stomach (2) the intestine/the cloaca (3) intestine/mesentery | the intestinal system (except pharynx and esophagus); the left respiratory tree; gonads | 6 weeks | [38] | |||
| Genes Type | Gene Name | Gene Expression During Regeneration | Species | Expression Site | Putative Function | Process Involved in | Reference |
|---|---|---|---|---|---|---|---|
| MMPs | Ef-72 kDa type IV collagenase | - | Eupentacta fraudatrix | coelomic and luminal epithelia | degradation of ECM proteins and facilitating cell movement | ECM remodeling and cell migration | [95] |
| Ef-MMP16 | - | Eupentacta fraudatrix | coelomic epithelium | migration and/or proliferation of coelomic epithelial cells | migration of coelomic epithelial cells | [95] | |
| ajMMP-16 like | high expression during the whole process, with the highest expression at 1 dpe followed by a constant drop to normal level from 7 dpe to 21 dpe | Apostichopus japonicus | no expression in normal intestine; in the regenerative intestine, expressed at coelomic and luminal epithelia | degrading ECM and growth factors; targeting ECM components and biological molecules | ECM remodeling | [96] | |
| ajMMP-2 like | highest expression at 6 hpe, slightly decreasing to approximately 2 at 1 dpe and 3 dpe, gradually declining to normal level | Apostichopus japonicus | no expression in normal intestine; in the regenerative intestine, expressed at luminal epithelia | [96] | |||
| TIMPs | Ef-tensilin3 | - | Eupentacta fraudatrix | coelomic epithelium and the ventral part of the luminal epithelium; opposite to EF-MMP16 | inhibiting the activity of MMPs | ECM remodeling | [95] |
| immune-related gene | Serum amyloid A | high expression during the whole process, with the highest expression at 15 dpe | Holothuria glaberrima | coelomic epithelium | stimulate cell migration and adhesion to an ECM substrate; involved in the formation of luminal epithelium and muscular layer | ECM remodeling; formation of luminal epithelium and the muscular layers | [75] |
| Melanotransferrin (MTf) | Hg MTf1/Aj MTf | increased expression from day 3, reaching the peak at 7 dpe | Holothuria glaberrima | mesothelium | immune activation | dedifferentiation of mesothelial cells and the following proliferation and migration | [97] |
| Hg MTf2 | |||||||
| Hg MTf3 | high expression at 3 dpe and 5 dpe, followed by decreasing gradually to normal level | ||||||
| Hg MTf4 | |||||||
| Wnt signaling pathway | WntA | up-regulated significantly from 6 h to day 14 with the maximum expression at 14 dpe | Apostichopus japonicus | in the luminal epidermal, muscle layer, and submucosa | be connected with cell proliferation | the proliferation, dedifferentiation and migration of luminal epithelium, muscle layer, and submucosa cells and apoptosis in the basal lamina of the mucosal epithelium at each stage of wound healing | [83] |
| Wnt9 | strongest expression on days 7–14 | Holothuria glaberrima | in normal intestine, no expression; in the regenerative intestine, expressed at mesothelium | control the transitions between the dedifferentiated mesothelial cells and the mesenchyma | a series of activities of the mesothelium | [80] | |
| gene involved in embryogenesis | Bmp1/Tll | up-regulated on days 3 through 12 | Holothuria glaberrima | in the apices of the developing folds of the luminal epithelial | remodeling of extracellular matrix | morphogenetic movements leading to folding of the luminal epithelium and gut looping | [80] |
| TCTP | highest expression at 3 dpe and 21 dpe | Holothuria glaberrima | in normal intestine, expressed at the apices of the luminal epithelial; in the regenerative intestine, expressed at mesothelium similar to survivin and mortalin | apoptosis suppression and regulation cell proliferation | deep transient dedifferentiation of mesothelial cells | [80] | |
| survivin | insignificant increase at 7 dpe and 14 dpe, significant increase at 21 dpe | Holothuria glaberrima | in normal intestine, expressed at the base of the luminal epithelium; in the regenerative intestine, mostly expressed at mesothelium | cell proliferation; suppression of the programmed cell death | dedifferentiation of mesothelial cells and extensive proliferation | [72] | |
| mortalin | two peaks of roughly 3-fold up-regulation at 7 dpe and 21 dpe | Holothuria glaberrima | in normal and regenerative intestine, expressed at mesothelium | cell proliferation | [72] | ||
| piwi | high expression during the whole process, with the highest expression at 3 dpe | Eupentacta fraudatrix | in normal intestine, expressed at mesothelium; in the regenerative intestine, expressed at ECM | - | do not participate in the formation of the luminal epithelium | [95] | |
| Sox gene family | Ef-Sox9/10 | - | Eupentacta fraudatrix | coelomic epithelium, mesenchymal cells, and the developing luminal epithelium | regulating the differentiation of mesenchymal cells into epithelial cells | coelomic epithelium transdifferentiation; the redifferentiation of myoepithelial cells and the formation of muscle layer | [95] |
| Ef-Sox17 | - | Eupentacta fraudatrix | at the site of immersion only in surface cells | regulation of the initial stages of transdifferentiation | [95] | ||
| pluripotency factors (Yamanaka factors) | SoxB1 | down-regulation at the early post-evisceration stages (3 dpe and 7 dpe) | Holothuria glaberrima | in normal intestine, in the luminal epithelium; no signal at 3 dpe; expressed in the mesothelium at 7 dpe; finally expressed in the luminal epithelium | not essential for cell dedifferentiation | - | [98] |
| Myc | Up-regulated at 3 dpe, but then returned to normal level, before being slightly (∼1.5 fold) down-regulated at 21 dpe | in normal intestine, in the luminal epithelium and mesothelium; expressed in the mesothelium at 3 dpe; expressed in the mesothelial epithelial cells at 7 dpe; finally expressed in the luminal epithelium | play an important role in dedifferentiation/regeneration | dedifferentiation of mesothelium | [98] | ||
| Klf1/2/4 | no significant differences | in normal intestine, in the luminal epithelium; expressed in the mesothelium at 3 dpe; expressed in the mesothelial epithelial cells at 7 dpe; finally expressed in the luminal epithelium | - | [98] | |||
| Oct 1/2/11 | - | [98] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, F.; Yang, H.; Sun, L. A Review of Histocytological Events and Molecular Mechanisms Involved in Intestine Regeneration in Holothurians. Biology 2022, 11, 1095. https://doi.org/10.3390/biology11081095
Su F, Yang H, Sun L. A Review of Histocytological Events and Molecular Mechanisms Involved in Intestine Regeneration in Holothurians. Biology. 2022; 11(8):1095. https://doi.org/10.3390/biology11081095
Chicago/Turabian StyleSu, Fang, Hongsheng Yang, and Lina Sun. 2022. "A Review of Histocytological Events and Molecular Mechanisms Involved in Intestine Regeneration in Holothurians" Biology 11, no. 8: 1095. https://doi.org/10.3390/biology11081095
APA StyleSu, F., Yang, H., & Sun, L. (2022). A Review of Histocytological Events and Molecular Mechanisms Involved in Intestine Regeneration in Holothurians. Biology, 11(8), 1095. https://doi.org/10.3390/biology11081095