

Integrated mRNA and miRNA Expression Profile Analysis of Female and Male Gonads in Acrossocheilus fasciatus
 , ,
, , 
Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Fish, Samples Collection, and RNA Extraction
2.2. cDNA Library, Small RNA Preparation, and Sequencing
2.3. mRNA sequencing and miRNA Sequencing Analysis, and Assembly
2.4. Differential Expression Analysis of mRNAs and miRNAs, Prediction and Annotation of DEGs and DEMs
2.5. Validation of Sex-Biased mRNAs and miRNAs
2.6. Cell Transfection and Dual-Luciferase Reporter Assay
3. Results
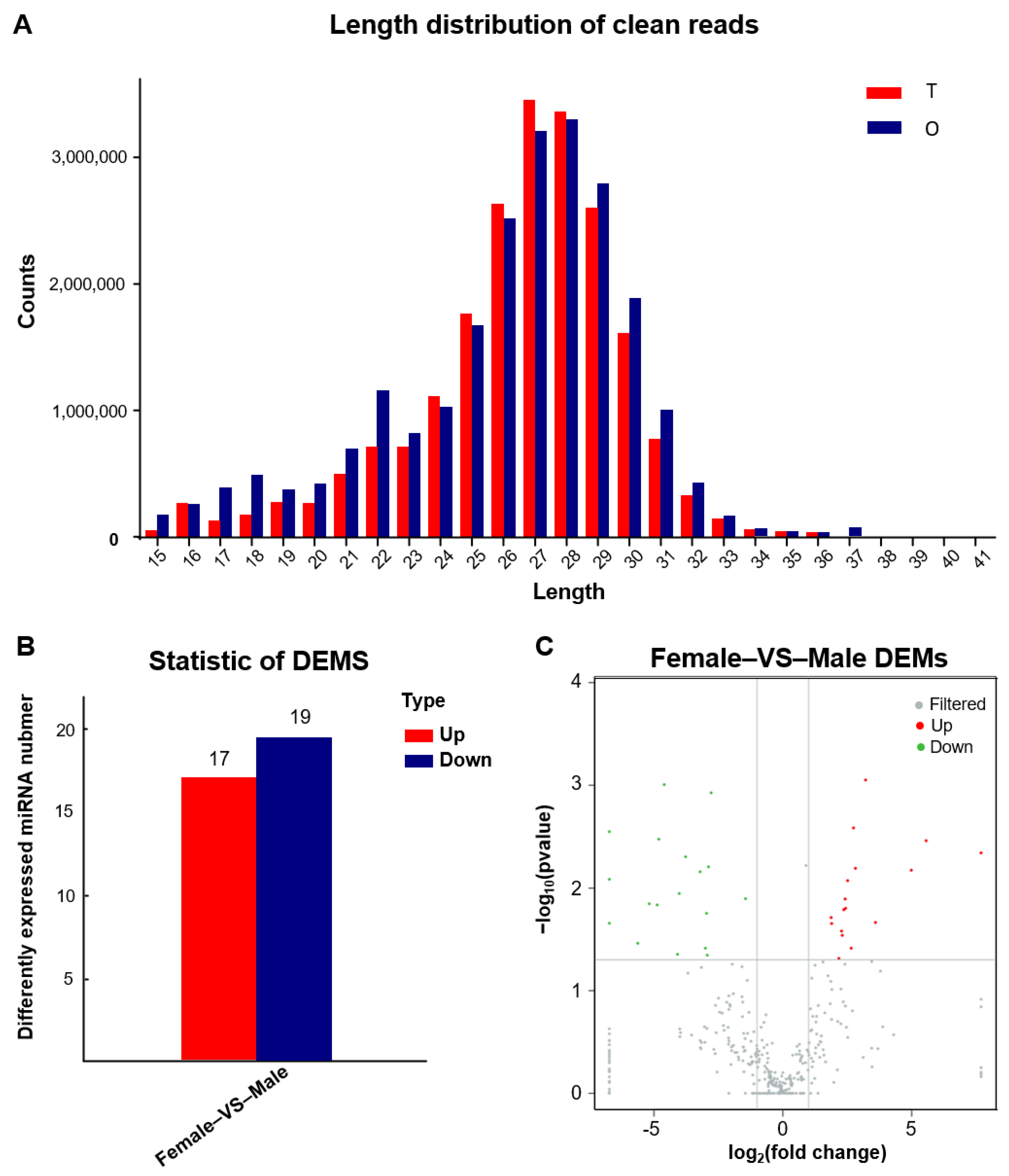
3.1. Characterization of Gonadal mRNAs and miRNAs
3.2. Screening for Differentially Expressed Genes (DEGs) and Functional Annotation
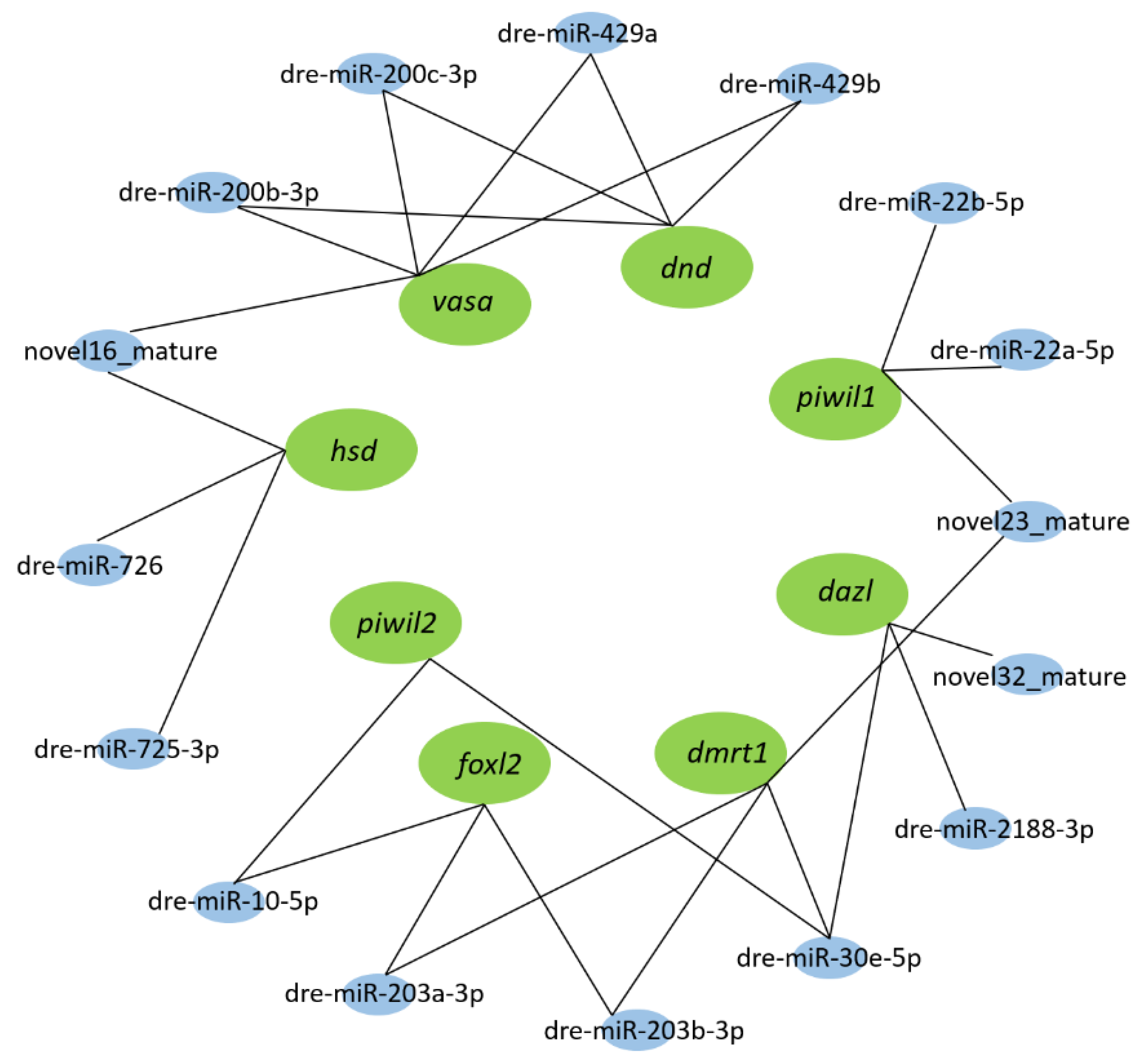
3.3. Screening for Differentially Expressed miRNAs (DEMs) and Functional Annotation of Their Target Genes
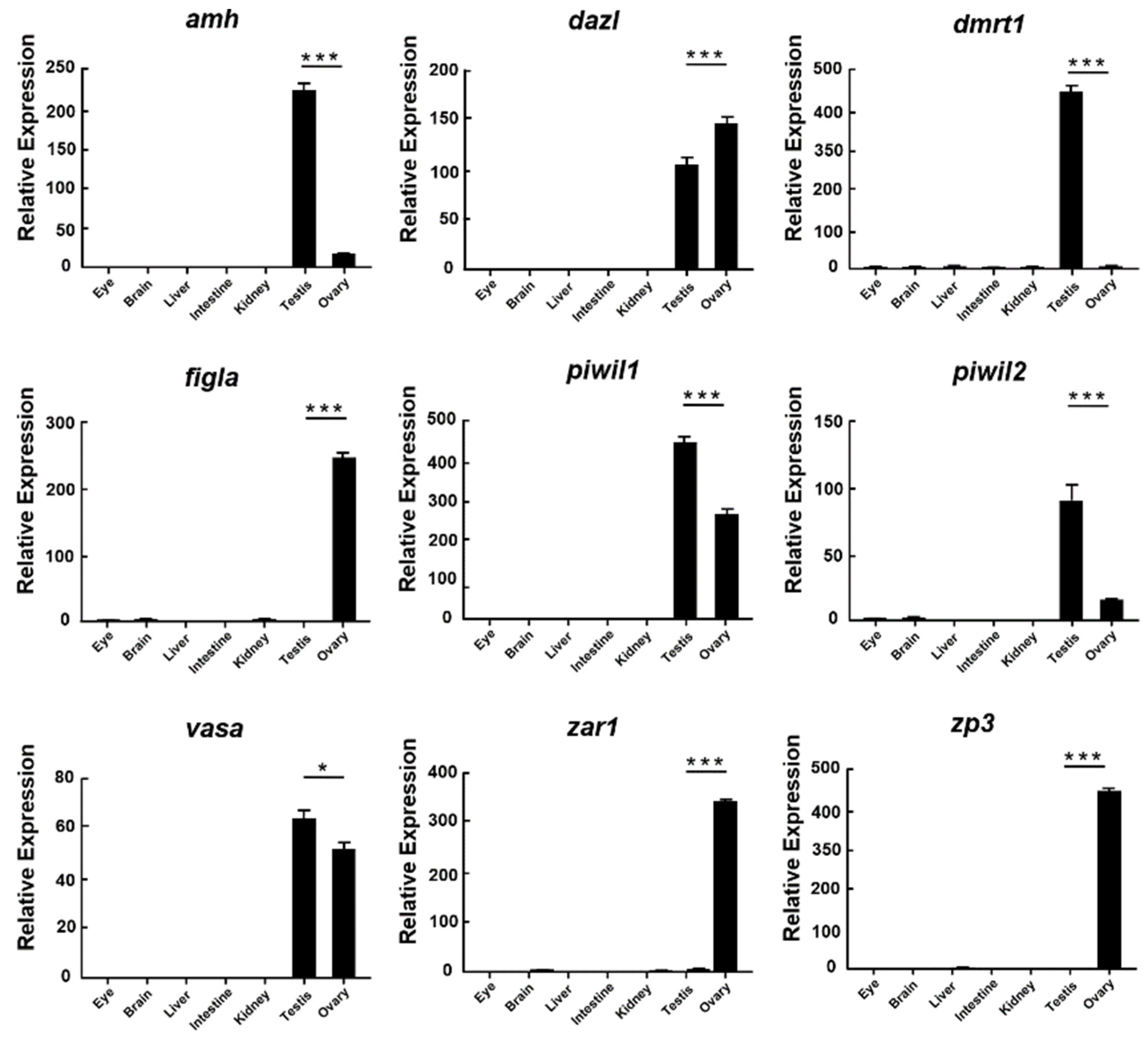
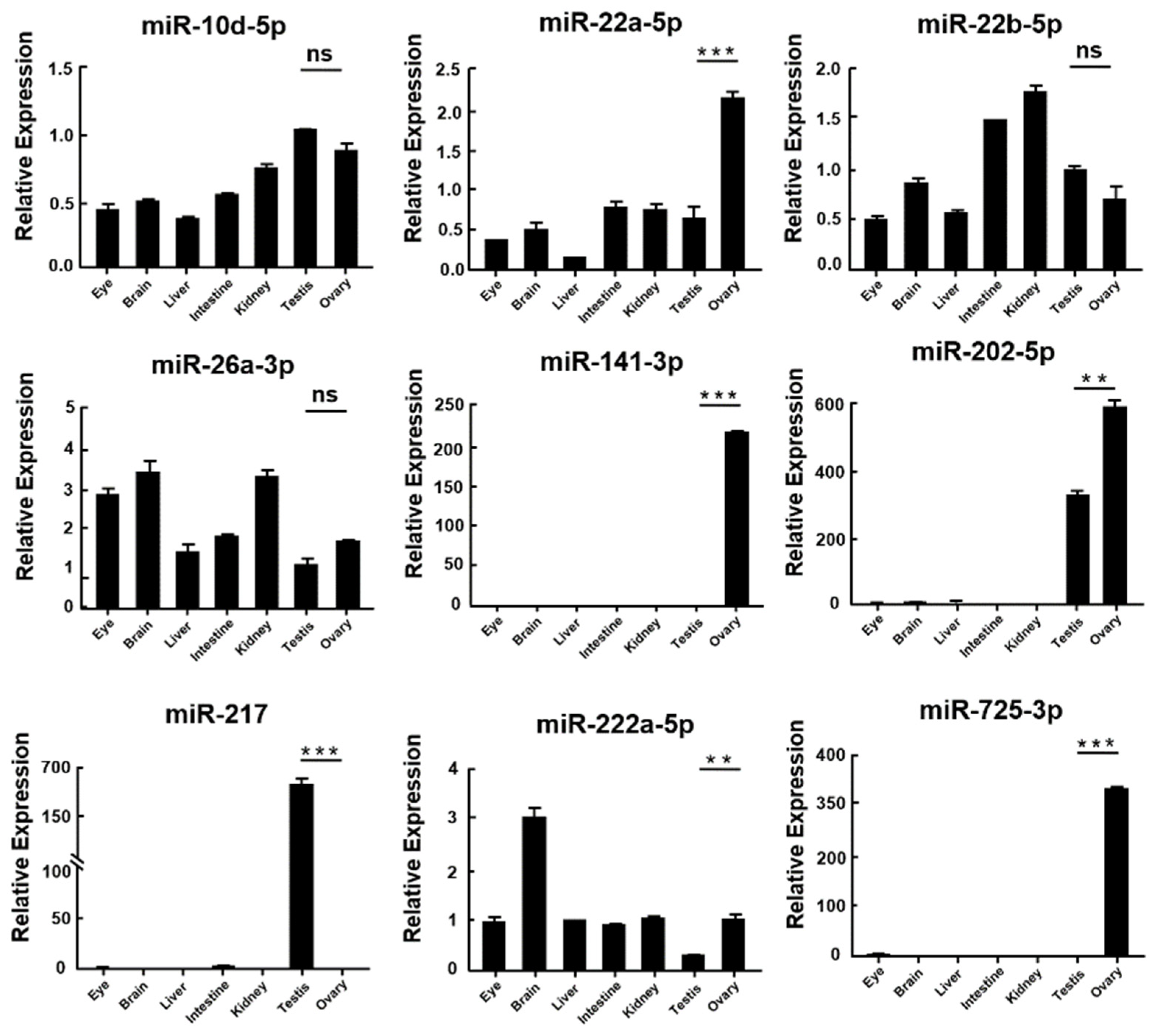
3.4. Validation of miRNAs and mRNA by qRT-PCR
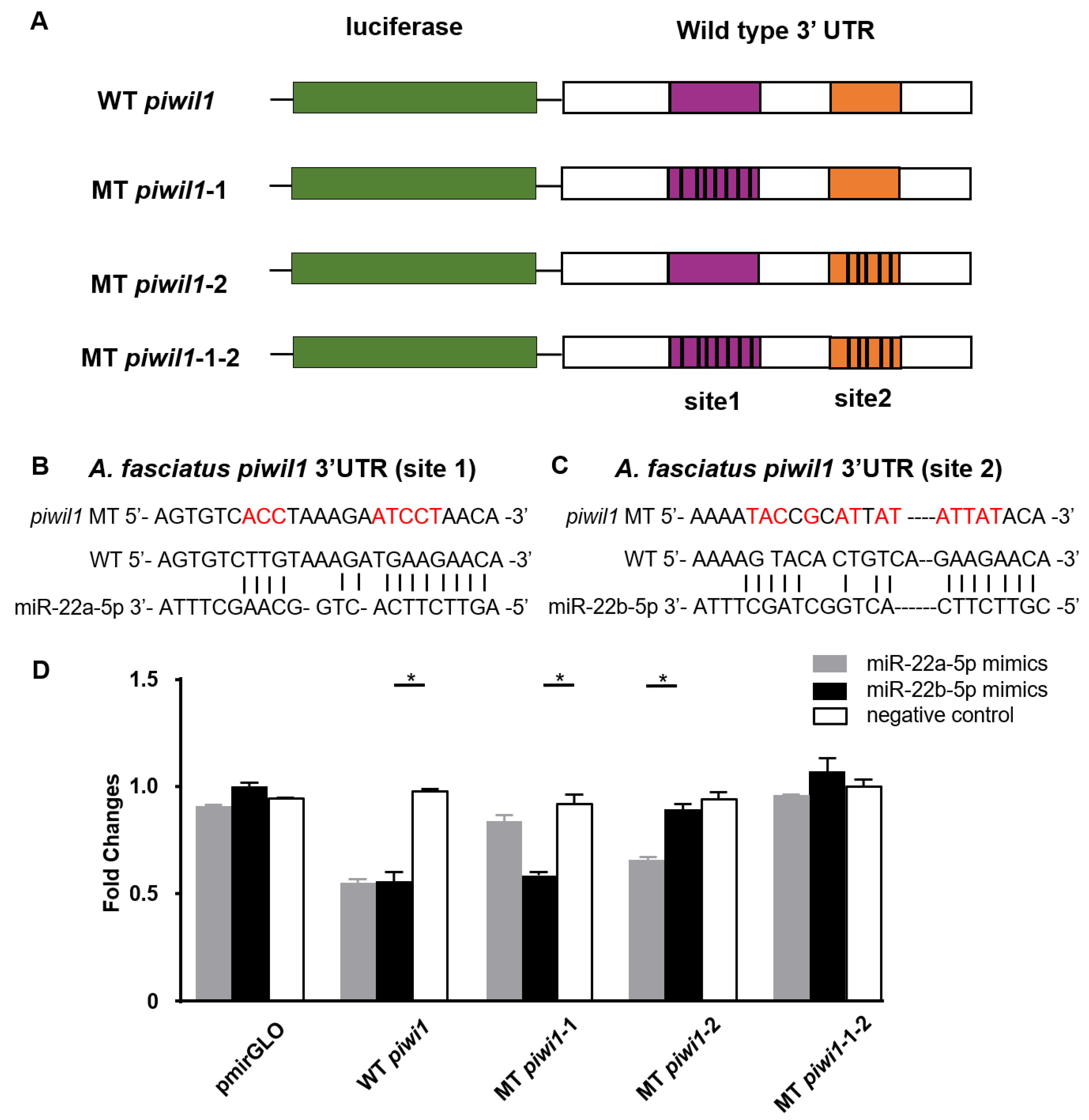
3.5. Piwil1 Is Regulated by miR-22a-5p and miR-22b-5p
3.6. Piwil2 Is Regulated by miR-10d-5p
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Xu, Z.; Wen, B.; Gao, J.; Chen, Z. Gonad-Specific Transcriptomes Reveal Differential Expression of Gene and miRNA Between Male and Female of the Discus Fish (Symphysodon aequifasciatus). Front. Physiol. 2020, 11, 754. [Google Scholar] [CrossRef] [PubMed]
- Iwakawa, H.O.; Tomari, Y. Life of RISC: Formation, action, and degradation of RNA-induced silencing complex. Mol. Cell 2022, 82, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hong, N.; Xu, H.; Song, J.; Hong, Y. Germline replacement by blastula cell transplantation in the fish medaka. Sci. Rep. 2016, 6, 29658. [Google Scholar] [CrossRef]
- Li, X.Y.; Gui, J.F. Diverse and variable sex determination mechanisms in vertebrates. Sci. China Life Sci. 2018, 61, 1503–1514. [Google Scholar] [CrossRef]
- Nanda, I.; Kondo, M.; Hornung, U.; Asakawa, S.; Winkler, C.; Shimizu, A.; Shan, Z.; Haaf, T.; Shimizu, N.; Shima, A.; et al. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc. Natl. Acad. Sci. USA 2002, 99, 11778–11783. [Google Scholar] [CrossRef]
- Myosho, T.; Otake, H.; Masuyama, H.; Matsuda, M.; Kuroki, Y.; Fujiyama, A.; Naruse, K.; Hamaguchi, S.; Sakaizumi, M. Tracing the emergence of a novel sex-determining gene in medaka, Oryzias luzonensis. Genetics 2012, 191, 163–170. [Google Scholar] [CrossRef]
- Whitfield, L.S.; Lovell-Badge, R.; Goodfellow, P.N. Rapid sequence evolution of the mammalian sex-determining gene SRY. Nature 1993, 364, 713–715. [Google Scholar] [CrossRef]
- Yerushalmi, G.M.; Salmon-Divon, M.; Ophir, L.; Yung, Y.; Baum, M.; Coticchio, G.; Fadini, R.; Mignini-Renzini, M.; Dal Canto, M.; Machtinger, R.; et al. Characterization of the miRNA regulators of the human ovulatory cascade. Sci. Rep. 2018, 8, 15605. [Google Scholar] [CrossRef] [PubMed]
- Mishima, T.; Takizawa, T.; Luo, S.S.; Ishibashi, O.; Kawahigashi, Y.; Mizuguchi, Y.; Ishikawa, T.; Mori, M.; Kanda, T.; Goto, T.; et al. MicroRNA (miRNA) cloning analysis reveals sex differences in miRNA expression profiles between adult mouse testis and ovary. Reproduction 2008, 136, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Presslauer, C.; Tilahun Bizuayehu, T.; Kopp, M.; Fernandes, J.M.; Babiak, I. Dynamics of miRNA transcriptome during gonadal development of zebrafish. Sci. Rep. 2017, 7, 43850. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Zhu, Y.; Wu, Y.; Yuan, C.; Chen, K.; Li, M. Identification and expression analysis of microRNAs in medaka gonads. Gene 2018, 646, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Hao, G.; Zhao, Z.; Wang, F.; Cao, J.; Yang, A. MicroRNA-224 regulates self-renewal of mouse spermatogonial stem cells via targeting DMRT1. J. Cell Mol. Med. 2016, 20, 1503–1512. [Google Scholar] [CrossRef]
- Guo, Y.; Wu, Y.; Shi, J.; Zhuang, H.; Ci, L.; Huang, Q.; Wan, Z.; Yang, H.; Zhang, M.; Tan, Y.; et al. miR-29a/b1 Regulates the Luteinizing Hormone Secretion and Affects Mouse Ovulation. Front. Endocrinol. 2021, 12, 636220. [Google Scholar] [CrossRef]
- Gay, S.; Bugeon, J.; Bouchareb, A.; Henry, L.; Delahaye, C.; Legeai, F.; Montfort, J.; Le Cam, A.; Siegel, A.; Bobe, J.; et al. MiR-202 controls female fecundity by regulating medaka oogenesis. PLoS Genet. 2018, 14, e1007593. [Google Scholar] [CrossRef]
- Shcherbata, H.R. miRNA functions in stem cells and their niches: Lessons from the Drosophila ovary. Curr. Opin. Insect Sci. 2019, 31, 29–36. [Google Scholar] [CrossRef]
- Guo, Q.; Jiang, Y.; Bai, H.; Chen, G.; Chang, G. miR-301a-5p Regulates TGFB2 during Chicken Spermatogenesis. Genes 2021, 12, 1695. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Q.; Hu, Y.; Xu, L.; Jiang, Y.; Zhang, C.; Ding, L.; Jiang, R.; Sun, J.; Sun, H.; et al. miR-181a increases FoxO1 acetylation and promotes granulosa cell apoptosis via SIRT1 downregulation. Cell Death Dis. 2017, 8, e3088. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.D.; Sheng, Z.; Hou, C.C.; Ni, J.; Han, Y.L.; Wang, Y.F.; Zhou, Y.; Fu, S.Y.; Zhu, J.Q. Molecular cloning, characterization and expression analysis of metallothionein in the liver of the teleost Acrossocheilus fasciatus exposed to cadmium chloride. Environ. Toxicol. Pharmacol. 2017, 53, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Guo, L.; Xiang, X.; Tao, J.; Chen, Y. Age and Growth of Acrossocheilus fasciatus (Barbinae, Cyprinidae) from the Puxi Stream in the Huangshan Mountain, China. J. Freshw. Ecol. 2011, 25, 79–83. [Google Scholar] [CrossRef]
- Tao, W.; Sun, L.; Shi, H.; Cheng, Y.; Jiang, D.; Fu, B.; Conte, M.A.; Gammerdinger, W.J.; Kocher, T.D.; Wang, D. Integrated analysis of miRNA and mRNA expression profiles in tilapia gonads at an early stage of sex differentiation. BMC Genom. 2016, 17, 328. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lin, G.; Fang, W.; Gao, D.; Huang, J.; Xie, J.; Lu, J. Identification and Comparison of microRNAs in the Gonad of the Yellowfin Seabream (Acanthopagrus latus). Int. J. Mol. Sci. 2020, 21, 5690. [Google Scholar] [CrossRef]
- Zeng, Q.; Hu, B.; Blanco, A.H.; Zhang, W.; Zhao, D.; Martinez, P.; Hong, Y. Full-Length Transcriptome Sequences Provide Insight into Hermaphroditism of Freshwater Pearl Mussel Hyriopsis schlegelii. Front. Genet. 2022, 13, 868742. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Zhang, H.; Cai, Q.; Tang, D.; Wang, G.; Liu, T.; Lv, B.; Wu, H. Integrated mRNA and miRNA expression profile analyses reveal the potential roles of sex-biased miRNA-mRNA pairs in gonad tissues of the Chinese concave-eared torrent frog (Odorrana tormota). J. Exp. Zool B Mol. Dev. Evol. 2019, 332, 69–80. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Duan, S.H.; Wang, G.L.; Li, J.L. Integrated mRNA and miRNA expression profile analysis of female and male gonads in Hyriopsis cumingii. Sci Rep. 2021, 11, 665. [Google Scholar] [CrossRef]
- Yue, Y.; Meng, Y.; Ma, H.; Hou, S.; Cao, G.; Hong, W.; Shi, Y.; Guo, P.; Liu, B.; Shi, F.; et al. A large family of Dscam genes with tandemly arrayed 5′ cassettes in Chelicerata. Nat. Commun. 2016, 7, 11252. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Friedlander, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [Green Version]
- John, B.; Sander, C.; Marks, D.S. Prediction of human microRNA targets. Methods Mol. Biol. 2006, 342, 101–113. [Google Scholar] [PubMed]
- Guo, S.; Zhong, Y.; Zhang, Y.; Zhu, Y.; Guo, J.; Fu, Y.; Li, M. Transcriptome analysis provides insights into long noncoding RNAs in medaka gonads. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 39, 100842. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, Y.; Zhu, T.; Song, P.; Guo, J.; Zhong, Y.; Gui, L.; Li, M. Vasa identifies germ cells in embryos and gonads of Oryzias celebensis. Gene 2022, 823, 146369. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhong, Y.; Guo, S.; Zhu, Y.; Guo, J.; Fu, Y.; Li, M. CircRNA profiling reveals circ880 functions as miR-375-3p sponge in medaka gonads. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 38, 100797. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zhang, L.; Chen, X. Differential expression analysis of Paralichthys olivaceus microRNAs in adult ovary and testis by deep sequencing. Gen. Comp. Endocrinol. 2014, 204, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zhong, H.; Zhou, Y.; Yu, F.; Gao, Y.; Luo, Y.; Tang, Z.; Guo, Z.; Guo, E.; Gan, X.; et al. Identification and characterization of microRNAs in ovary and testis of Nile tilapia (Oreochromis niloticus) by using solexa sequencing technology. PLoS ONE 2014, 9, e86821. [Google Scholar] [CrossRef]
- Xu, H.; Li, M.; Gui, J.; Hong, Y. Cloning and expression of medaka dazl during embryogenesis and gametogenesis. Gene Expr. Patterns 2007, 7, 332–338. [Google Scholar] [CrossRef]
- Li, M.; Zhu, F.; Li, Z.; Hong, N.; Hong, Y. Dazl is a critical player for primordial germ cell formation in medaka. Sci. Rep. 2016, 6, 28317. [Google Scholar] [CrossRef]
- Hasuwa, H.; Iwasaki, Y.W.; Au Yeung, W.K.; Ishino, K.; Masuda, H.; Sasaki, H.; Siomi, H. Production of functional oocytes requires maternally expressed PIWI genes and piRNAs in golden hamsters. Nat. Cell Biol. 2021, 23, 1002–1012. [Google Scholar] [CrossRef]
- Zhou, Y.; Sun, W.; Cai, H.; Bao, H.; Zhang, Y.; Qian, G.; Ge, C. The Role of Anti-Mullerian Hormone in Testis Differentiation Reveals the Significance of the TGF-beta Pathway in Reptilian Sex Determination. Genetics 2019, 213, 1317–1327. [Google Scholar] [CrossRef]
- Kobayashi, T.; Matsuda, M.; Kajiura-Kobayashi, H.; Suzuki, A.; Saito, N.; Nakamoto, M.; Shibata, N.; Nagahama, Y. Two DM domain genes, DMY and DMRT1, involved in testicular differentiation and development in the medaka, Oryzias latipes. Dev. Dyn. 2004, 231, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, C.Y.; Zhao, Y.; Dean, J. FIGLA, LHX8 and SOHLH1 transcription factor networks regulate mouse oocyte growth and differentiation. Nucleic Acids Res. 2020, 48, 3525–3541. [Google Scholar] [CrossRef]
- Rong, Y.; Ji, S.Y.; Zhu, Y.Z.; Wu, Y.W.; Shen, L.; Fan, H.Y. ZAR1 and ZAR2 are required for oocyte meiotic maturation by regulating the maternal transcriptome and mRNA translational activation. Nucleic Acids Res. 2019, 47, 11387–11402. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Zhu, S.; Zhang, H.; Li, C.; Qiu, D.; Ge, J.; Guo, X.; Wang, Q. Mitofusin1 in oocyte is essential for female fertility. Redox Biol. 2019, 21, 101110. [Google Scholar] [CrossRef]
- Zhou, H.; Whitworth, C.; Pozmanter, C.; Neville, M.C.; Van Doren, M. Doublesex regulates fruitless expression to promote sexual dimorphism of the gonad stem cell niche. PLoS Genet. 2021, 17, e1009468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, W.; Jin, Y.; Jia, P.; Jia, K.; Yi, M. MiR-202-5p is a novel germ plasm-specific microRNA in zebrafish. Sci. Rep. 2017, 7, 7055. [Google Scholar] [CrossRef] [PubMed]
- Skaftnesmo, K.O.; Edvardsen, R.B.; Furmanek, T.; Crespo, D.; Andersson, E.; Kleppe, L.; Taranger, G.L.; Bogerd, J.; Schulz, R.W.; Wargelius, A. Integrative testis transcriptome analysis reveals differentially expressed miRNAs and their mRNA targets during early puberty in Atlantic salmon. BMC Genom. 2017, 18, 801. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xu, D.; Xu, Y.; Chen, L.; Li, C.; Dai, X.; Zhang, L.; Zheng, L. MicroRNA-141-3p targets DAPK1 and inhibits apoptosis in rat ovarian granulosa cells. Cell Biochem. Funct. 2017, 35, 197–201. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Sobinoff, A.P.; Sutherland, J.M.; McLaughlin, E.A. Intracellular signalling during female gametogenesis. Mol. Hum. Reprod. 2013, 19, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.Y.; Dai, X.L.; Ran, X.Q.; Cen, Y.X.; Niu, X.; Li, S.; Huang, S.H.; Wang, J.F. Identification and profile of microRNAs in Xiang pig testes in four different ages detected by Solexa sequencing. Theriogenology 2018, 117, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Bi, L.; Sun, L.; Jin, Z.; Zhang, S.; Shen, Z. MicroRNA-10a/b are regulators of myeloid differentiation and acute myeloid leukemia. Oncol. Lett. 2018, 15, 5611–5619. [Google Scholar] [CrossRef]
- Zuo, H.; Liu, X.; Luo, M.; Yang, L.; Zhu, Z.; Weng, S.; He, J.; Xu, X. miR-10c Facilitates White Spot Syndrome Virus Infection by Targeting Toll3 in Litopenaeus vannemei. Front. Immunol. 2021, 12, 733730. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Liu, Q.; Zhu, L.J.; Gao, H.; Cui, M.; Liu, J.; Zhao, P.; Liu, J.; Chen, L.; et al. Conserved microRNA mediates heating tolerance in germ cells versus surrounding somatic cells. RNA Biol. 2019, 16, 1494–1503. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, W.; He, J.; Yaqoob, M.A.; Gui, L.; Ren, J.; Li, J.; Li, M. Integrated mRNA and miRNA Expression Profile Analysis of Female and Male Gonads in Acrossocheilus fasciatus. Biology 2022, 11, 1296. https://doi.org/10.3390/biology11091296
Wei W, He J, Yaqoob MA, Gui L, Ren J, Li J, Li M. Integrated mRNA and miRNA Expression Profile Analysis of Female and Male Gonads in Acrossocheilus fasciatus. Biology. 2022; 11(9):1296. https://doi.org/10.3390/biology11091296
Chicago/Turabian StyleWei, Wenbo, Jiamei He, Muhammad Amjad Yaqoob, Lang Gui, Jianfeng Ren, Jiale Li, and Mingyou Li. 2022. "Integrated mRNA and miRNA Expression Profile Analysis of Female and Male Gonads in Acrossocheilus fasciatus" Biology 11, no. 9: 1296. https://doi.org/10.3390/biology11091296
APA StyleWei, W., He, J., Yaqoob, M. A., Gui, L., Ren, J., Li, J., & Li, M. (2022). Integrated mRNA and miRNA Expression Profile Analysis of Female and Male Gonads in Acrossocheilus fasciatus. Biology, 11(9), 1296. https://doi.org/10.3390/biology11091296