


Preserving Pure Siamese Crocodile Populations: A Comprehensive Approach Using Multi-Genetic Tools

 ,
, 
 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collection and DNA Extraction
2.2. Development of DArT Arrays
2.3. Genotyping Assay and Marker Selection
2.4. Genetic Diversity and Population Structure Analyses
2.5. Species Delimitation
2.6. Homology Searching
2.7. Functional Annotation and Gene Ontology of Siamese and Saltwater Crocodiles
2.8. Identification of Conserved and Species-Diagnostic SNPs
2.9. Validation Assays
3. Results
3.1. Genotyping Variation, Inbreeding Coefficient, and Relatedness
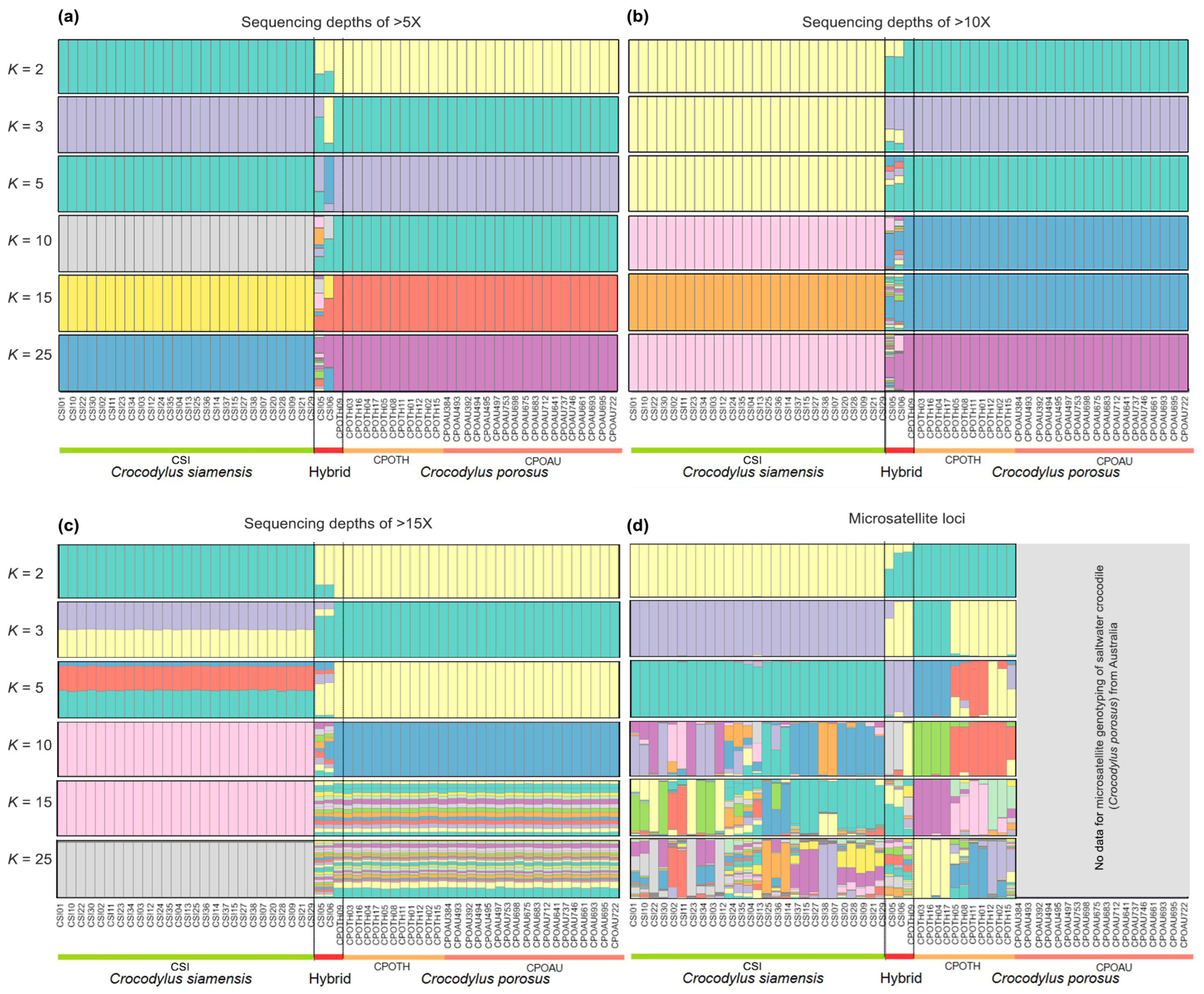
3.2. Clustering of Siamese and Saltwater Crocodile Populations
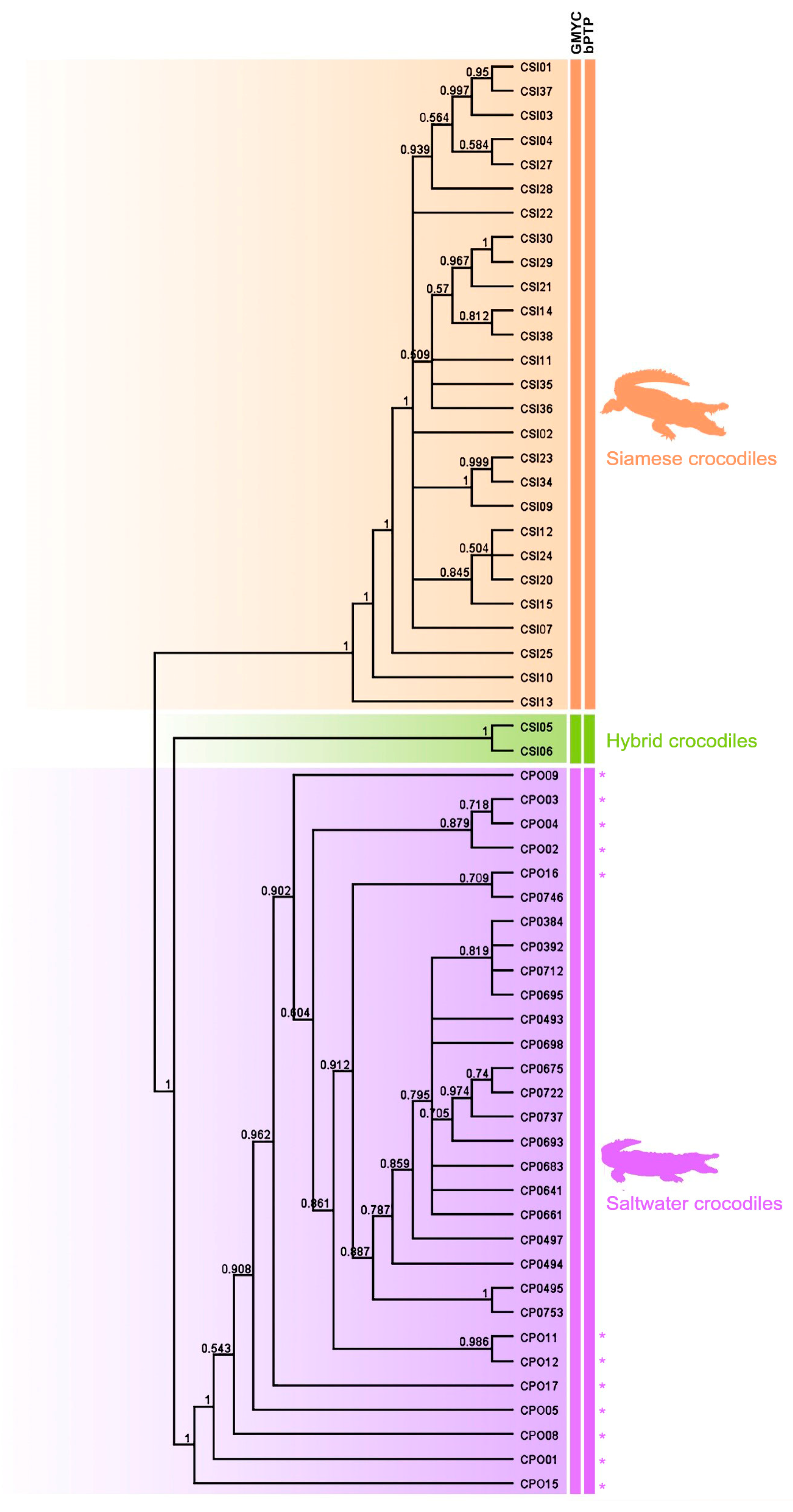
3.3. Species Delimitation
3.4. Identification of Conserved and Species-Diagnostic SNPs
3.5. Homology of Putative Species-Diagnostic Loci
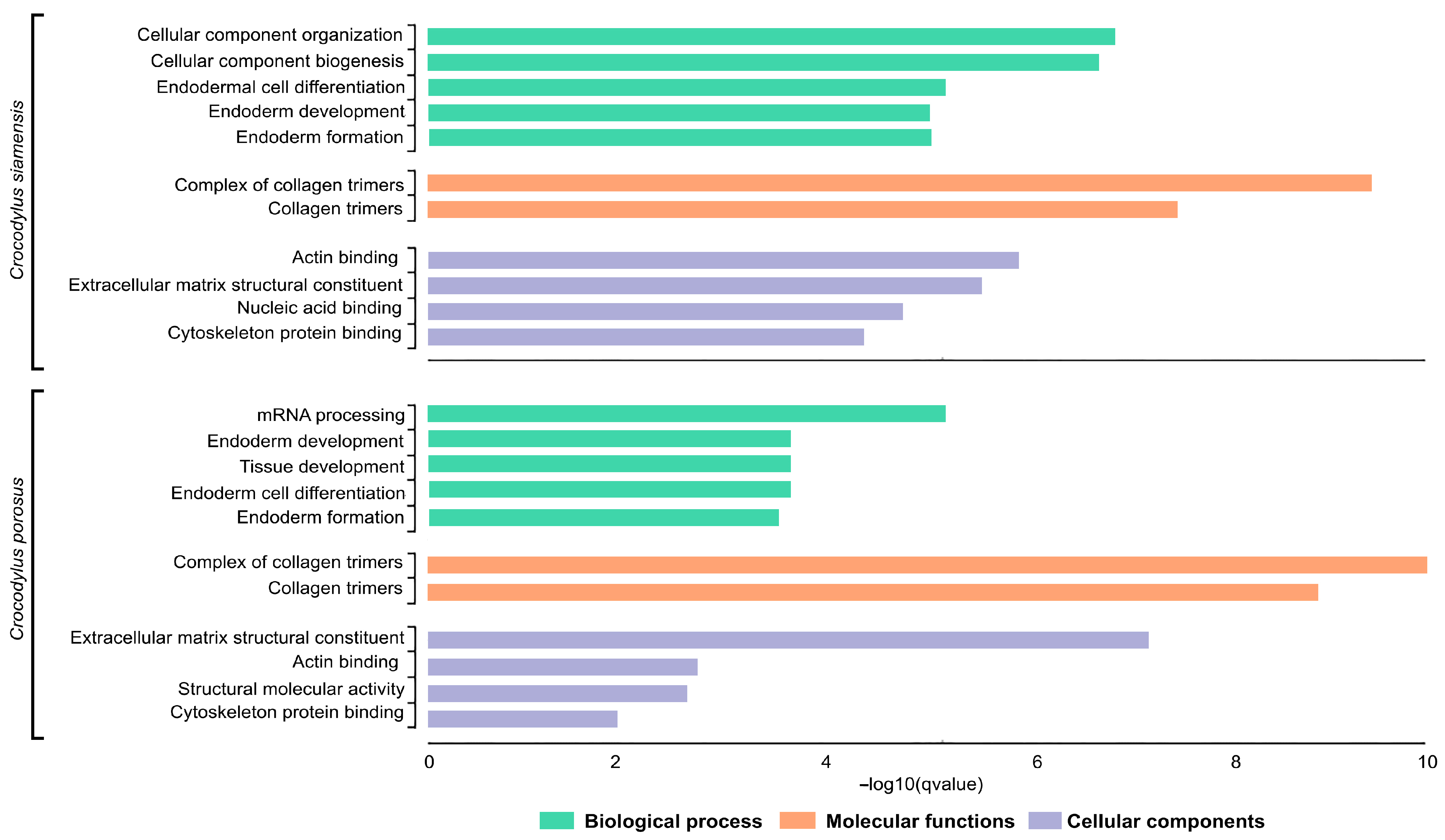
3.6. Functional Classification and Enrichment Analysis of Siamese and Saltwater Crocodile Loci
3.7. Validation of Species-Diagnostic Loci with DNA Markers
4. Discussion
4.1. Can the Evolutionary Relationships within and between Siamese and Saltwater Crocodiles Be Clarified?
4.2. Can Parental and Hybrid Individuals Be Differentiated into Separate Lineages?
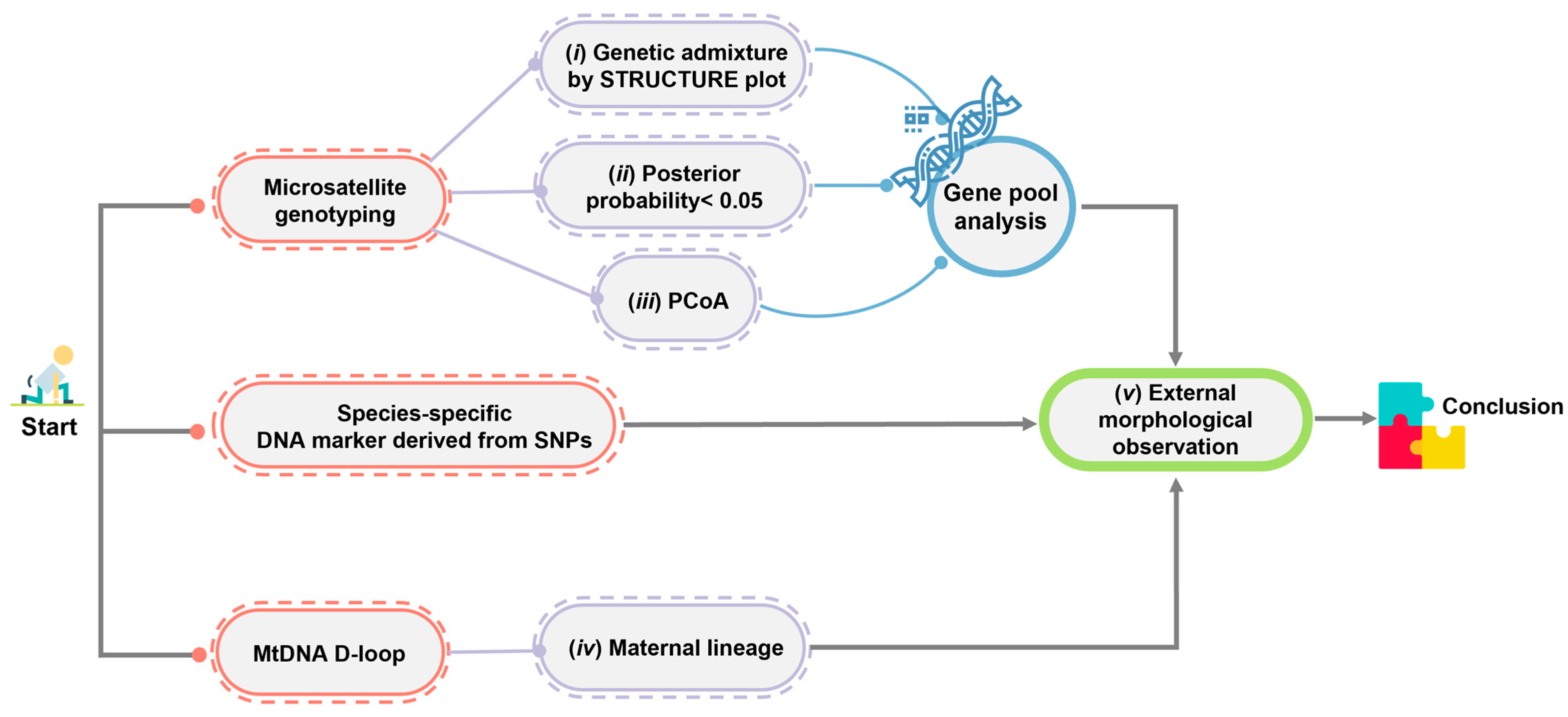
4.3. Enhancing Siamese Crocodile Identification through a Combination of Microsatellite Genotyping, Mitochondrial DNA D-Loop Sequencing, and PCR-Based Species-Diagnostic Markers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef]
- Jackiw, R.N.; Mandil, G.; Hager, H.A. A framework to guide the conservation of species hybrids based on ethical and ecological considerations. Conserv. Biol. 2015, 29, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Lapbenjakul, S.; Thapana, W.; Twilprawat, P.; Muangmai, N.; Kanchanaketu, T.; Temsiripong, Y.; Unajak, S.; Peyachoknagul, S.; Srikulnath, K. High genetic diversity and demographic history of captive Siamese and Saltwater crocodiles suggest the first step toward the establishment of a breeding and reintroduction program in Thailand. PLoS ONE 2017, 12, e0184526. [Google Scholar] [CrossRef] [PubMed]
- Ariyaraphong, N.; Wongloet, W.; Wattanadilokchatkun, P.; Panthum, T.; Singchat, W.; Thong, T.; Lisachov, A.; Ahmad, S.F.; Muangmai, N.; Han, K.; et al. Should the identification guidelines for Siamese crocodiles be revised? differing post-occipital scute scale numbers show phenotypic variation does not result from hybridization with saltwater crocodiles. Biology 2023, 12, 535. [Google Scholar] [CrossRef]
- Srikulnath, K.; Thapana, W.; Muangmai, N. Role of chromosome changes in Crocodylus evolution and diversity. Genom. Inform. 2015, 13, 102–111. [Google Scholar] [CrossRef]
- Milián-García, Y.; Ramos-Targarona, R.; Pérez-Fleitas, E.; Sosa-Rodríguez, G.; Guerra-Manchena, L.; Alonso-Tabet, M.; Espinosa-López, G.; Russello, M.A. Genetic evidence of hybridization between the critically endangered Cuban crocodile and the American crocodile: Implications for population history and in situ/ex situ conservation. Heredity 2015, 114, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Ihlow, F.; Bonke, R.; Hartmann, T.; Geissler, P.; Behler, N.; Rödder, D. Habitat suitability, coverage by protected areas and population connectivity for the Siamese crocodile Crocodylus siamensis Schneider, 1801. Aquat. Conserv. Mar. Freshw. Ecosyst. 2015, 25, 544–554. [Google Scholar] [CrossRef]
- Webb, G.J.W.; Manolis, C.; Brien, M.L.; Balaguera-Reina, S.A.; Isberg, S. Crocodylus porosus. In The IUCN Red List of Threatened Species. 2021, p. e. T5668A3047556. Available online: https://doi.org/10.2305/IUCN.UK.2021-2.RLTS.T5668A3047556.en (accessed on 4 October 2023).
- Srikulnath, K.; Thongpan, A.; Suputtitada, S.; Apisitwanich, S. New haplotype of the complete mitochondrial genome of Crocodylus siamensis and its species-specific DNA markers: Distinguishing C. siamensis from C. porosus in Thailand. Mol. Biol. Rep. 2012, 39, 4709–4717. [Google Scholar] [CrossRef]
- Suvanakorn, P.; Youngprapakorn, C. Crocodile farming in Thailand. In Wildlife Management: Crocodiles and Alligators; Webb, G.J.W., Manolis, S.C., Whitehead, P.J., Eds.; Surrey Beatty and Sons, Pty. Ltd.: Chipping Norton, NSW, Australia, 1987; pp. 341–343. [Google Scholar]
- Chiu, M.C.; Nukazawa, K.; Resh, V.H.; Watanabe, K. Environmental effects, gene flow and genetic drift: Unequal influences on genetic structure across landscapes. J. Biogeogr. 2023, 50, 352–364. [Google Scholar] [CrossRef]
- Laopichienpong, N.; Kraichak, E.; Singchat, W.; Sillapaprayoon, S.; Muangmai, N.; Suntrarachun, S.; Baicharoen, S.; Peyachoknagul, S.; Chanhome, L.; Ezaz, T.; et al. Genome-wide SNP analysis of Siamese cobra (Naja kaouthia) reveals the molecular basis of transitions between Z and W sex chromosomes and supports the presence of an ancestral super-sex chromosome in amniotes. Genomics 2021, 113, 624–636. [Google Scholar] [CrossRef]
- Gamble, T.; Zarkower, D. Identification of sex-specific molecular markers using restriction site-associated DNA sequencing. Mol. Ecol. Resour. 2014, 14, 902–913. [Google Scholar] [CrossRef]
- Fukuda, Y.; Moritz, C.; Jang, N.C.; Webb, G.; Campbell, H.; Christian, K.; Lindner, G.; Banks, S. Environmental resistance and habitat quality influence dispersal of the saltwater crocodile. Mol. Ecol. 2022, 31, 1076–1092. [Google Scholar] [CrossRef] [PubMed]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity arrays: A solid state technology for sequence information independent genotyping. Nucleic Acids Res. 2001, 29, e25. [Google Scholar] [CrossRef]
- Kilian, A.; Wenzl, P.; Huttner, E.; Carling, J.; Xia, L.; Blois, H.; Caig, V.; Heller-Uszynska, K.; Jaccoud, D.; Hopper, C.; et al. Diversity arrays technology: A generic genome profiling technology on open platforms. Methods Mol. Biol. 2012, 888, 67–89. [Google Scholar] [CrossRef]
- Ren, R.; Ray, R.; Li, P.; Xu, J.; Zhang, M.; Liu, G.; Yao, X.; Kilian, A.; Yang, X. Construction of a high-density DArTseq SNP-based genetic map and identification of genomic regions with segregation distortion in a genetic population derived from a cross between feral and cultivated-type watermelon. Mol. Genet. Genom. 2015, 290, 1457–1470. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Peakall, R.O.D.; Smouse, P.E. GENALEX 6: Genetic analysis in excel. population genetic software for teaching and research. Mol. Ecol. 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Jombart, T.; Collins, C. A Tutorial for Discriminant Analysis of Principal Components (DAPC) Using Adegenet 2.0.0; Imperial College London, MRC Centre for Outbreak Analysis and Modelling: London, UK, 2015. [Google Scholar]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef]
- Pickrell, J.; Pritchard, J. Inference of population splits and mixtures from genome-wide allele frequency data. PLoS Genet. 2012, 8, e1002967. [Google Scholar] [CrossRef] [PubMed]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A.P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Lima, T.; Auchincloss, A.H.; Coudert, E.; Keller, G.; Michoud, K.; Rivoire, C.; Bulliard, V.; de Castro, E.; Lachaize, C.; Baratin, D.; et al. HAMAP: A database of completely sequenced microbial proteome sets and manually curated microbial protein families in UniProtKB/Swiss-Prot. Nucleic Acids Res. 2009, 37, D471–D478. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Chen, Y.E.; Song, D.; McDermott, M.; Woyshner, K.; Manousopoulou, A.; Wang, N.; Li, W.; Wang, L.; Li, J.J. Clipper: P-value-free FDR control on high-throughput data from two conditions. Genome Biol. 2021, 22, 288. [Google Scholar] [CrossRef]
- Al-Qurainy, F.; Alshameri, A.; Gaafar, A.R.; Khan, S.; Nadeem, M.; Alameri, A.A.; Tarroum, M.; Ashraf, M. Comprehensive stress-based de novo transcriptome assembly and annotation of guar (Cyamopsis tetragonoloba (L.) Taub.): An important industrial and forage crop. Int. J. Genom. 2019, 2019, 7295859. [Google Scholar] [CrossRef] [PubMed]
- Platt, S.G.; Lynam, A.J.; Temsiripong, Y.; Kampanakngarn, M. Occurrence of the Siamese crocodile (Crocodylus siamensis) in Kaeng Krachan National Park, Thailand. Nat. Hist. Bull. Siam Soc. 2002, 50, 7–14. [Google Scholar]
- Chattopadhyay, B.; Garg, K.M.; Soo, Y.J.; Low, G.W.; Frechette, J.L.; Rheindt, F.E. Conservation genomics in the fight to help the recovery of the critically endangered Siamese crocodile Crocodylus siamensis. Mol. Ecol. 2019, 28, 936–950. [Google Scholar] [CrossRef] [PubMed]
- Melville, J.; Haines, M.L.; Boysen, K.; Hodkinson, L.; Kilian, A.; Smith Date, K.L.; Potvin, D.A.; Parris, K.M. Identifying hybridization and admixture using SNPs: Application of the DArTseq platform in phylogeographic research on vertebrates. R. Soc. Open Sci. 2017, 4, 161061. [Google Scholar] [CrossRef] [PubMed]
- De Meeûs, T. Revisiting FIS, FST, Wahlund Effects, and Null Alleles. J. Hered. 2018, 109, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Reddy, U.K.; Abburi, L.; Abburi, V.L.; Saminathan, T.; Cantrell, R.; Vajja, V.G.; Reddy, R.; Tomason, Y.R.; Levi, A.; Wehner, T.C.; et al. A genome-wide scan of selective sweeps and association mapping of fruit traits using microsatellite markers in watermelon. J. Hered. 2015, 106, 166–176. [Google Scholar] [CrossRef]
- Kaiser, S.A.; Taylor, S.A.; Chen, N.; Sillett, T.S.; Bondra, E.R.; Webster, M.S. A comparative assessment of SNP and microsatellite markers for assigning parentage in a socially monogamous bird. Mol. Ecol. Resour. 2017, 17, 183–193. [Google Scholar] [CrossRef]
- Ling, C.; Lixia, W.; Rong, H.; Fujun, S.; Wenping, Z.; Yao, T.; Yaohua, Y.; Bo, Z.; Liang, Z. Comparative analysis of microsatellite and SNP markers for parentage testing in the golden snub-nosed monkey (Rhinopithecus roxellana). Conserv. Genet. Res 2020, 12, 611–620. [Google Scholar] [CrossRef]
- Tereba, A.; Konecka, A. Comparison of microsatellites and SNP markers in genetic diversity level of two Scots pine stands. Environ. Sci. Proc. 2020, 3, 4. [Google Scholar] [CrossRef]
- Qu, L.; Li, X.; Xu, G.; Chen, K.; Yang, H.; Zhang, L.; Wu, G.; Hou, Z.; Xu, G.; Yang, N. Evaluation of genetic diversity in Chinese indigenous chicken breeds using microsatellite markers. Sci. China Life Sci. 2006, 49, 332–341. [Google Scholar] [CrossRef]
- Knowles, L.L. The burgeoning field of statistical phylogeography. J. Evol. Biol. 2004, 17, 1–10. [Google Scholar] [CrossRef]
- Garrick, R.C.; Bonatelli, I.A.; Hyseni, C.; Morales, A.; Pelletier, T.A.; Perez, M.F.; Rice, E.; Satler, J.D.; Symula, R.E.; Thomé, M.T.C.; et al. The evolution of phylogeographic data sets. Mol. Ecol. 2015, 24, 1164–1171. [Google Scholar] [CrossRef]
- Caldwell, J. World Trade in Crocodilian Skins 2008–2010; UNEP-WCMC: Cambridge, CA, USA, 2010. [Google Scholar]
- Getpech, Y. Crocodile conservation and captive breeding in Thailand. In Proceedings of the 1st Regional Species Meeting of the IUCN-SSC Crocodile Specialist Group, Bangkok, Thailand, 4–6 April 2011; pp. 27–28. [Google Scholar]
- Caldwell, J. World Trade in Crocodilian Skins 2018–2020; UNEP-WCMC: Cambridge, CA, USA, 2022. [Google Scholar]
- Fitzsimmons, N.N.; Buchan, J.C.; Lam, P.V.; Polet, G.; Hung, T.T.; Thang, N.Q.; Gratten, J. Identification of purebred Crocodylus siamensis for reintroduction in Vietnam. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2002, 294, 373–381. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Hohenlohe, P.A.; Luikart, G. Genomics and the future of conservation genetics. Nat. Rev. Genet. 2010, 11, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Suchan, T.; Pitteloud, C.; Gerasimova, N.S.; Kostikova, A.; Schmid, S.; Arrigo, N.; Pajkovic, M.; Ronikier, M.; Alvarez, N. Hybridization capture using RAD probes (hyRAD), a new tool for performing genomic analyses on collection specimens. PLoS ONE 2016, 11, e0151651. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Locus Id | Sequence 5′–3′ | Product Size | A Positive Band on the Target Species |
|---|---|---|---|---|
| CST09-F | 23111606 | ACTGCAGACTGAGTGAGTTTC | 320 bp | Crocodylus siamensis |
| CST09-R | 23111606 | CTGTGCCTGAATTGACTGCCA | ||
| CPT08-F | 34606790 | TTGCCCTTAGCCCCAGTCTA | 450 bp | Crocodylus siamensis |
| CPT08-R | 34606790 | TACTGGGAGGCATCAAACCA | ||
| CST01-F | 23113297 | AACATTGCTCTGGATGGCCTT | 400 bp | Crocodylus porosus |
| CST01-R | 23113297 | CGGAAATGCCAGCTTTTAGT |
| Population | N | Na | Ne | I | Ho | He | PIC | F | |
|---|---|---|---|---|---|---|---|---|---|
| CSI 1 | Mean | 29 | 1.511 | 1.095 | 0.147 | 0.005 | 0.079 | 0.058 | 0.931 |
| S.E. | 0 | 0.007 | 0.001 | 0.002 | 0.000 | 0.001 | 0.090 | 0.004 | |
| CPOTH 2 | Mean | 12 | 1.511 | 1.194 | 0.217 | 0.020 | 0.133 | 0.000 | 0.907 |
| S.E. | 0 | 0.007 | 0.003 | 0.003 | 0.002 | 0.002 | 0.000 | 0.005 | |
| CPOAU 3 | Mean | 18 | 0.891 | 0.891 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| S.E. | 0 | 0.004 | 0.004 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| All Population | Mean | 59 | 1.511 | 1.051 | 0.096 | 0.002 | 0.046 | 0.044 | 0.934 |
| S.E. | 0 | 0.021 | 0.001 | 0.001 | 0.000 | 0.001 | 0.046 | 0.004 |
| Population 1 | Population 2 | df | SE | t-Test | p-Value | |
|---|---|---|---|---|---|---|
| Observed heterozygosity (Ho) | CSI 1 | CPOTH 2 | 0.015 | 0.002 | −7.5 | <0.05 |
| CSI 1 | CPOAU 3 | 0.005 | 0 | N/A | N/A | |
| CPOTH 2 | CPOAU 3 | 0.02 | 0.002 | 10 | <0.05 | |
| Expected heterozygosity (He) | CSI 1 | CPOTH 2 | −0.054 | 0.002 | −24.150 | <0.05 |
| CSI 1 | CPOAU 3 | N/A | N/A | N/A | N/A | |
| CPOTH 2 | CPOAU 3 | N/A | N/A | N/A | N/A |
| Population | Ho | He | df | t-Test | p-Value |
|---|---|---|---|---|---|
| CSI 1 | 0.005 ± 0.000 | 0.079 ± 0.001 | −0.074 | −74 | <0.05 |
| CPOTH 2 | 0.020 ± 0.002 | 0.133 ± 0.002 | −0.113 | −39.95 | <0.05 |
| CPOAU 3 | 0.000 ± 0.000 | 0.000 ± 0.000 | N/A | N/A | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panthum, T.; Ariyaraphong, N.; Wongloet, W.; Wattanadilokchatkun, P.; Laopichienpong, N.; Rasoarahona, R.; Singchat, W.; Ahmad, S.F.; Kraichak, E.; Muangmai, N.; et al. Preserving Pure Siamese Crocodile Populations: A Comprehensive Approach Using Multi-Genetic Tools. Biology 2023, 12, 1428. https://doi.org/10.3390/biology12111428
Panthum T, Ariyaraphong N, Wongloet W, Wattanadilokchatkun P, Laopichienpong N, Rasoarahona R, Singchat W, Ahmad SF, Kraichak E, Muangmai N, et al. Preserving Pure Siamese Crocodile Populations: A Comprehensive Approach Using Multi-Genetic Tools. Biology. 2023; 12(11):1428. https://doi.org/10.3390/biology12111428
Chicago/Turabian StylePanthum, Thitipong, Nattakan Ariyaraphong, Wongsathit Wongloet, Pish Wattanadilokchatkun, Nararat Laopichienpong, Ryan Rasoarahona, Worapong Singchat, Syed Farhan Ahmad, Ekaphan Kraichak, Narongrit Muangmai, and et al. 2023. "Preserving Pure Siamese Crocodile Populations: A Comprehensive Approach Using Multi-Genetic Tools" Biology 12, no. 11: 1428. https://doi.org/10.3390/biology12111428
APA StylePanthum, T., Ariyaraphong, N., Wongloet, W., Wattanadilokchatkun, P., Laopichienpong, N., Rasoarahona, R., Singchat, W., Ahmad, S. F., Kraichak, E., Muangmai, N., Duengkae, P., Fukuda, Y., Banks, S., Temsiripong, Y., Ezaz, T., & Srikulnath, K. (2023). Preserving Pure Siamese Crocodile Populations: A Comprehensive Approach Using Multi-Genetic Tools. Biology, 12(11), 1428. https://doi.org/10.3390/biology12111428