
Morphogenic Effect of Exogenous Glucocorticoid Hormones in the Girardia tigrina Planarian (Turbellaria, Tricladida)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
- (1)
- To study the effect of hydrocortisone in the concentration range from 10−3 M to 10−6 M and methylprednisolone in the concentration range from 10−4 M to 10−6 M on the reparative regeneration process of the head and tail planarian blastemas (new unpigmented tissue formed at the regeneration site);
- (2)
- To study the effect of exogenous steroid hormones hydrocortisone and methylprednisolone on the proliferation of planarian neoblast stem cells;
- (3)
- To search for homologs of glucocorticoid receptor proteins in a closely related species Schmidtea mediterranea, with fully sequenced and annotated genome.
2. Materials and Methods
2.1. The Object
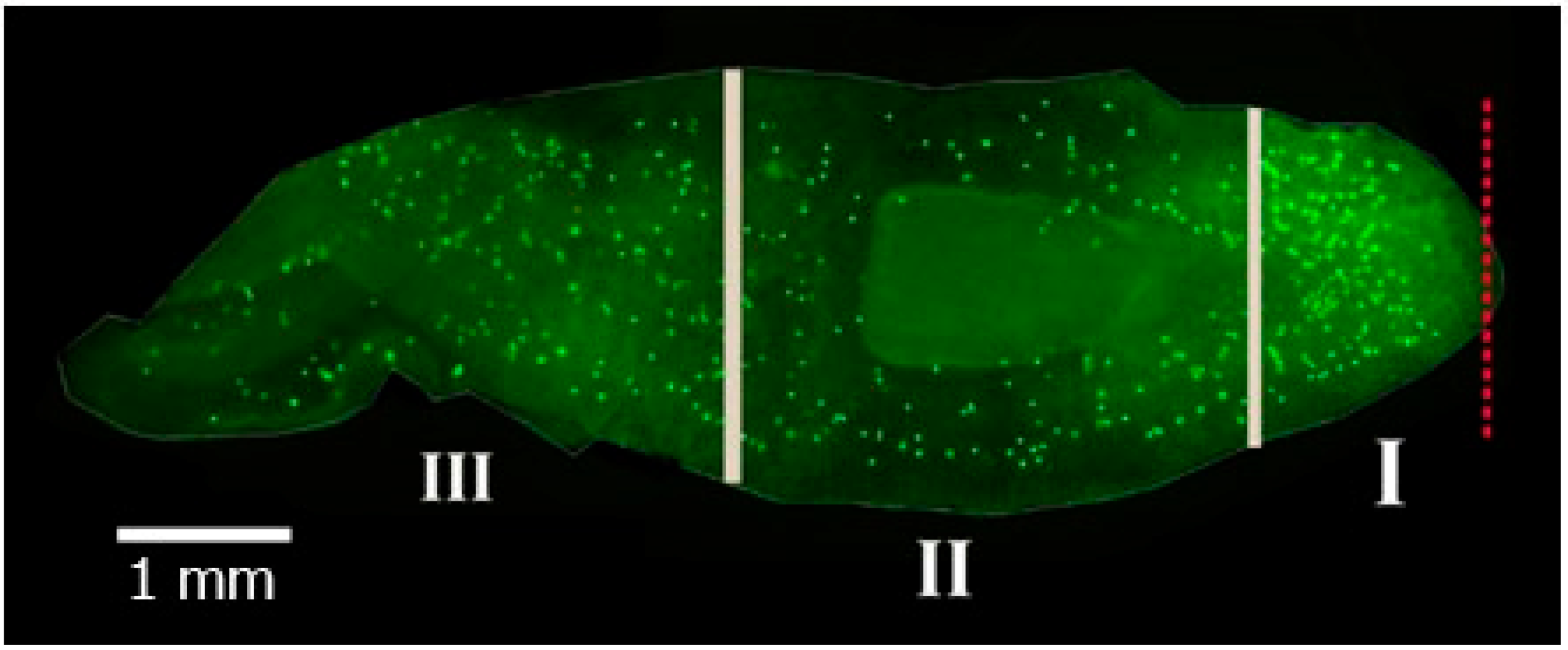
2.2. Morphometric Analysis
2.3. Immunohistochemical Analysis
2.4. The Search for Homologues of Receptor Proteins
2.5. Statistical Data Processing
3. Results
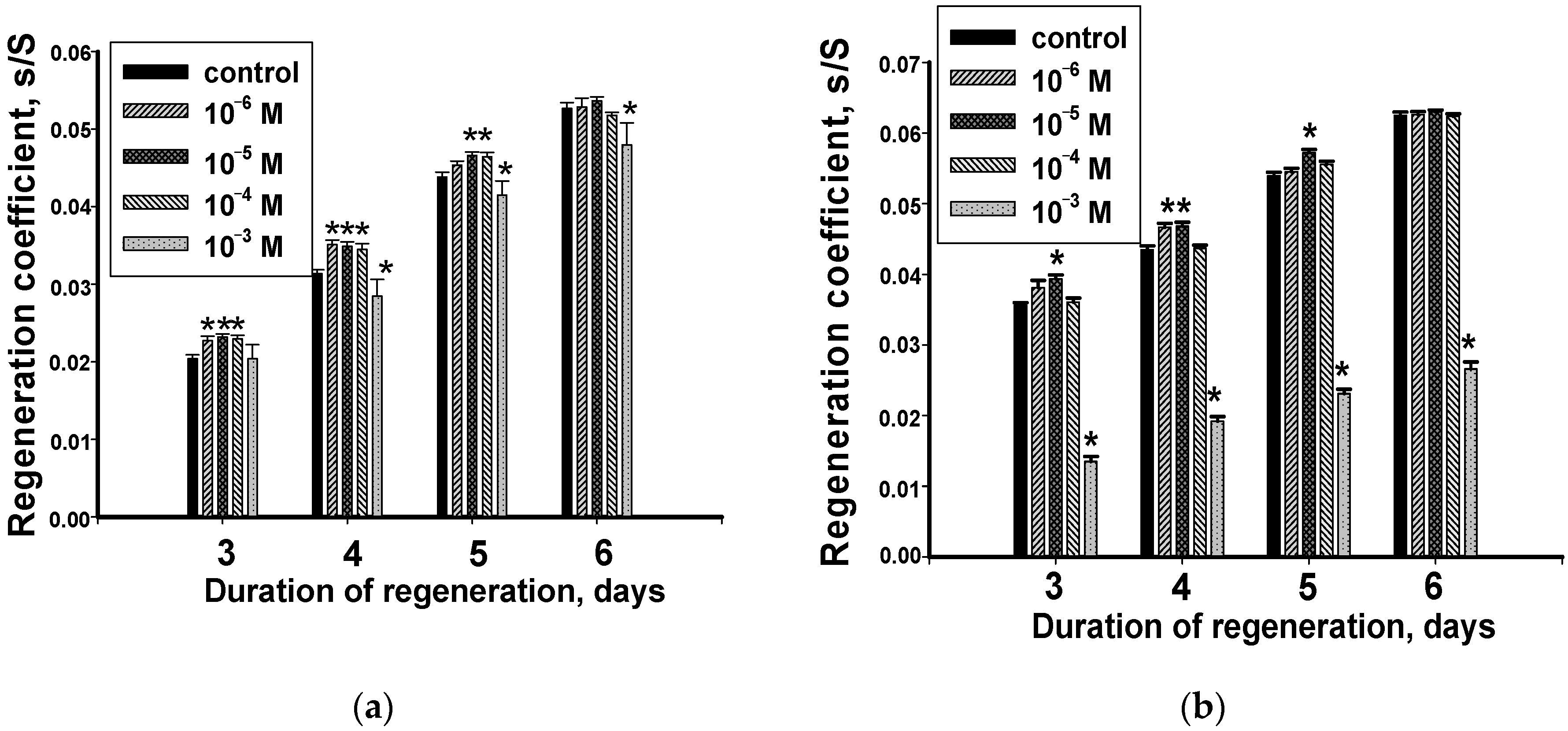
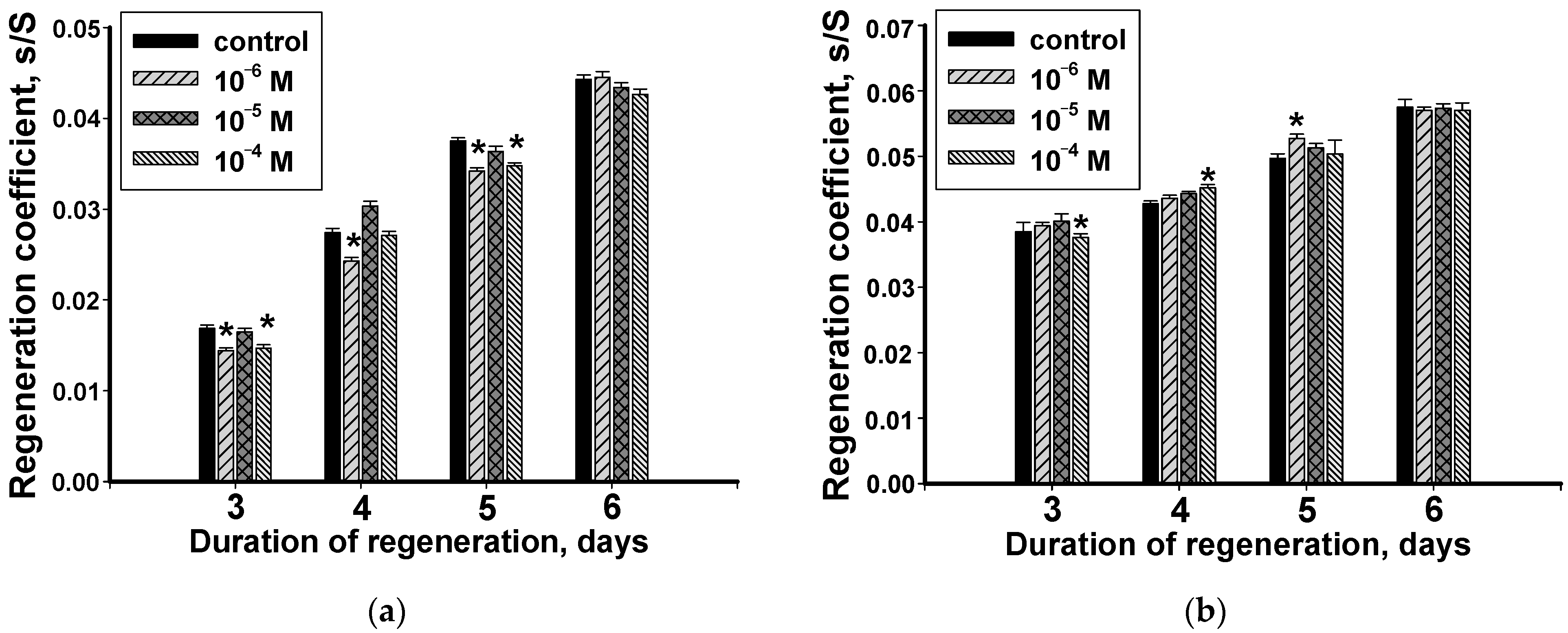
3.1. Dynamics of Changes in the Regeneration Coefficient of the Regenerating Head and Tail Blastema with Hydrocortisone
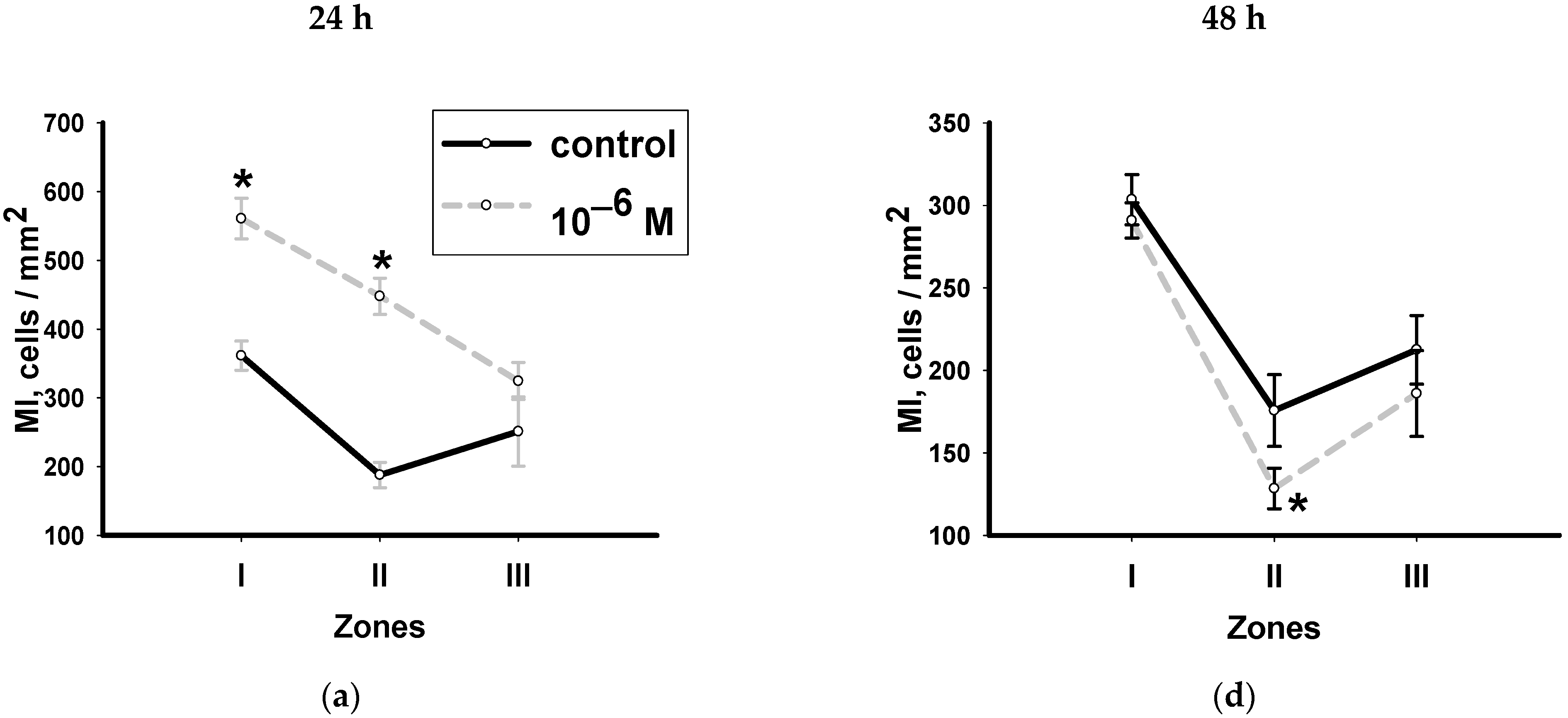
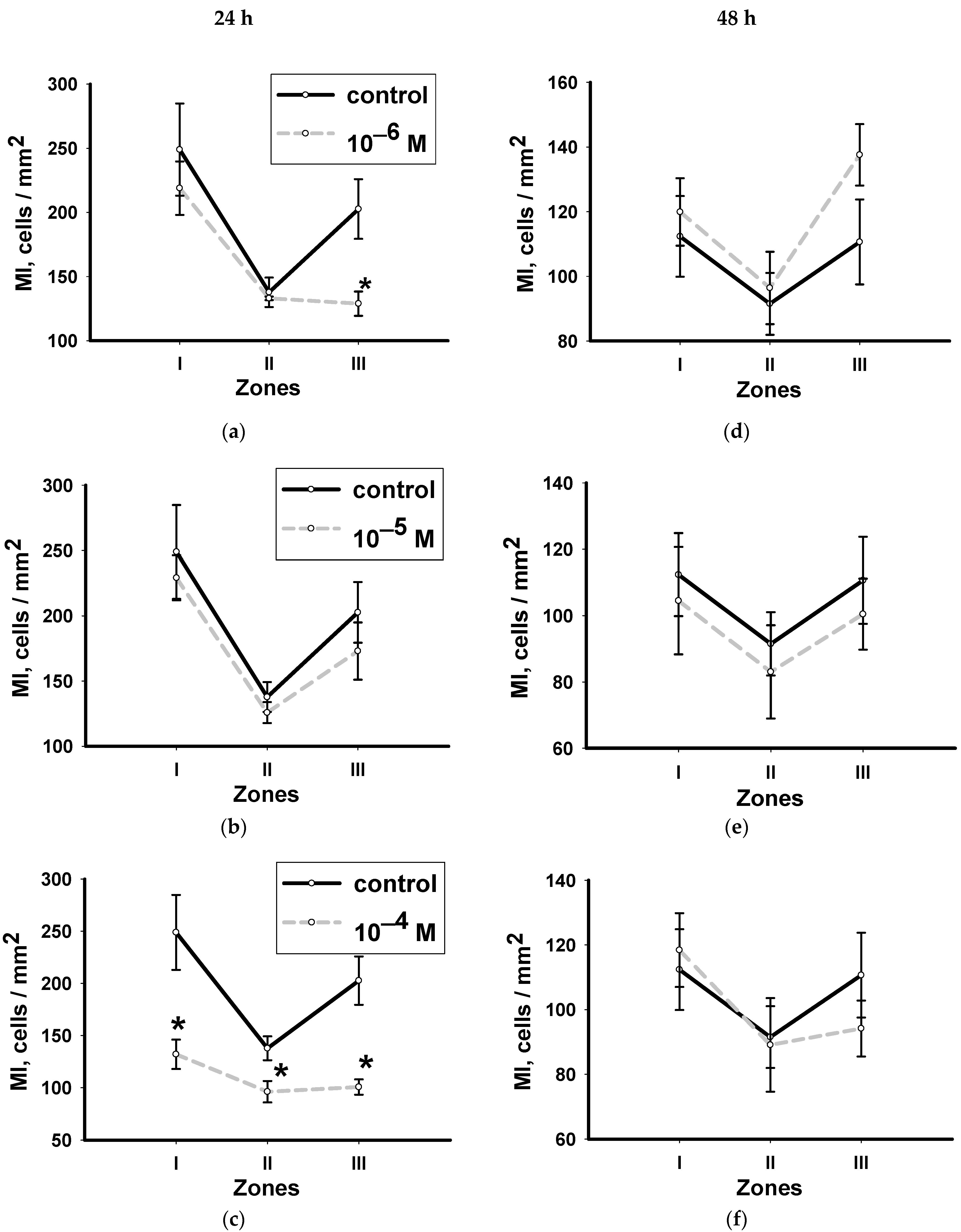
3.2. Dynamics of Changes in the Mitotic Index with Hydrocortisone in Various Zones of Head Blastema Regenerants
3.3. Dynamics of Changes in the Regeneration Coefficient of Head and Tail Blastema with Methylprednisolone
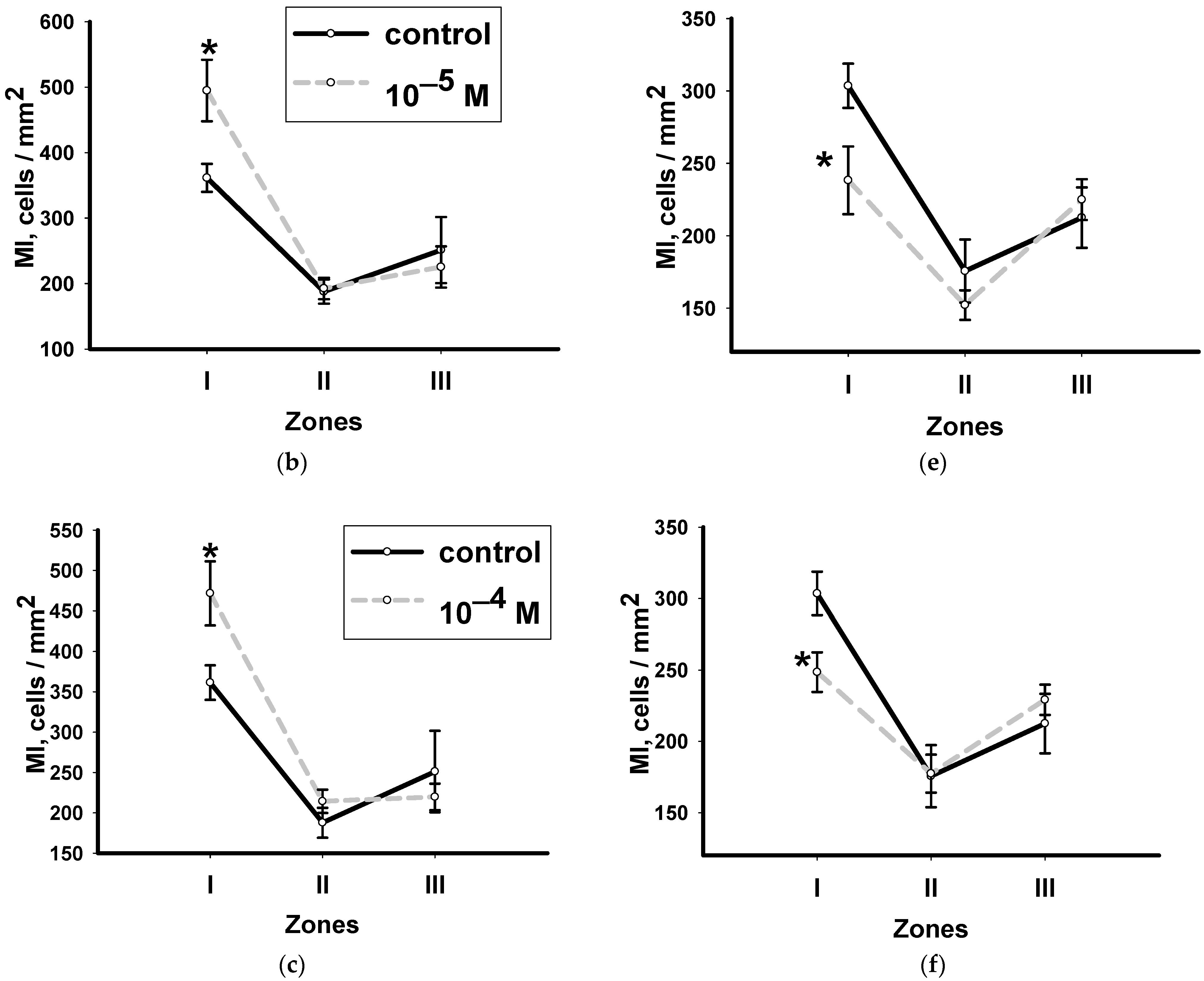
3.4. Dynamics of Changes in the Mitotic Index with Methylprednisolone in Various Zones of Head Blastema Regenerants
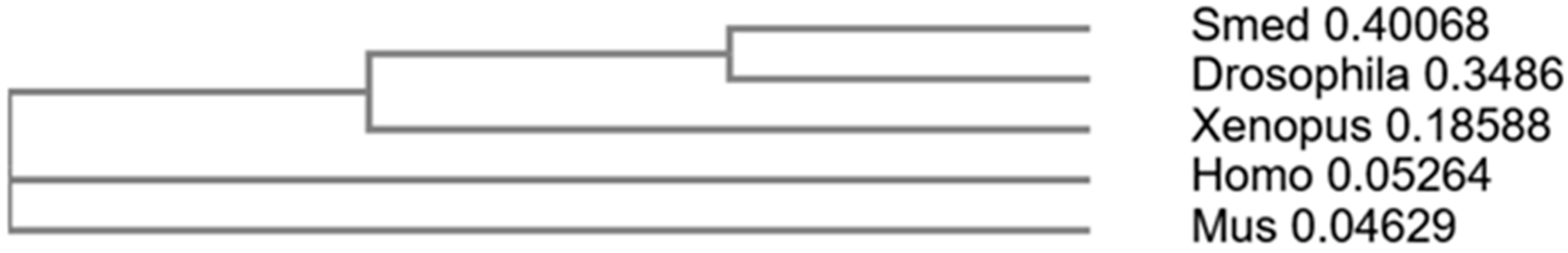
3.5. The Search for Homologs of Steroid Hormone Receptor Proteins
4. Discussion
5. Conclusions
- Hydrocortisone and its synthetic analogue methylprednisolone are able to influence the course of the morphogenetic process in the Girardia tigrina planarian. There is a dependence of the nature and exposure degree of hormones on their qualitative specificity, the concentration and the regenerating fragment; the head or tail area. The anterior regenerants have less sensitivity to the studied hormones.
- The nature of the hydrocortisone effect on the regeneration process differs from its synthetic analogue methylprednisolone which can act as an antagonist and an agonist of hydrocortisone. A high concentration of hydrocortisone suppresses the recovery of the amputated area, and low ones stimulate it. The nature of the methylprednisolone effect in the range of concentrations similar to the hydrocortisone ones, was more dependent on the specific concentration of the hormone and was mainly inhibitory.
- Both hormones change the mitotic activity of neoblasts during the first peak of proliferation. Their effects are the opposite: hydrocortisone stimulates the process in the periwound zone, and methylprednisolone, on the contrary, inhibits it mainly in the zone furthest from the wound. The second wave of mitotic activity does not depend on methylprednisolone, and hydrocortisone inhibits the proliferation of neoblasts.
- An orthologous protein of the glucocorticoid receptor was found in the SmedGD genomic database of the Schmidtea mediterranea planaria. Based on the obtained results, it can be proposed that flatworms have nuclear receptors for glucocorticoid steroid hormones. All of them have conservative domains that characterize the performed functions and probably play a role in many biological processes in planarians.
- It can be assumed as a working hypothesis that exogenous hormones from the glucocorticoid group affect the endogenous mechanisms of endocrine regeneration, including through hormone–receptor interaction. The revealed features of the effects of hydrocortisone and methylprednisolone may be associated with the formation of specific hormone–receptor complexes, as well as with the dependence of the methylprednisolone effects on the type of interaction; as an agonist or an antagonist.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, S.J.; Pearson, B.J. (Neo)blast from the past: New insights into planarian stem cell lineages. Curr. Opin. Genet. Dev. 2016, 40, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Sheiman, I.M. Morphological activity of regulatory neuropeptides and regeneration of planaria. Biomedicine 2008, 1, 79–87. [Google Scholar]
- Antebi, A. Nuclear receptor signal transduction in C. elegans. In WormBook: The Online Review of C. elegans Biology; WormBook: Eugene, OR, USA, 2015; pp. 1–49. [Google Scholar] [CrossRef]
- Labbé, R.M.; Irimia, M.; Currie, K.W.; Lin, A.; Zhu, S.J.; Brown, D.D.R.; Ross, E.J.; Voisin, V.; Bader, G.D.; Blencowe, B.J.; et al. A comparative transcriptomic analysis reveals conserved features of stem cell pluripotency in planarians and mammals. Stem Cells 2012, 30, 1734–1745. [Google Scholar] [CrossRef] [PubMed]
- Maule, A.G.; Shaw, C.; Halton, D.W.; Johnston, C.F.; Fairweather, I. Localization, quantification, and characterization of pancreatic polypeptide immunoreactivity in the parasitic flatworm Diclidophora merlangi and its fish host (Merlangius merlangus). Gen. Comp. Endocrinol. 1989, 74, 50–56. [Google Scholar] [CrossRef]
- Kreshchenko, N.D.; Terenina, N.B.; Kuchin, A.V. Some details of muscles innervations by FMRF-like nerve elements in planarian Girardia tigrina. Zoomorphology 2018, 137, 231–240. [Google Scholar] [CrossRef]
- Terenina, N.B.; Kreshchenko, N.D.; Mochalova, N.B.; Movsesyan, S.O. Serotonin and Neuropeptide FMRFamide in the Attachment Organs of Trematodes. Helminthologia 2018, 55, 185–194. [Google Scholar] [CrossRef]
- Kreshchenko, N.; Terenina, N.; Ermakov, A. Serotonin Signalling in Flatworms: An Immunocytochemical Localisation of 5-HT7 Type of Serotonin Receptors in Opisthorchis felineus and Hymenolepis diminuta. Biomolecules 2021, 11, 1212. [Google Scholar] [CrossRef]
- Kudikina, N.P. Steroid Hormones in the Life Cycle of Bivalves; Publishing House of Immanuel Kant Russian State University: Kaliningrad, Russia, 2008; pp. 79–88. [Google Scholar]
- Kreshchenko, N.; Sheiman, I.M. Pharynx regeneration in planaria. The effect of neuropeptides. Russ. J. Dev. Biol. 1994, 25, 350–356. [Google Scholar]
- Kudikina, N.P. Organization of Endocrine Function in Flatworms; Publishing House of Immanuel Kant Russian State University: Kaliningrad, Russia, 2011; pp. 126–131. [Google Scholar]
- Brindley, P.J.; da Costa, J.M.C.; Sripa, B. Why Does Infection with Some Helminths Cause Cancer? Trends Cancer 2015, 1, 174–182. [Google Scholar] [CrossRef]
- Vale, N.; Gouveia, M.J.; Botelho, M.; Sripa, B.; Suttiprapa, S.; Rinaldi, G.; Gomes, P.; Brindley, P.J.; Correia da Costa, J.M. Carcinogenic liver fluke Opisthorchis viverrini oxysterols detected by LC-MS/MS survey of soluble fraction parasite extract. Parasitol. Int. 2013, 62, 535–542. [Google Scholar] [CrossRef]
- Nikitina, S. Biologically Active Steroid Compounds of Invertebrates; Publishing House of Immanuel Kant Russian State University: Kaliningrad, Russia, 2019. [Google Scholar]
- Kudikina, N.P. Influence of Exogenous Hormonal Compounds on the Regeneration Process in Flatworms; Publishing House of Immanuel Kant Russian State University: Kaliningrad, Russia, 2012; pp. 84–91. [Google Scholar]
- Kudikina, N.P.; Ermakov, A.M.; Omelnitskaya, E.A.; Skorobogatykh, I.A. The Morphogenetic Effects of Exogenous Sex Steroid Hormones in the Planarian Girardia tigrina (Turbellaria, Tricladida). Biophysics 2019, 64, 765–771. [Google Scholar] [CrossRef]
- Auphan, N.; DiDonato, J.A.; Rosette, C.; Helmberg, A.; Karin, M. Immunosuppression by glucocorticoids: Inhibition of NF-kappa B activity through induction of I kappa B synthesis. Science 1995, 270, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Paragliola, R.M.; Papi, G.; Pontecorvi, A.; Corsello, S.M. Treatment with Synthetic Glucocorticoids and the Hypothalamus-Pituitary-Adrenal Axis. Int. J. Mol. Sci. 2017, 18, 2201. [Google Scholar] [CrossRef] [PubMed]
- Ermakova, O.N.; Ermakov, A.M.; Tiras Kh, P.; Lednev, V.V. Melatonin effect on the regeneration of the flatworm Girardia tigrina. Ontogenez 2009, 40, 466–469. [Google Scholar] [CrossRef]
- Newmark, P.A.; Sánchez Alvarado, A. Bromodeoxyuridine specifically labels the regenerative stem cells of planarians. Dev. Biol. 2000, 220, 142–153. [Google Scholar] [CrossRef]
- Turner, H.E.; Eastell, R.; Grossman, A. Oxford Desk Reference: Endocrinology; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Abdanipour, A.; Sagha, M.; Noori-Zadeh, A.; Pakzad, I.; Tiraihi, T. In vitro study of the long-term cortisol treatment effects on the growth rate and proliferation of the neural stem/precursor cells. Neurol. Res. 2015, 37, 117–124. [Google Scholar] [CrossRef]
- Newnham, J.P.; Moss, T.J. Antenatal glucocorticoids and growth: Single versus multiple doses in animal and human studies. Semin. Neonatol. 2001, 6, 285–292. [Google Scholar] [CrossRef]
- Rosan, V.B. Fundamentals of Endocrinology; Publishing House of Moscow State University: Moscow, Russia, 2010. [Google Scholar]
- Gujral, J.; Yau, M.; Yang, A.C.; Kastury, R.; Romero, C.J.; Wallach, E.; Wilkes, M.; Costin, G.; Rapaport, R. Primary Cortisol Deficiency and Growth Hormone Deficiency in a Neonate with Hypoglycemia: Coincidence or Consequence? J. Endocr. Soc. 2019, 3, 838–846. [Google Scholar] [CrossRef]
- Dong, J.; Li, J.; Li, J.; Cui, L.; Meng, X.; Qu, Y.; Wang, H. The proliferative effect of cortisol on bovine endometrial epithelial cells. Reprod. Biol. Endocrinol. 2019, 17, 97. [Google Scholar] [CrossRef]
- Takanosawa, M.; Nishino, H.; Ohta, Y.; Ichimura, K. Glucocorticoids enhance regeneration of murine olfactory epithelium. Acta Oto-Laryngol. 2009, 129, 1002–1009. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, L.; Duff, G.W. A negative regulatory region containing a glucocorticosteroid response element (nGRE) in the human interleukin-1beta gene. DNA Cell Biol. 1997, 16, 145–152. [Google Scholar] [CrossRef]
- Li, X.G.; Lin, X.J.; Du, J.H.; Xu, S.Z.; Lou, X.F.; Chen, Z. Combination of methylprednisolone and rosiglitazone promotes recovery of neurological function after spinal cord injury. Neural Regen. Res. 2016, 11, 1678–1684. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, Y.; He, L.; Pang, M.; Luo, C.; Liu, B.; Rong, L. High-dose methylprednisolone for acute traumatic spinal cord injury: A meta-analysis. Neurology 2019, 93, e841–e850. [Google Scholar] [CrossRef] [PubMed]
- Lander, R.; Petersen, C.P. Wnt, Ptk7, and FGFRL expression gradients control trunk positional identity in planarian regeneration. eLife 2016, 5, e12850. [Google Scholar] [CrossRef] [PubMed]
- Escobedo, G.; Larralde, C.; Chavarria, A.; Cerbón, M.A.; Morales-Montor, J. Molecular mechanisms involved in the differential effects of sex steroids on the reproduction and infectivity of Taenia crassiceps. J. Parasitol. 2004, 90, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Li, D.J.; McMann, C.L.; Reddien, P.W. Nuclear receptor NR4A is required for patterning at the ends of the planarian anterior-posterior axis. eLife 2019, 8, e42015. [Google Scholar] [CrossRef]
- Musvasva, E.; Williams, K.A.; Muller, W.J.; Villet, M.H. Preliminary observations on the effects of hydrocortisone and sodium methohexital on development of Sarcophaga (Curranea) tibialis Macquart (Diptera: Sarcophagidae), and implications for estimating post mortem interval. Forensic Sci. Int. 2001, 120, 37–41. [Google Scholar] [CrossRef]
- Patricio-Gómez, J.M.; Valdez, R.A.; Veloz, A.; Aguilar-Vega, L.; Zurabian, R.; Romano, M.C. The synthesis of steroids by Taenia crassiceps WFU cysticerci and tapeworms is related to the developmental stages of the parasites. Gen. Comp. Endocrinol. 2018, 259, 154–160. [Google Scholar] [CrossRef]
- Aceves-Ramos, A.; de la Torre, P.; Hinojosa, L.; Ponce, A.; García-Villegas, R.; Laclette, J.P.; Bobes, R.J.; Romano, M.C. Cloning, characterization and functional expression of Taenia solium 17 beta-hydroxysteroid dehydrogenase. Gen. Comp. Endocrinol. 2014, 203, 186–192. [Google Scholar] [CrossRef]
- Romano, M.; Jiménez, P.; Miranda, C.; Valdez, R. Parasites and steroid hormones: Corticosteroid and sex steroid synthesis, their role in the parasite physiology and development. Front. Neurosci. 2015, 9, 224. [Google Scholar] [CrossRef]
- Ziniel, P.D.; Karumudi, B.; Barnard, A.H.; Fisher, E.M.; Thatcher, G.R.; Podust, L.M.; Williams, D.L. The Schistosoma mansoni Cytochrome P450 (CYP3050A1) Is Essential for Worm Survival and Egg Development. PLoS Negl. Trop. Dis. 2015, 9, e0004279. [Google Scholar] [CrossRef] [PubMed]
- Romano, M.C.; Valdéz, R.A.; Cartas, A.L.; Gómez, Y.; Larralde, C. Steroid hormone production by parasites: The case of Taenia crassiceps and Taenia solium cysticerci. J. Steroid Biochem. Mol. Biol. 2003, 85, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Tewari, A.G.; Stern, S.R.; Oderberg, I.M.; Reddien, P.W. Cellular and Molecular Responses Unique to Major Injury Are Dispensable for Planarian Regeneration. Cell Rep. 2018, 25, 2577–2590.e73. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ermakov, A.; Kudykina, N.; Bykova, A.; Tkacheva, U. Morphogenic Effect of Exogenous Glucocorticoid Hormones in the Girardia tigrina Planarian (Turbellaria, Tricladida). Biology 2023, 12, 292. https://doi.org/10.3390/biology12020292
Ermakov A, Kudykina N, Bykova A, Tkacheva U. Morphogenic Effect of Exogenous Glucocorticoid Hormones in the Girardia tigrina Planarian (Turbellaria, Tricladida). Biology. 2023; 12(2):292. https://doi.org/10.3390/biology12020292
Chicago/Turabian StyleErmakov, Artem, Natalia Kudykina, Arina Bykova, and Ulyana Tkacheva. 2023. "Morphogenic Effect of Exogenous Glucocorticoid Hormones in the Girardia tigrina Planarian (Turbellaria, Tricladida)" Biology 12, no. 2: 292. https://doi.org/10.3390/biology12020292
APA StyleErmakov, A., Kudykina, N., Bykova, A., & Tkacheva, U. (2023). Morphogenic Effect of Exogenous Glucocorticoid Hormones in the Girardia tigrina Planarian (Turbellaria, Tricladida). Biology, 12(2), 292. https://doi.org/10.3390/biology12020292