


Transcriptome Profile Reveals Differences between Remote and Ischemic Myocardium after Acute Myocardial Infarction in a Swine Model
 , , , , , ,
, , , , , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. AMI Model Creation and Tissue Collection
2.2. Microarray Expression Analysis
2.3. Proteomic Analysis
2.4. Enrichment Analysis (Funtional Annotation)
3. Results
3.1. Transcriptomic Analysis from Remote and Infarcted Myocardium Tissue
3.2. Proteomic Validation of Differentially Expressed Transcripts
3.3. Enrichment Analysis of Validated Genes
3.4. Immune System Process in Infarcted Myocardium
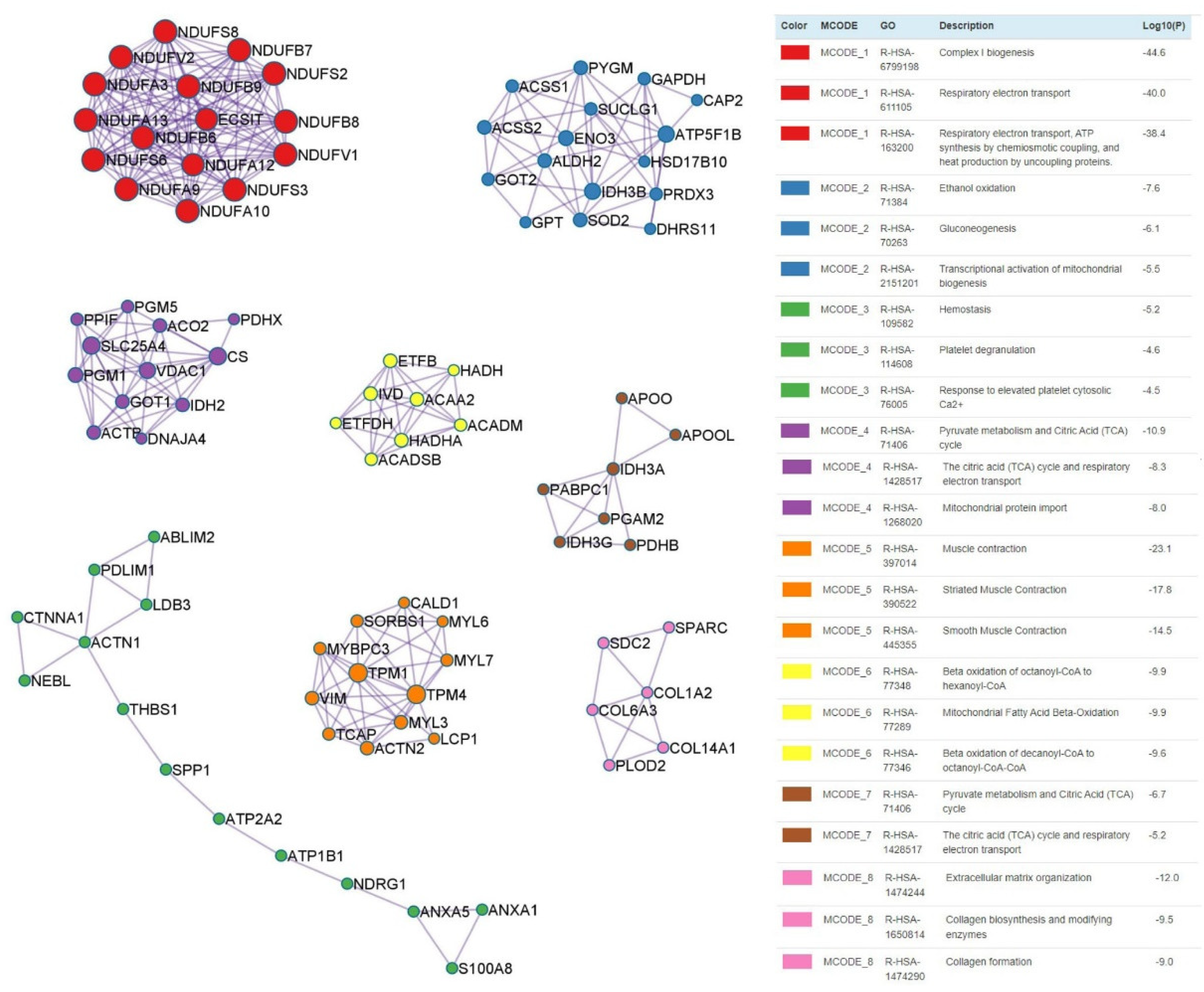
3.5. Protein–Protein Interaction Enrichment between Infarcted and Remote Myocardium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sutton, M.G.S.J.; Sharpe, N. Left Ventricular Remodeling After Myocardial Infarction: Pathophysiology and Therapy. Circulation 2000, 101, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Zuurbier, C.J.; Bertrand, L.; Beauloye, C.R.; Andreadou, I.; Ruiz-Meana, M.; Jespersen, N.R.; Kula-Alwar, D.; Prag, H.A.; Eric Botker, H.; Dambrova, M.; et al. Cardiac Metabolism as a Driver and Therapeutic Target of Myocardial Infarction. J. Cell. Mol. Med. 2020, 24, 5937–5954. [Google Scholar] [CrossRef] [PubMed]
- Lehnart, S.E.; Maier, L.S.; Hasenfuss, G. Abnormalities of Calcium Metabolism and Myocardial Contractility Depression in the Failing Heart. Heart Fail. Rev. 2009, 14, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frangogiannis, N. The Inflammatory Response in Myocardial Infarction. Cardiovasc. Res. 2002, 53, 31–47. [Google Scholar] [CrossRef]
- Shahzad, S.; Mateen, S.; Hasan, A.; Moin, S. GRACE Score of Myocardial Infarction Patients Correlates with Oxidative Stress Index, HsCRP and Inflammation. Immunobiology 2019, 224, 433–439. [Google Scholar] [CrossRef]
- Schirone, L.; Forte, M.; Palmerio, S.; Yee, D.; Nocella, C.; Angelini, F.; Pagano, F.; Schiavon, S.; Bordin, A.; Carrizzo, A.; et al. A Review of the Molecular Mechanisms Underlying the Development and Progression of Cardiac Remodeling. Oxidative Med. Cell. Longev. 2017, 2017, 3920195. [Google Scholar] [CrossRef]
- Kuppe, C.; Ramirez Flores, R.O.; Li, Z.; Hayat, S.; Levinson, R.T.; Liao, X.; Hannani, M.T.; Tanevski, J.; Wünnemann, F.; Nagai, J.S.; et al. Spatial Multi-Omic Map of Human Myocardial Infarction. Nature 2022, 608, 766–777. [Google Scholar] [CrossRef]
- Yamada, S.; Ko, T.; Hatsuse, S.; Nomura, S.; Zhang, B.; Dai, Z.; Inoue, S.; Kubota, M.; Sawami, K.; Yamada, T.; et al. Spatiotemporal Transcriptome Analysis Reveals Critical Roles for Mechano-Sensing Genes at the Border Zone in Remodeling after Myocardial Infarction. Nat. Cardiovasc. Res. 2022, 1, 1072–1083. [Google Scholar] [CrossRef]
- Warr, A.; Affara, N.; Aken, B.; Beiki, H.; Bickhart, D.M.; Billis, K.; Chow, W.; Eory, L.; Finlayson, H.A.; Flicek, P.; et al. An Improved Pig Reference Genome Sequence to Enable Pig Genetics and Genomics Research. GigaScience 2020, 9, giaa051. [Google Scholar] [CrossRef]
- Pan, Z.; Yao, Y.; Yin, H.; Cai, Z.; Wang, Y.; Bai, L.; Kern, C.; Halstead, M.; Chanthavixay, G.; Trakooljul, N.; et al. Pig Genome Functional Annotation Enhances the Biological Interpretation of Complex Traits and Human Disease. Nat Commun. 2021, 12, 5848. [Google Scholar] [CrossRef]
- Karlsson, M.; Sjöstedt, E.; Oksvold, P.; Sivertsson, Å.; Huang, J.; Álvez, M.B.; Arif, M.; Li, X.; Lin, L.; Yu, J.; et al. Genome-Wide Annotation of Protein-Coding Genes in Pig. BMC Biol. 2022, 20, 25. [Google Scholar] [CrossRef] [PubMed]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE Guidelines 2.0: Updated Guidelines for Reporting Animal Research. BMC Vet. Res. 2020, 16, 242. [Google Scholar] [CrossRef] [PubMed]
- Crisostomo, V.; Baez-Diaz, C.; Maestre, J.; Garcia-Lindo, M.; Sun, F.; Casado, J.G.; Blazquez, R.; Abad, J.L.; Palacios, I.; Rodriguez-Borlado, L.; et al. Delayed Administration of Allogeneic Cardiac Stem Cell Therapy for Acute Myocardial Infarction Could Ameliorate Adverse Remodeling: Experimental Study in Swine. J Transl. Med. 2015, 13, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, E.; Marinaro, F.; de Pedro, M.d.l.Á.; Sánchez-Margallo, F.M.; Gómez-Serrano, M.; Ponath, V.; Pogge von Strandmann, E.; Jorge, I.; Vázquez, J.; Fernández-Pereira, L.M.; et al. The Immunomodulatory Signature of Extracellular Vesicles From Cardiosphere-Derived Cells: A Proteomic and MiRNA Profiling. Front. Cell Dev. Biol. 2020, 8, 321. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Consortium Expansion of the Gene Ontology Knowledgebase and Resources. Nucleic Acids Res. 2017, 45, D331–D338. [CrossRef] [Green Version]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2019, 48(D1), D498–D503. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. Pathophysiology of Myocardial Infarction. In Comprehensive Physiology; Terjung, R., Ed.; Wiley: Hoboken, NJ, USA, 2015; pp. 1841–1875. ISBN 978-0-470-65071-4. [Google Scholar]
- Maier, T.; Güell, M.; Serrano, L. Correlation of MRNA and Protein in Complex Biological Samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef] [Green Version]
- Popovic, D.; Koch, B.; Kueblbeck, M.; Ellenberg, J.; Pelkmans, L. Multivariate Control of Transcript to Protein Variability in Single Mammalian Cells. Cell Syst. 2018, 7, 398–411.e6. [Google Scholar] [CrossRef] [Green Version]
- Chandra Shekar, K.; Yannopoulos, D.; Kosmopoulos, M.; Riess, M.L. Differential Effects of Reperfusion on Cardiac Mitochondrial Subpopulations in a Preclinical Porcine Model of Acute Myocardial Infarction. Front. Cell Dev. Biol. 2022, 10, 843733. [Google Scholar] [CrossRef] [PubMed]
- Bugger, H.; Pfeil, K. Mitochondrial ROS in Myocardial Ischemia Reperfusion and Remodeling. Biochim. Biophys. Acta—Mol. Basis Dis. 2020, 1866, 165768. [Google Scholar] [CrossRef] [PubMed]
- Saparov, A.; Ogay, V.; Nurgozhin, T.; Chen, W.C.W.; Mansurov, N.; Issabekova, A.; Zhakupova, J. Role of the Immune System in Cardiac Tissue Damage and Repair Following Myocardial Infarction. Inflamm. Res. 2017, 66, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Nahrendorf, M.; Swirski, F.K. Monocyte and Macrophage Heterogeneity in the Heart. Circ. Res. 2013, 112, 1624–1633. [Google Scholar] [CrossRef]
- Lee, W.W.; Marinelli, B.; van der Laan, A.M.; Sena, B.F.; Gorbatov, R.; Leuschner, F.; Dutta, P.; Iwamoto, Y.; Ueno, T.; Begieneman, M.P.V.; et al. PET/MRI of Inflammation in Myocardial Infarction. J. Am. Coll. Cardiol. 2012, 59, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Shi, K.; Xu, H.; He, Y.; Ma, M.; Zhang, L.; Wang, J.; Li, X.; Fu, C.; Li, H.; et al. Inflammation in Remote Myocardium and Left Ventricular Remodeling After Acute Myocardial Infarction: A Pilot Study Using T2 Mapping. J. Magn. Reson. Imaging 2022, 55, 555–564. [Google Scholar] [CrossRef]
- Ruparelia, N.; Digby, J.E.; Jefferson, A.; Medway, D.J.; Neubauer, S.; Lygate, C.A.; Choudhury, R.P. Myocardial Infarction Causes Inflammation and Leukocyte Recruitment at Remote Sites in the Myocardium and in the Renal Glomerulus. Inflamm. Res. 2013, 62, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Paradies, G.; Petrosillo, G.; Pistolese, M.; Di Venosa, N.; Federici, A.; Ruggiero, F.M. Decrease in Mitochondrial Complex I Activity in Ischemic/Reperfused Rat Heart: Involvement of Reactive Oxygen Species and Cardiolipin. Circ. Res. 2004, 94, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Ide, T.; Tsutsui, H.; Kinugawa, S.; Utsumi, H.; Kang, D.; Hattori, N.; Uchida, K.; Arimura, K.; Egashira, K.; Takeshita, A. Mitochondrial Electron Transport Complex I Is a Potential Source of Oxygen Free Radicals in the Failing Myocardium. Circ. Res. 1999, 85, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, B.; Yang, X.; Zhang, C.; Jiao, Y.; Li, P.; Liu, Y.; Li, Z.; Qiao, B.; Bond Lau, W.; et al. S100a8/A9 Signaling Causes Mitochondrial Dysfunction and Cardiomyocyte Death in Response to Ischemic/Reperfusion Injury. Circulation 2019, 140, 751–764. [Google Scholar] [CrossRef]
- Page, B.; Young, R.; Iyer, V.; Suzuki, G.; Lis, M.; Korotchkina, L.; Patel, M.S.; Blumenthal, K.M.; Fallavollita, J.A.; Canty, J.M. Persistent Regional Downregulation in Mitochondrial Enzymes and Upregulation of Stress Proteins in Swine With Chronic Hibernating Myocardium. Circ. Res. 2008, 102, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, J.; Young, R.; Page, B.J.; Shen, X.; Tata, N.; Li, J.; Duan, X.; Fallavollita, J.A.; Canty, J.M. Reproducible Ion-Current-Based Approach for 24-Plex Comparison of the Tissue Proteomes of Hibernating versus Normal Myocardium in Swine Models. J. Proteome Res. 2014, 13, 2571–2584. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Dobaczewski, M.; Gonzalez-Quesada, C.; Chen, W.; Biernacka, A.; Li, N.; Lee, D.-W.; Frangogiannis, N.G. Endogenous Thrombospondin 1 Protects the Pressure-Overloaded Myocardium by Modulating Fibroblast Phenotype and Matrix Metabolism. Hypertension 2011, 58, 902–911. [Google Scholar] [CrossRef] [Green Version]
- Chiasson, V.; Takano, A.P.C.; Guleria, R.S.; Gupta, S. Deficiency of MicroRNA MiR-1954 Promotes Cardiac Remodeling and Fibrosis. JAHA 2019, 8, e012880. [Google Scholar] [CrossRef] [PubMed]
- Hofstra, L.; Liem, I.H.; Dumont, E.A.; Boersma, H.H.; van Heerde, W.L.; Doevendans, P.A.; DeMuinck, E.; Wellens, H.; Kemerink, G.J.; Reutelingsperger, C.P.; et al. Visualisation of Cell Death in Vivo in Patients with Acute Myocardial Infarction. Lancet 2000, 356, 209–212. [Google Scholar] [CrossRef]
- Yoshimi, N.; Futamura, T.; Bergen, S.E.; Iwayama, Y.; Ishima, T.; Sellgren, C.; Ekman, C.J.; Jakobsson, J.; Pålsson, E.; Kakumoto, K.; et al. Cerebrospinal Fluid Metabolomics Identifies a Key Role of Isocitrate Dehydrogenase in Bipolar Disorder: Evidence in Support of Mitochondrial Dysfunction Hypothesis. Mol. Psychiatry 2016, 21, 1504–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watany, M.M.; Nagi Abd-Ellatif, R.; Ezzelregal Abdeldayem, M.; El-sayed El-Horany, H. Association between Genetic Variations of Mitochondrial Isocitrate Dehydrogenase (IDH2) and Acute Myocardial Infarction. Gene 2022, 829, 146497. [Google Scholar] [CrossRef]
- Subramani, J.; Kundumani-Sridharan, V.; Das, K.C. Thioredoxin Protects Mitochondrial Structure, Function and Biogenesis in Myocardial Ischemia-Reperfusion via Redox-Dependent Activation of AKT-CREB- PGC1α Pathway in Aged Mice. Aging 2020, 12, 19809–19827. [Google Scholar] [CrossRef]
- Tucholski, T.; Cai, W.; Gregorich, Z.R.; Bayne, E.F.; Mitchell, S.D.; McIlwain, S.J.; de Lange, W.J.; Wrobbel, M.; Karp, H.; Hite, Z.; et al. Distinct Hypertrophic Cardiomyopathy Genotypes Result in Convergent Sarcomeric Proteoform Profiles Revealed by Top-down Proteomics. Proc. Natl. Acad. Sci. USA 2020, 117, 24691–24700. [Google Scholar] [CrossRef]
- Dobaczewski, M.; Gonzalez-Quesada, C.; Frangogiannis, N.G. The Extracellular Matrix as a Modulator of the Inflammatory and Reparative Response Following Myocardial Infarction. J. Mol. Cell. Cardiol. 2010, 48, 504–511. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pulido, M.; de Pedro, M.Á.; Álvarez, V.; Marchena, A.M.; Blanco-Blázquez, V.; Báez-Díaz, C.; Crisóstomo, V.; Casado, J.G.; Sánchez-Margallo, F.M.; López, E. Transcriptome Profile Reveals Differences between Remote and Ischemic Myocardium after Acute Myocardial Infarction in a Swine Model. Biology 2023, 12, 340. https://doi.org/10.3390/biology12030340
Pulido M, de Pedro MÁ, Álvarez V, Marchena AM, Blanco-Blázquez V, Báez-Díaz C, Crisóstomo V, Casado JG, Sánchez-Margallo FM, López E. Transcriptome Profile Reveals Differences between Remote and Ischemic Myocardium after Acute Myocardial Infarction in a Swine Model. Biology. 2023; 12(3):340. https://doi.org/10.3390/biology12030340
Chicago/Turabian StylePulido, María, María Ángeles de Pedro, Verónica Álvarez, Ana María Marchena, Virginia Blanco-Blázquez, Claudia Báez-Díaz, Verónica Crisóstomo, Javier G. Casado, Francisco Miguel Sánchez-Margallo, and Esther López. 2023. "Transcriptome Profile Reveals Differences between Remote and Ischemic Myocardium after Acute Myocardial Infarction in a Swine Model" Biology 12, no. 3: 340. https://doi.org/10.3390/biology12030340
APA StylePulido, M., de Pedro, M. Á., Álvarez, V., Marchena, A. M., Blanco-Blázquez, V., Báez-Díaz, C., Crisóstomo, V., Casado, J. G., Sánchez-Margallo, F. M., & López, E. (2023). Transcriptome Profile Reveals Differences between Remote and Ischemic Myocardium after Acute Myocardial Infarction in a Swine Model. Biology, 12(3), 340. https://doi.org/10.3390/biology12030340