
European Pond Turtle (Emys orbicularis) Nest Predation: A Study with Artificial Nests

{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
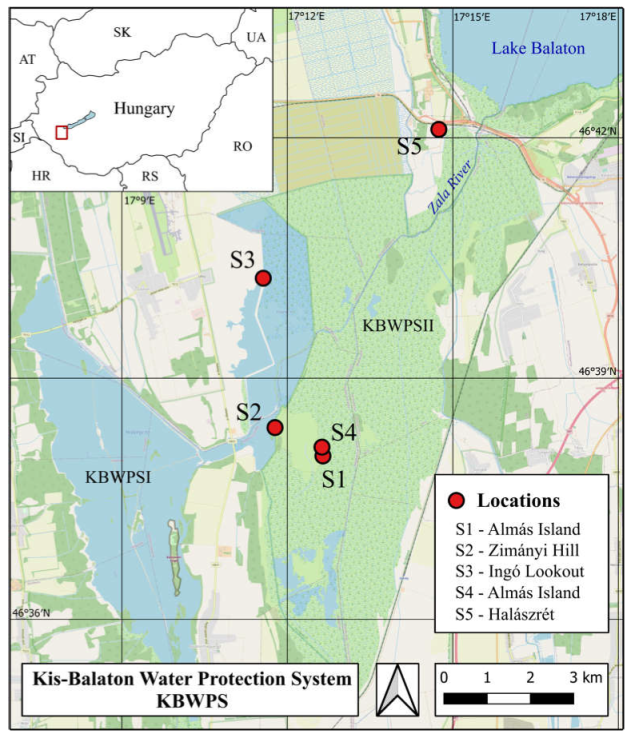
2.1. Study Area and Sampling Sites
2.2. European Pond Turtle and Its Predators
2.3. Experiment with Artificial Turtle Nests
2.4. Testing the Effect of Turtle Urine and Marker Stick
2.5. Data Analysis
3. Results
3.1. Predation on Previously Depredated Real and New Artificial Nests
3.2. The Predation Depends on Spatial Distribution of Nests
3.3. Identified Turtle Egg Predators
3.4. The Effect of Turtle Urine and Marker Stick on Predation Events
4. Discussion
4.1. High-Rate Nest Predation in the First Days after Egg Laying
4.2. Predation Rates in Previously Depredated Real and New Artificial Nests Was Similar
4.3. Spatial Distribution of Nests Affected Predation Rates
4.4. Red Fox Is Identified as Dominant Predator of Turtle Eggs
4.5. Cues Moderately Affected Predators to Locate Nests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fritz, U. Die Europäische Sumpfschildkröte (Emys orbicularis); Laurenti Verlag: Bielefeld, Germany, 2003. [Google Scholar]
- Fritz, U.; Chiari, Y. Conservation actions for European pond turtles—A summary of current efforts in distinct European countries. Herpetol. Notes 2013, 6, 105. [Google Scholar]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Philos. Trans. R. Soc. B 2010, 365, 2093–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markovic, D.; Carrizo, S.; Freyhof, J.; Cid, N.; Lengyel, S.; Scholz, M.; Kasperdius, H.; Darwall, W. Europe’s freshwater biodiversity under climate change: Distribution shifts and conservation needs. Divers. Distrib. 2014, 20, 1097–1107. [Google Scholar] [CrossRef]
- Albert, J.S.; Destouni, G.; Duke-Sylvester, S.M.; Magurran, A.E.; Oberdorff, T.; Reis, R.E.; Winemiller, K.O.; Ripple, W.J. Scientists’ warning to humanity on the freshwater biodiversity crisis. Ambio 2021, 50, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Németh, G.; Lóczy, D.; Gyenizse, P. Long-term land use and landscape pattern changes in a marshland of Hungary. Sustainability 2021, 13, 12664. [Google Scholar] [CrossRef]
- Ramsar Sites Information Service. Kis-Balaton. Available online: https://rsis.ramsar.org/ris/185 (accessed on 20 April 2022).
- Battin, J. When good animals love bad habitats: Ecological traps and the conservation of animal populations. Conserv. Biol. 2004, 18, 1482–1491. [Google Scholar] [CrossRef]
- Hale, R.; Swearer, S.E. Ecological traps: Current evidence and future directions. Proc. Royal Soc. B 2016, 283, 20152647. [Google Scholar] [CrossRef] [Green Version]
- Magyari, M.; (Balaton Uplands National Park Directorate, Veszprém, Hungary). Personal communication, 2022.
- Schindler, M.; Frötscher, H.; Hille, A.; Bruck, M.R.; Schmidt, M.; Kornilev, Y.V. Nest protection during a long-term conservation project as a tool to increase the autochthonous population of Emys orbicularis (L., 1758) in Austria. Acta Zool. Bulg. 2017, 69 (Suppl. S10), 147–154. [Google Scholar]
- Hamilton, A.M.; Freedman, A.H.; Franz, R. Effects of deer feeders, habitat and sensory cues on predation rates on artificial turtle nests. Am. Midl. Nat. 2002, 147, 123–134. [Google Scholar] [CrossRef]
- Marchand, M.N.; Litvaitis, J.A.; Maier, T.J.; DeGraaf, R.M. Use of artificial nests to investigate predation on freshwater turtle nests. Wildl. Soc. Bull. 2002, 30, 1092–1098. Available online: https://www.fs.usda.gov/treesearch/pubs/14346 (accessed on 30 April 2020).
- Marchand, M.N.; Litvaitis, J.A. Effects of landscape composition, habitat features, and nest distribution on predation rates of simulated turtle nests. Biol. Conserv. 2004, 117, 243–251. [Google Scholar] [CrossRef]
- Horváth, E.; Kaňuch, P.; Uhrin, M. Predation on nests of the European pond turtle (Emys orbicularis): Remarks from failed field experiments. Herpetol. Notes 2021, 14, 1067–1072. [Google Scholar]
- Cserny, T.; Nagy-Bodor, E. Limnogeological investigations on Lake Balaton (Hungary). In Lake Basins through Space and Time; Gierlowski-Kordesch, E.H., Kelts, K.R., Eds.; Studies in Geology 46; American Association of Petroleum Geologists: Tulsa, OK, USA, 2000; pp. 605–617. [Google Scholar] [CrossRef]
- Tátrai, I.; Kálmán, M.; Korponai, J.; Paulovits, G.; Pomogyi, P. The role of the Kis-Balaton Water Protection System in the control of water quality of Lake Balaton. Ecol. Eng. 2000, 16, 73–78. [Google Scholar] [CrossRef]
- NATURA 2000 Standard Data Form—Kis-Balaton (HUBF30003). Available online: https://natura2000.eea.europa.eu/Natura2000/SDF.aspx?site=HUBF30003 (accessed on 20 April 2022).
- Árva, D.; Tóth, M.; Mozsár, A.; Specziár, A. The roles of environment, site position, and seasonality in taxonomic and functional organization of chironomid assemblages in a heterogeneous wetland, Kis-Balaton (Hungary). Hydrobiologia 2017, 787, 353–373. [Google Scholar] [CrossRef] [Green Version]
- Lanszki, Z.; Horváth, G.F.; Bende, Z.; Lanszki, J. Differences in the diet and trophic niche of three sympatric carnivores in a marshland. Mammal Res. 2020, 65, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Mandl, É.; (Hungarian Meteorological Service, Budapest, Hungary). Personal communication, 2022.
- Farkas, B. Noteworthy facts about the European pond turtle. In Past, Present, Future of the European Pond Turtle; Kovács, Z., Ed.; Budapest Zoo: Budapest, Hungary, 2008; pp. 8–67. [Google Scholar]
- Zuffi, M.A.L.; Odetti, F. Double egg-deposition in the European pond turtle, Emys orbicularis, from central Italy. Ital. J. Zool. 1998, 65, 187–189. [Google Scholar] [CrossRef] [Green Version]
- Kiss, I.; Erdélyi, G.; Szabó, B. Nesting activity and reproductive success of Emys orbicularis in Babat Valley (Gödöllő, Hungary). Herpetol. Conserv. Biol. 2021, 16, 624–638. [Google Scholar]
- Zuffi, M.A. Conservation biology of the European pond turtle, Emys orbicularis, in Italy: Review of systematics and reproductive ecology patterns (Reptilia, Emydidae). Ital. J. Zool. 2004, 71, 103–105. [Google Scholar] [CrossRef] [Green Version]
- Novotný, M.; Danko, S.; Havas, P. Activity cycle and reproductive characteristics of the European pond turtle (Emys orbicularis) in the Tajba National Nature Reserve, Slovakia. Biologia 2004, 59 (Suppl. S14), 113–121. [Google Scholar]
- Rogner, M. Nesting. In European Pond Turtle Emys Orbicularis; Rogner, M., Ed.; Chimaira: Frankfurt am Main, Germany, 2009; pp. 95–99. [Google Scholar]
- Meeske, M. Nesting ecology of European pond turtle (Emys orbicularis) in South Lithuania. Acta Zool. Litu. 1997, 7, 143–150. [Google Scholar] [CrossRef]
- Mitrus, S.; Zemanek, M. Distribution and biology of Emys orbicularis (L) in Poland. Stapfia 2000, 69, 107–118. [Google Scholar]
- Marián, M.; Szabó, I. Contribution to the biology of propagation of the tortoise Emys orbicularis L. Állat. Közlem. 1961, 48, 85–90. [Google Scholar]
- Purger, J.J.; Kurucz, K.; Tóth, Á.; Batáry, P. Coating plasticine eggs can eliminate the overestimation of predation on artificial ground nests. Bird Study 2012, 59, 350–352. [Google Scholar] [CrossRef]
- Major, R.E.; Kendal, C.E. The contribution of artificial nest experiments to understanding avian reproductive success: A review of methods and conclusions. Ibis 1996, 138, 298–307. [Google Scholar] [CrossRef]
- Bateman, P.W.; Fleming, P.A.; Wolfe, A.K. A different kind of ecological modelling: The use of clay model organisms to explore predator-prey interactions in vertebrates. J. Zool. 2017, 301, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Loman, J.; Göransson, G. Egg shell dumps and crow Corvus cornix predation on simulated birds’ nests. Oikos 1978, 30, 461–466. [Google Scholar] [CrossRef] [Green Version]
- Byer, N.W.; Reid, B.N.; Seigel, R.A.; Peery, M.Z. Applying lessons from avian ecology to herpetological research: Techniques for analyzing nest survival due to predation. Herpetol. Conserv. Biol. 2018, 13, 517–532. [Google Scholar]
- Wilhoft, D.C.; Del Baglivo, M.G.; Del Baglivo, M.D. Observations on mammalian prediation of snapping turtle nests (Reptilia, Testudines, Chelydridae). J. Herpetol. 1979, 13, 435–438. [Google Scholar] [CrossRef]
- Tuberville, T.D.; Burke, V.J. Do flag markers attract turtle nest predators? J. Herpetol. 1994, 28, 514–516. [Google Scholar] [CrossRef]
- Purger, J.J.; Mészáros, L.A. Possible effects of nest predation on the breeding success of Ferruginous Ducks Aythya nyroca. Bird Conserv. Int. 2006, 16, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Pärt, T.; Wretenberg, J. Do artificial nests reveal relative nest predation risk on real nests? J. Avian Biol. 2002, 33, 39–46. [Google Scholar] [CrossRef]
- Mayfield, H.F. Suggestions for calculating nest success. Wilson Bull. 1975, 87, 456–466. Available online: http://www.jstor.org/stable/4160682 (accessed on 16 March 2015).
- Johnson, D.H. Estimating nest success: The Mayfield method and an alternative. Auk 1979, 96, 651–661. [Google Scholar] [CrossRef]
- Halupka, K. J-Test. Available online: http://zeb.uni.wroc.pl/halupka/ (accessed on 20 February 2020).
- Farkas, B.; (Bercsényi St. 21, Gyúró, Hungary); Harmos, K.; (Bükk National Park Directorate, Eger, Hungary); Halpern, B.; (ELKH-ELTE-MTM Integrative Ecology Research Group, Budapest, Hungary). Mocsári teknős fészek-predáció monitorozása és lehetséges védelmi intézkedések tesztelése a Középső-Ipoly-völgyben. Project: Fenntartható természetvédelem magyarországi Natura 2000 területeken, SH/4/8. Scientific report. Unpublished work. 2014; 1–27. [Google Scholar]
- Najbar, B.; Szuszkiewicz, E. Reproductive ecology of the European pond turtle (Emys orbicularis Linnaeus, 1758) (Testudines: Emydidae) in western Poland. Acta Zool. Cracov. 2005, 48, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Riley, J.L.; Litzgus, J.D. Cues used by predators to detect freshwater turtle nests may persist late into incubation. Can. Field-Nat. 2014, 128, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Geller, G.A.; Parker, S.L. What are the primary cues used by mammalian predators to locate freshwater turtle nests? A critical review of the evidence. Front. Ecol. Evol. 2022, 9, 784786. [Google Scholar] [CrossRef]
- Congdon, J.D.; Breitenbach, G.L.; van Loben Sels, R.C.; Tinkle, D.W. Reproduction and nesting ecology of snapping turtles (Chelydra serpentina) in southeastern Michigan. Herpetologica 1987, 43, 39–54. Available online: https://www.jstor.org/stable/3892436 (accessed on 3 February 2018).
- Eckrich, C.E.; Owens, D.W. Solitary versus arribada nesting in the Olive Ridley sea Turtles (Lepidochelys olivacea): A test of the predator-satiation hypothesis. Herpetologica 1995, 51, 349–354. Available online: https://www.jstor.org/stable/3893041 (accessed on 26 September 2022).
- Spencer, R.J.; Thompson, M.B. Experimental analysis of the impact of foxes on freshwater turtle populations. Conserv. Biol. 2005, 19, 845–854. [Google Scholar] [CrossRef]
- Strickland, J.; Colbert, P.; Janzen, F.J. Experimental analysis of effects of markers and habitat structure on predation of turtle nests. J. Herpetol. 2010, 44, 467–470. [Google Scholar] [CrossRef]
- Bernstein, N.P.; McCollum, A.; Black, R.W. How do predators locate nests of ornate box turtles (Terrapene ornata)? A field experiment. Herpetol. Conserv. Biol. 2015, 10, 44–53. [Google Scholar]
- Geller, G.A. A test of substrate sweeping as a strategy to reduce raccoon predation of freshwater turtle nests, with insights from supplemental artificial nests. Chelonian Conserv. Biol. 2015, 14, 64–72. [Google Scholar] [CrossRef]
- Perazzo, G.X.; Garcez, D.K.; Trindade, C.R.; Pereira, K.M.; Tozetti, A.M. Is the presence of eggs a relevant cue for predators of freshwater chelonian nests? Neotrop. Biol. Conserv. 2018, 13, 350–355. [Google Scholar] [CrossRef]
- Congdon, J.D.; Tinkle, D.W.; Breitenbach, G.L.; van Loben Sels, R.C. Nesting ecology and hatching success in the turtle Emydoidea blandingi. Herpetol. 1983, 39, 417–429. Available online: https://www.jstor.org/stable/3892538 (accessed on 22 February 2016).
- Czaja, R.A.; Kanonik, A.; Burke, R.L. The effect of rainfall on predation of Diamond-backed Terrapin (Malaclemys terrapin) nests. J. Herpetol. 2018, 52, 402–405. [Google Scholar] [CrossRef]
- Báldi, A. The use of artificial nests for estimating rates of nest survival. Ornis Hung. 1999, 8, 39–55. Available online: http://www.ornis.hu/articles/OrnisHungarica_vol8-9_p39-55.pdf (accessed on 24 October 2022).
- Iverson, J.B.; Klondaris, H.; Angell, C.S.; Tori, W.P. Olfaction as a cue for nest-site choice in turtles. Chelonian Conserv. Biol. 2016, 15, 206–213. [Google Scholar] [CrossRef]
- Marchand, T.; Le Gal, A.S.; Georges, J.Y. Fine scale behaviour and time-budget in the cryptic ectotherm European pond turtle Emys orbicularis. PLoS ONE 2021, 16, e0256549. [Google Scholar] [CrossRef]
- Cadi, A.; Nemoz, M.; Thienpont, S.; Joly, P. Home range, movements, and habitat use of the European pond turtle (Emys orbicularis) in the Rhône-Alpes region, France. Biologia 2004, 59 (Suppl. S14), 89–94. [Google Scholar]
- Bennie, J.J.; Duffy, J.P.; Inger, R.; Gaston, K.J. Biogeography of time partitioning in mammals. Proc. Natl. Acad. Sci. USA 2014, 111, 13727–13732. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, D.W.; Brown, L.; Yerli, S.; Canbolat, A.F. Behavior of red foxes, Vulpes vulpes, caching eggs of loggerhead turtles, Caretta caretta. J. Mammal. 1994, 75, 985–988. [Google Scholar] [CrossRef]
- Reynolds, J.C.; Aebischer, N.J. Comparison and quantification of carnivore diet by faecal analysis: A critique, with recommendations, based on a study of the fox Vulpes vulpes. Mamm. Rev. 1991, 21, 97–122. [Google Scholar] [CrossRef]
- Burke, R.L.; Felice, S.M.; Sobel, S.G. Raccoon (Procyon lotor) predation behavior changes affects turtle (Malaclemys terrapin) nest censuses. Chelonian Conserv. Biol. 2009, 8, 209–211. [Google Scholar] [CrossRef]
- Lanszki, J. Diet of badgers living in a deciduous forest in Hungary. Mamm. Biol. 2004, 69, 354–358. [Google Scholar] [CrossRef]
- Lanszki, J.; Zalewski, A.; Horváth, G. Comparison of red fox Vulpes vulpes and pine marten Martes martes food habits in a deciduous forest in Hungary. Wildlife Biol. 2007, 13, 258–271. [Google Scholar] [CrossRef] [Green Version]
- Drobenkov, S.M. Reproductive ecology of the pond turtle (Emys orbicularis L.) in the northeastern part of the species range. Russ. J. Ecol. 2000, 31, 49–54. Available online: https://link.springer.com/article/10.1007/BF02799726 (accessed on 7 September 2022). [CrossRef]
- Bravo, C.; Pays, O.; Sarasa, M.; Bretagnolle, V. Revisiting an old question: Which predators eat eggs of ground-nesting birds in farmland landscapes? Sci. Total Environ. 2020, 244, 140895. [Google Scholar] [CrossRef]
- Long, R.A.; MacKay, P.; Ray, J.; Zielinski, W. (Eds.) Noninvasive Survey Methods for Carnivores; Island Press: Washington, DC, USA, 2012; pp. 1–400. [Google Scholar]
- Dawson, S.J.; Adams, P.; Huston, R.M.; Fleming, P.A. Environmental factors influence nest excavation by foxes. J. Zool. 2014, 294, 104–113. [Google Scholar] [CrossRef]
- Caravaggi, A.; Banks, P.B.; Burton, A.C.; Finlay, C.M.V.; Haswell, P.M.; Hayward, M.W.; Rowcliffe, M.J.; Wood, M.D. A review of camera trapping for conservation behaviour research. Remote Sens. Ecol. Conserv. 2017, 3, 109–122. [Google Scholar] [CrossRef]
- Christiansen, J.L.; Gallaway, B.J. Raccoon removal, nesting success, and hatchling emergence in Iowa turtles with special reference to Kinosternon flavescens (Kinosternidae). Southwest. Nat. 1984, 29, 343–348. [Google Scholar] [CrossRef]
- Spencer, R.J. Experimentally testing nest site selection: Fitness trade-offs and predation risk in turtles. Ecology 2002, 83, 2136–2144. [Google Scholar] [CrossRef]
- Chessman, B.C. Introduced red foxes (Vulpes vulpes) driving Australian freshwater turtles to extinction? A critical evaluation of the evidence. Pac. Conserv. Biol. 2021, 28, 462–471. [Google Scholar] [CrossRef]
- Escoriza, D.; Franch, M.; Ramos, S.; Sunyer-Sala, P.; Boix, D. Demographics and survivorship in the European pond turtle (Emys orbicularis): A 31-year study. Herpetol. Conserv. Biol. 2020, 15, 41–48. [Google Scholar]
- Bona, M.; Novotný, M.; Danko, S.; Burešová, A. Nest site fidelity in the Slovakian population of the European pond turtle Emys orbicularis. Amphibia-Reptilia 2012, 33, 207–213. [Google Scholar] [CrossRef]
- Purger, J.J.; Molnár, T.G. An unexpected recapture of European pond turtle (Emys orbicularis Linnaeus, 1758) in the Barcs Juniper woodland (Hungary). Nat. Somogy. 2022, 39, 5–10. [Google Scholar] [CrossRef]
- Mitrus, S. Fidelity to nesting area of the European pond turtle, Emys orbicularis Linnaeus, 1758). Belg. J. Zool. 2006, 136, 25–30. Available online: http://biblio.naturalsciences.be/associated_publications/bjz/136-1/volume-136-1-pp-25-30.pdf (accessed on 11 November 2018).
- Najbar, B.; Szuszkiewicz, E. Nest-site fidelity of the European pond turtle (Emys orbicularis Linnaeus, 1758) (Testudines: Emydidae) in western Poland. Acta Zool. Cracov. 2007, 50, 1–8. [Google Scholar] [CrossRef]
- Vilardell, A.; Capalleras, X.; Budó, J.; Pons, P. Predator identification and effects of habitat management and fencing on depredation rates of simulated nests of an endangered population of Hermann’s tortoises. Eur. J. Wildl. Res. 2012, 58, 707–713. [Google Scholar] [CrossRef]
- Buzuleciu, S.A.; Spencer, M.E.; Parker, S.L. Predator exclusion cage for turtle nests: A novel design. Chelonian Conserv. Biol. 2015, 14, 196–201. [Google Scholar] [CrossRef]
- Bocz, R.; Batáry, P.; Purger, J.J. Scent, rather than fur pattern, determines predation of mice: An in-the-wild experiment with plasticine mouse models. J. Zool. 2022, 316, 223–228. [Google Scholar] [CrossRef]
- Burke, R.L.; Schneider, C.M.; Dolinger, M.T. Cues used by raccoons to find turtle nests: Effects of flags, human scent, and diamond-backed terrapin sign. J. Herpetol. 2005, 39, 312–315. [Google Scholar] [CrossRef]
- Edmunds, S.; Kasparov, C.N.; Yoon, J.B.; Kanonik, A.; Burke, R.L. Twelve years later: Reassessing visual and olfactory cues raccoons use to find diamondback terrapin nests. J. Herpet. 2018, 52, 307–312. [Google Scholar] [CrossRef] [Green Version]
- Rollinson, N.; Brooks, R.J. Marking nests increases the frequency of nest depredation in a northern population of Painted Turtles (Chrysemys picta). J. Herpet. 2007, 41, 174–176. [Google Scholar] [CrossRef]
- Díaz-Ruiz, F.; Caro, J.; Delibes-Mateos, M.; Arroyo, B.; Ferreras, P. Drivers of red fox (Vulpes vulpes) daily activity: Prey availability, human disturbance or habitat structure? J. Zool. 2016, 298, 128–138. [Google Scholar] [CrossRef] [Green Version]
- Roshier, D.A.; Carter, A. Space use and interactions of two introduced mesopredators, European red fox and feral cat, in an arid landscape. Ecosphere 2021, 12, e03628. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purger, J.J.; Molnár, T.G.; Lanszki, Z.; Lanszki, J. European Pond Turtle (Emys orbicularis) Nest Predation: A Study with Artificial Nests. Biology 2023, 12, 342. https://doi.org/10.3390/biology12030342
Purger JJ, Molnár TG, Lanszki Z, Lanszki J. European Pond Turtle (Emys orbicularis) Nest Predation: A Study with Artificial Nests. Biology. 2023; 12(3):342. https://doi.org/10.3390/biology12030342
Chicago/Turabian StylePurger, Jenő J., Tamás Gergely Molnár, Zsófia Lanszki, and József Lanszki. 2023. "European Pond Turtle (Emys orbicularis) Nest Predation: A Study with Artificial Nests" Biology 12, no. 3: 342. https://doi.org/10.3390/biology12030342