
_Sun.png)


Thermal Profiles of Chainsaw Hollows and Natural Hollows during Extreme Heat Events

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Hollow Creation
2.2. Monitoring Cavity Temperatures
2.3. Statistical Analyses
3. Results
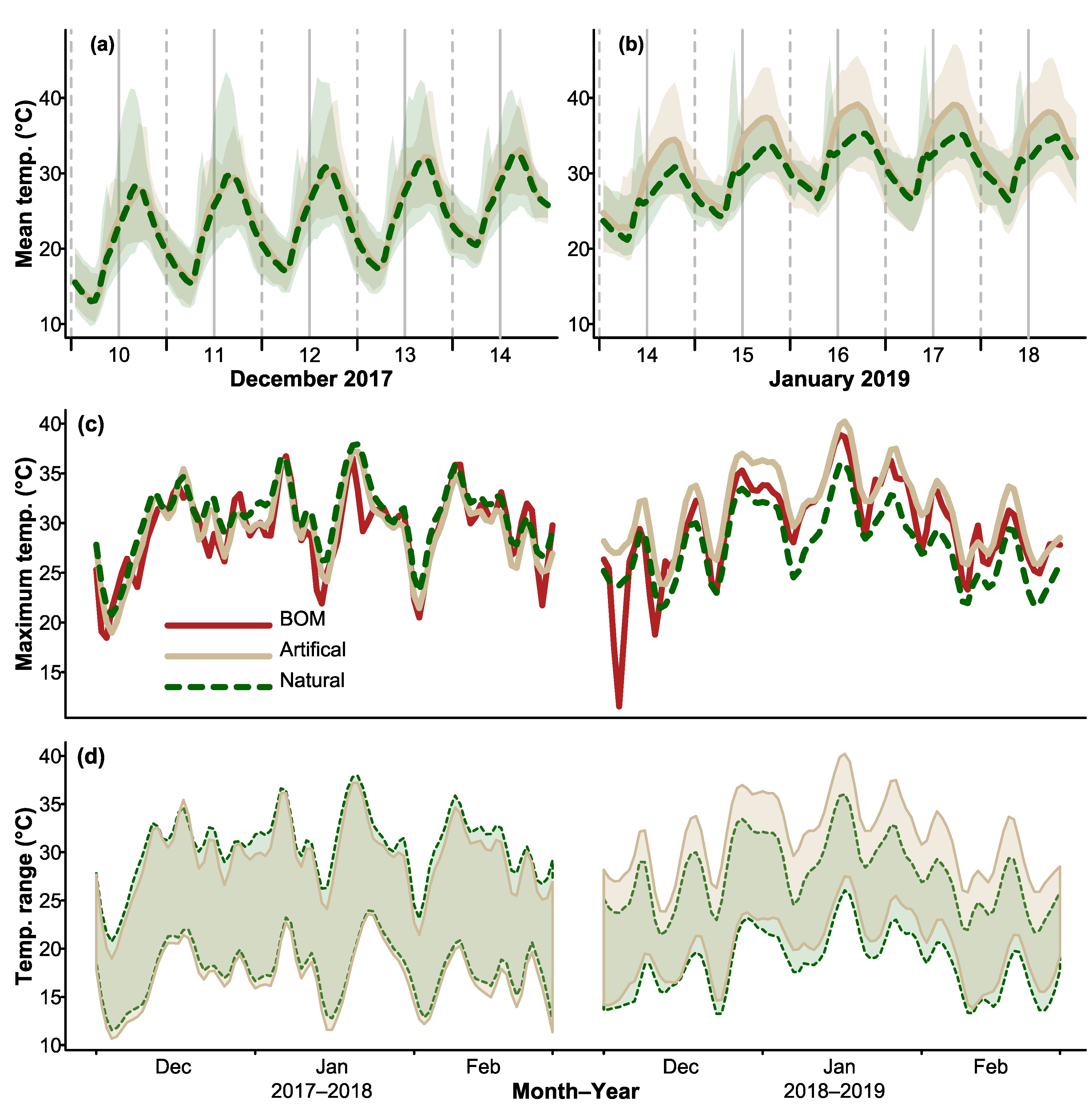
3.1. Daily Thermal Profiles of CHs and Natural Hollows
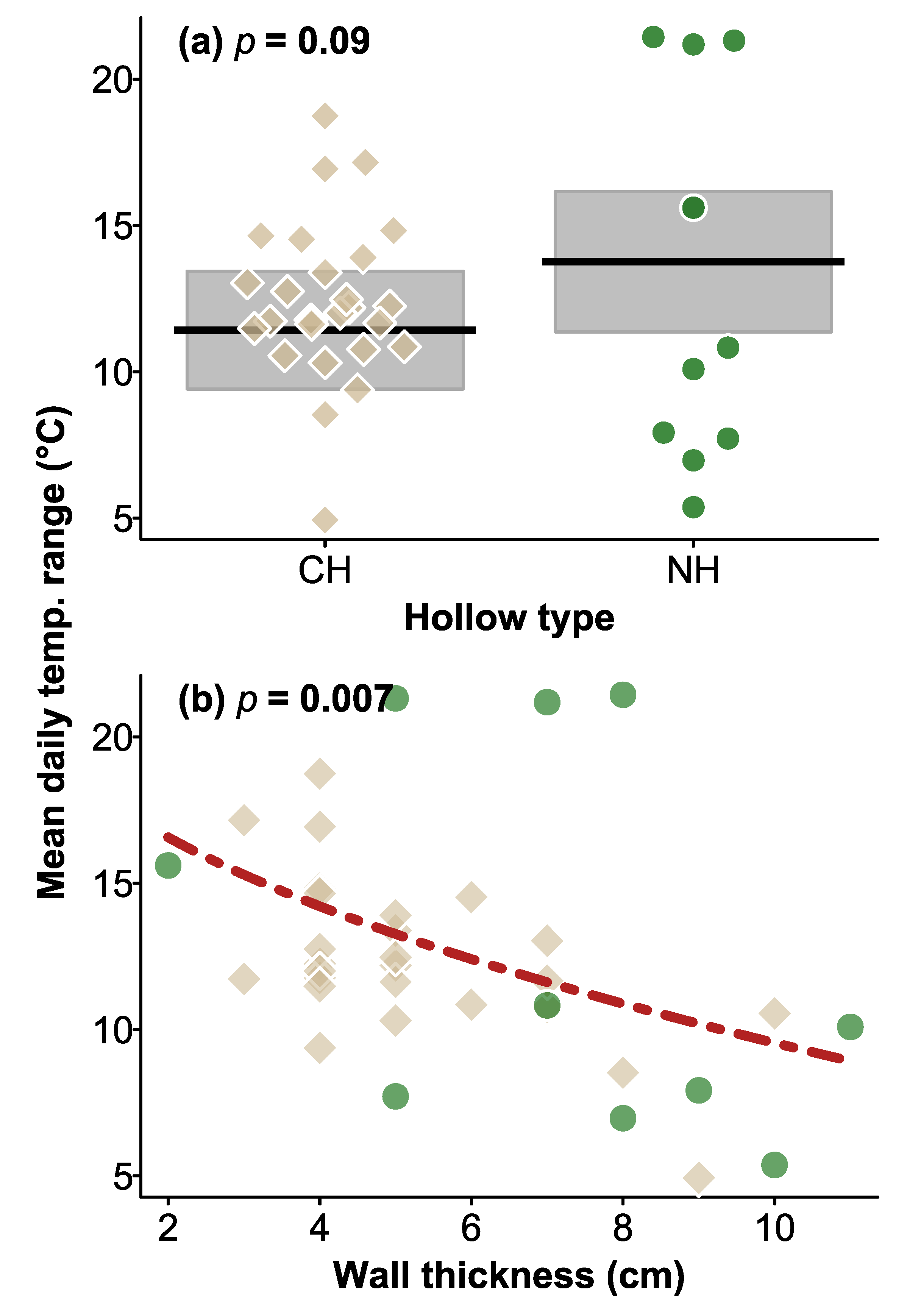
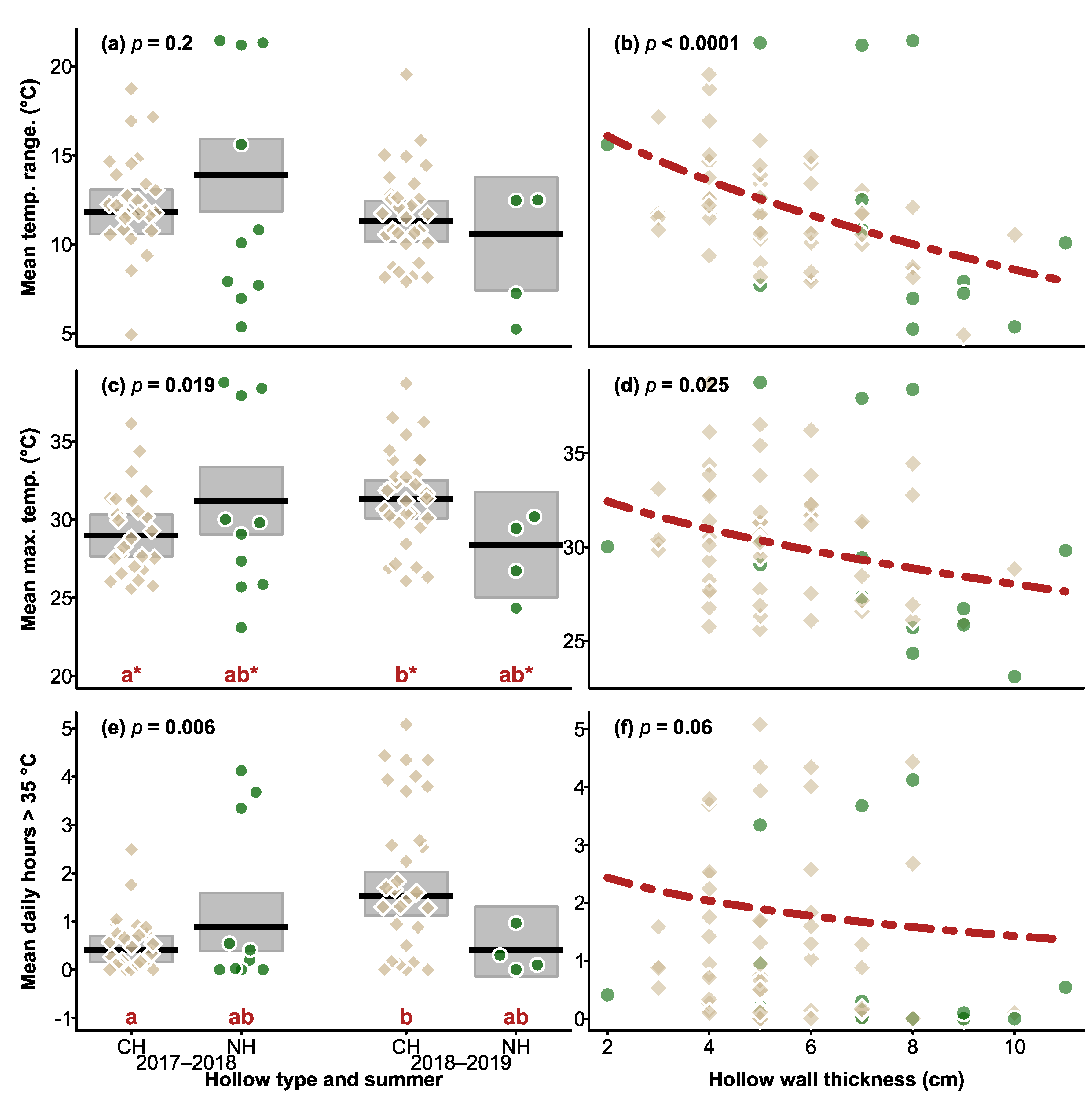
3.2. Daily Temperature Ranges in CHs and Natural Hollows
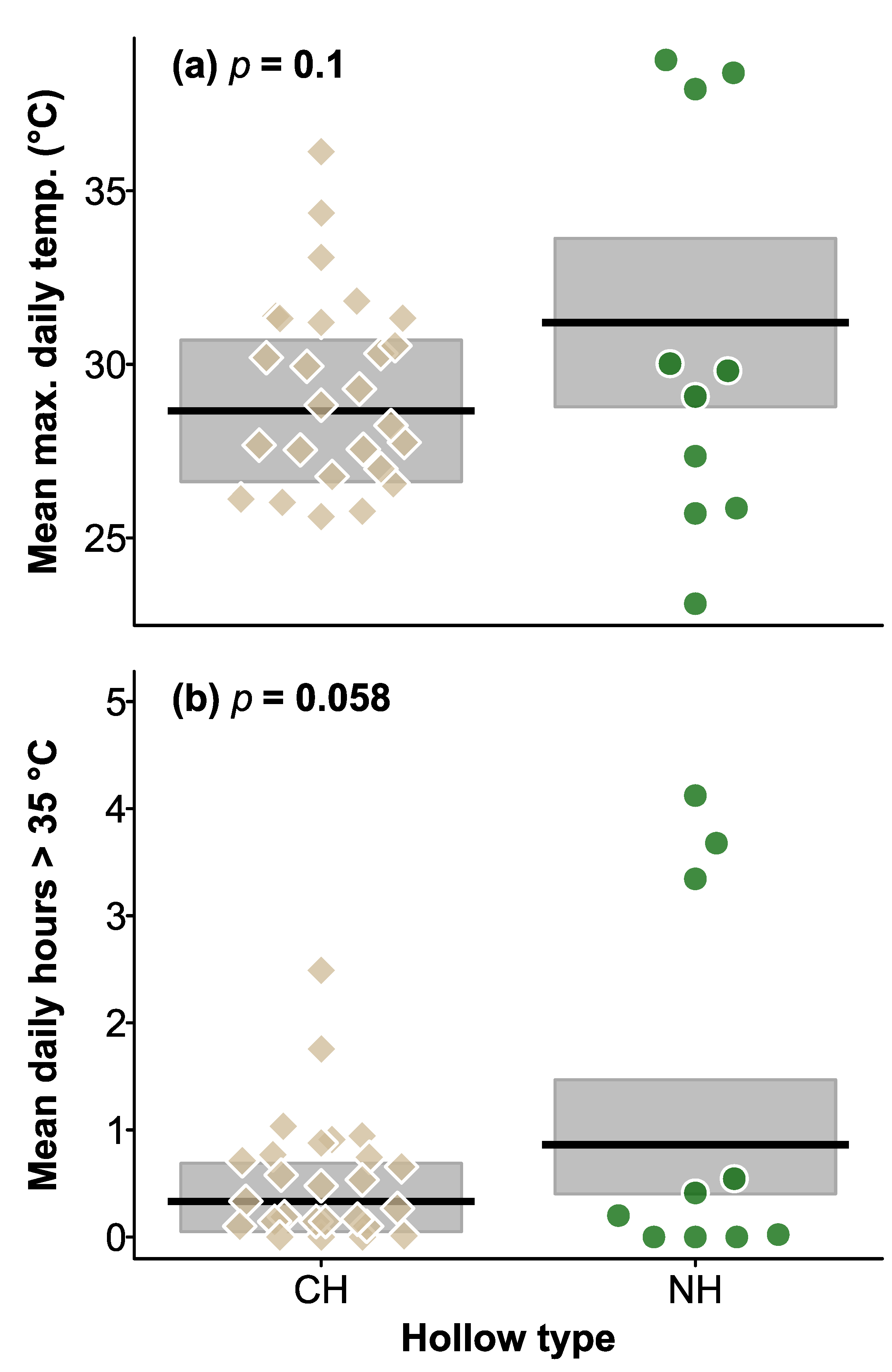
3.3. Maximum Daily Temperatures in CHs and Natural Hollows
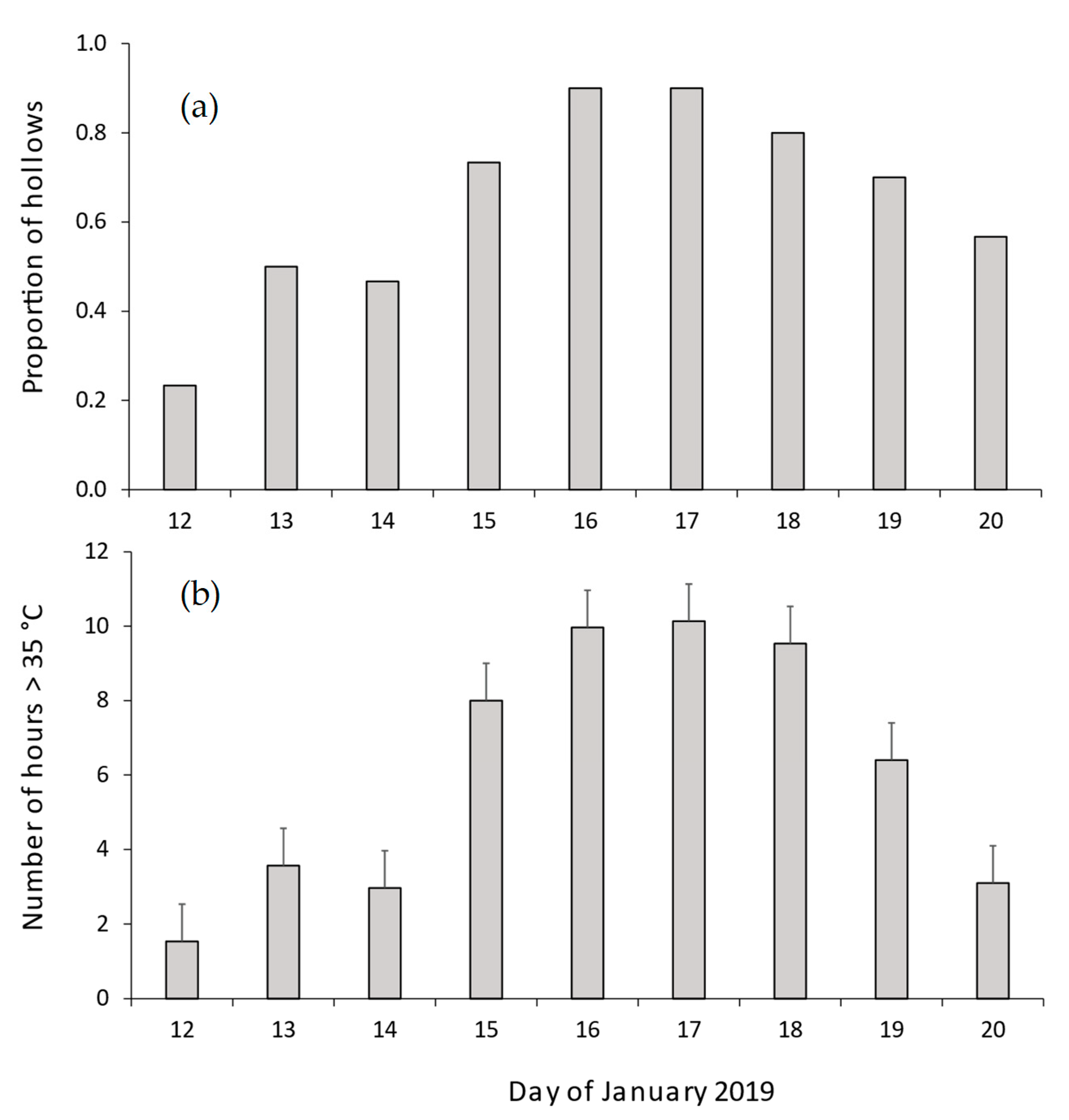
3.4. Number of Hours > 35 °C in Natural Hollows and CHs in Summer, and during the 2019 Heatwave
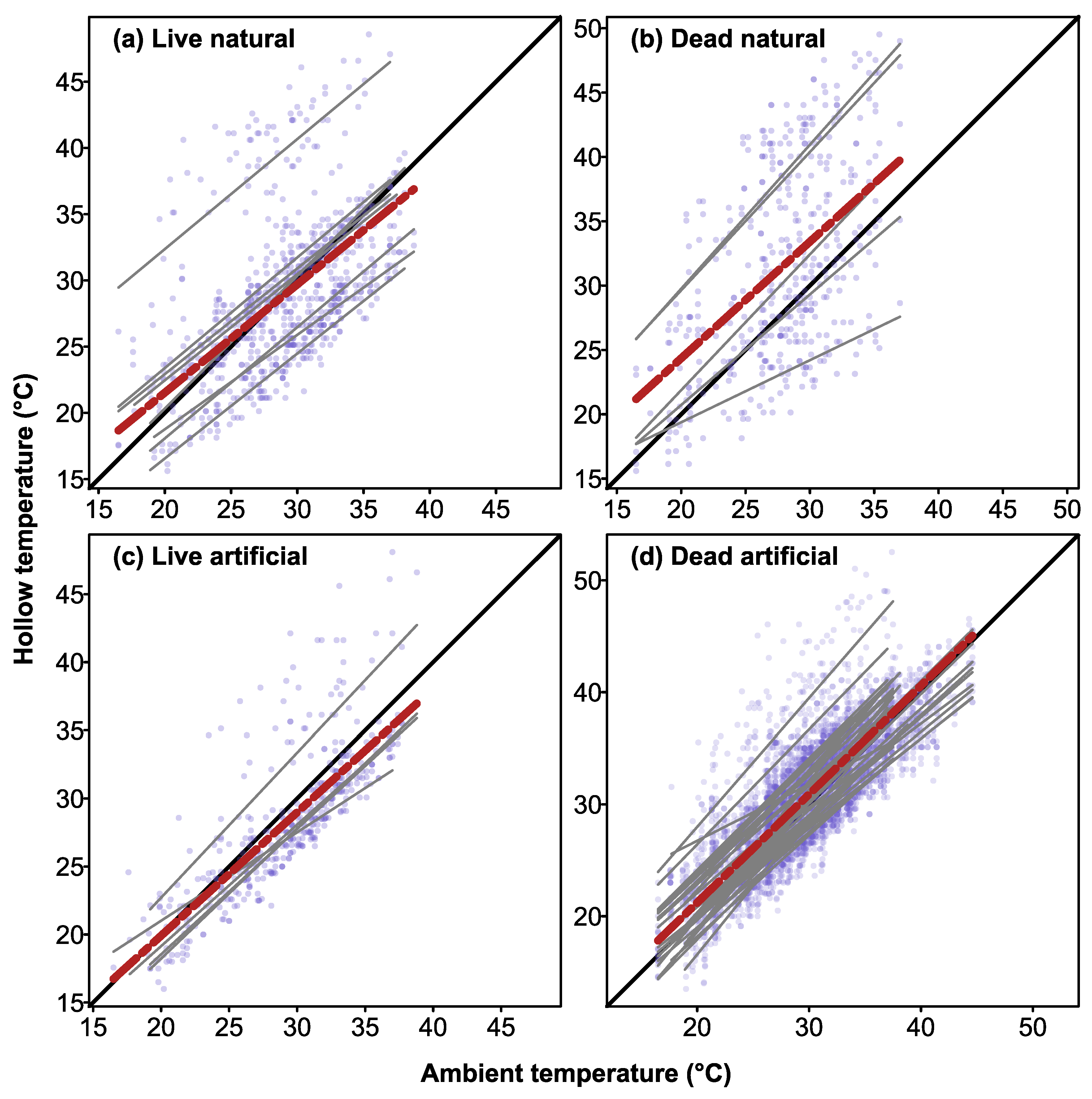
3.5. Thermal Buffering within Hollows
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bradshaw, C.J.A. Little left to lose: Deforestation and forest degradation in Australia since European colonization. J. Plant Ecol. 2012, 5, 109–120. [Google Scholar] [CrossRef]
- Evans, M.C. Deforestation in Australia: Drivers, trends, and policy responses. Pac. Conserv. Biol. 2016, 22, 130–150. [Google Scholar] [CrossRef]
- Ozolins, A.; Brack, C.; Freudenberger, D. Abundance and decline of isolated trees in the agricultural landscapes of central New South Wales, Australia. Pac. Conserv. Biol. 2001, 7, 195–203. [Google Scholar] [CrossRef]
- Walker, J.; Bullen, F.; Williams, B. Ecohydrological changes in the Murray-Darling Basin. I. The number of trees cleared over two centuries. J. Appl. Ecol. 1993, 30, 265–273. [Google Scholar] [CrossRef]
- Koch, A.; Munks, S.; Driscoll, D. The use of hollow-bearing trees by vertebrate fauna in wet and dry. Wildl. Res. 2008, 35, 727–746. [Google Scholar] [CrossRef]
- Gibbons, P.; Lindenmayer, D.; Barry, S.C.; Tanton, M.T. Hollow selection by vertebrate fauna in forests of southeastern Australia and implications for forest management. Biol. Conserv. 2002, 103, 1–12. [Google Scholar] [CrossRef]
- Mackowski, C.M. The ontogeny of hollow in blackbutt (Eucalyptus pilularis) and its relevance to the management of forests for possums, gliders and timber. In Possums and Gliders; Smith, A., Hume, I., Eds.; Surrey Beatty & Sons Pty Limited: Chipping Norton, Australia, 1984; pp. 553–567. [Google Scholar]
- Wormington, K.R.; Lamb, D.; McCallum, H.I.; Moloney, D.J. Habitat requirements for the conservation of arboreal marsupials in dry sclerophyll forests of Southeast Queensland, Australia. For. Sci. 2002, 48, 217–227. [Google Scholar]
- Lindenmayer, D.B.; Blanchard, W.; Blair, D.; McBurney, L.; Taylor, C.; Scheele, B.C.; Westgate, M.J.; Robinson, N.; Foster, C. The response of arboreal marsupials to long-term changes in forest disturbance. Anim. Conserv. 2021, 24, 246–258. [Google Scholar] [CrossRef]
- McLean, C.M.; Kavanagh, R.P.; Penman, T.; Bradstock, R. The threatened status of the hollow dependent arboreal marsupial, the Greater Glider (Petauroides volans), can be explained by impacts from wildfire and selective logging. For. Ecol. Manag. 2018, 415, 19–25. [Google Scholar] [CrossRef]
- NSW Scientific Committee. Loss of Hollow-Bearing Trees—Key Threatening Process Listing. Available online: https://www.environment.nsw.gov.au/topics/animals-and-plants/threatened-species/nsw-threatened-species-scientific-committee/determinations/final-determinations/2004-2007/loss-of-hollow-bearing-trees-key-threatening-process-listing (accessed on 5 August 2022).
- Le Roux, D.S.; Ikin, K.; Lindenmayer, D.B.; Bistricer, G.; Manning, A.D.; Gibbons, P. Effects of entrance size, tree size and landscape context on nest box occupancy: Considerations for management and biodiversity offsets. For. Ecol. Manag. 2016, 366, 135–142. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Crane, M.; Evans, M.C.; Maron, M.; Gibbons, P.; Bekessy, S.; Blanchard, W. The anatomy of a failed offset. Biol. Conserv. 2017, 210, 286–292. [Google Scholar] [CrossRef]
- Rueegger, N. Artificial tree hollow creation for cavity-using wildlife—Trialling an alternative method to that of nest boxes. For. Ecol. Manag. 2017, 405, 404–412. [Google Scholar] [CrossRef]
- Griffiths, S.R.; Lentini, P.E.; Semmens, K.; Watson, S.J.; Lumsden, L.F.; Robert, K.A. Chainsaw-carved cavities better mimic the thermal properties of natural tree hollows than nest boxes and log hollows. Forests 2018, 9, 235. [Google Scholar] [CrossRef] [Green Version]
- Rowland, J.A.; Briscoe, N.J.; Handasyde, K.A. Comparing the thermal suitability of nest-boxes and tree-hollows for the conservation-management of arboreal marsupials. Biol. Conserv. 2017, 209, 341–348. [Google Scholar] [CrossRef]
- McComb, W.C.; Noble, R.E. Microclimates of nest boxes and natural cavities in bottomland hardwoods. J. Wildl. Manag. 1981, 45, 284–289. [Google Scholar] [CrossRef]
- Honey, R.; McLean, C.; Murray, B.R.; Webb, J.K. Insulated nest boxes provide thermal refuges for wildlife in urban bushland during summer heatwaves. J. Urban Ecol. 2021, 7, juab032. [Google Scholar] [CrossRef]
- Griffiths, S.R. Overheating turns a bat box into a death trap. Pac. Conserv. Biol. 2022, 28, 97–98. [Google Scholar] [CrossRef]
- Carey, A.B.; Gill, J.D. Direct habitat improvements—Some recent advances. In Snag Habitat Management: Proceedings of the Symposium. USDA Forest Service General Technical Report RM-99; USDA Forest Service: Fort Collins, CO, USA, 1983; pp. 80–87. [Google Scholar]
- Gano, R.D.; Mosher, J.A. Artificial cavity construction: And alternative to nest boxes. Wildl. Soc. Bull. 1983, 11, 74–76. [Google Scholar]
- Griffiths, S.R.; Semmens, K.; Watson, S.J.; Jones, C.S. Installing chainsaw-carved hollows in medium-sized live trees increases rates of visitation by hollow-dependent fauna. Restor. Ecol. 2020, 28, 1225–1236. [Google Scholar] [CrossRef]
- Best, K.; Haslem, A.; Maisey, A.C.; Semmens, K.; Griffiths, S.R. Occupancy of chainsaw-carved hollows by an Australian arboreal mammal is influenced by cavity attributes and surrounding habitat. For. Ecol. Manag. 2022, 503, 119747. [Google Scholar] [CrossRef]
- Honey, R.; McLean, C.M.; Murray, B.R.; Callan, M.N.; Webb, J.K. Choice of monitoring method can influence estimates of usage of artificial hollows by vertebrate fauna. Aust. J. Zool. 2021, 69, 18–25. [Google Scholar] [CrossRef]
- Terry, W.; Goldingay, R.L.; van der Ree, R. Can chainsaw carved hollows provide an effective solution to the loss of natural tree cavities for arboreal mammals? For. Ecol. Manag. 2021, 490, 119122. [Google Scholar] [CrossRef]
- Griffiths, S.R.; Robert, K.A.; Jones, C.S. Chainsaw hollows carved into live trees provide well insulated supplementary shelters for wildlife during extreme heat. Wildl. Res. 2022, 49, 596–609. [Google Scholar] [CrossRef]
- Ikin, K.; Mortelliti, A.; Stein, J.; Michael, D.; Crane, M.; Okada, S.; Wood, J.; Lindenmayer, D. Woodland habitat structures are affected by both agricultural land management and abiotic conditions. Landsc. Ecol. 2015, 30, 1387–1403. [Google Scholar] [CrossRef]
- Edwards, N.T.; Hanson, P.J. Stem respiration in a closed-canopy upland oak forest. Tree Physiol. 1996, 16, 433–439. [Google Scholar] [CrossRef]
- Keith, D. Ocean Shores to Desert Dunes: The Native Vegetation of New South Wales and the ACT; Department of Environment and Conservation (NSW): Sydney, Australia, 2006. [Google Scholar]
- Kane, B.C.P.; Ryan, H.D.P.I. The accuracy of formulas used to assess strength loss due to decay in trees. J. Arboric. 2004, 30, 347–356. [Google Scholar] [CrossRef]
- Mattheck, C.; Bethge, K.; Erb, D. Failure criteria for trees. Arboric. J. 1993, 17, 201–209. [Google Scholar] [CrossRef]
- Kenyon, P.; Kenyon, P. Pruning for Habitat Workshop: Habitat Creation; Victorian Tree Industry Organisation: Selby, Australia, 2010. [Google Scholar]
- Perkins, S.E.; Alexander, L.V. On the Measurement of Heat Waves. J. Clim. 2013, 26, 4500–4517. [Google Scholar] [CrossRef]
- Dawson, T.J. Temperature regulation and evaporative water loss in brush-tailed possum Trichosurus vulpecula. Comp. Biochem. Physiol. 1969, 28, 401–407. [Google Scholar] [CrossRef]
- Fleming, M.R. Thermoregulation and torpor in the sugar glider, Petaurus breviceps (Marsupialia, Petauridae). Aust. J. Zool. 1980, 28, 521–534. [Google Scholar] [CrossRef]
- Holloway, J.C.; Geiser, F. Effects of helium/oxygen and temperature on aerobic metabolism in the marsupial sugar glider, Petaurus breviceps. Physiol. Biochem. Zool. 2001, 74, 219–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, M.R. The thermal physiology of the feathertail glider, Acrobates pygmaeus (Marsupialia, Burramyidae). Aust. J. Zool. 1985, 33, 667–681. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Eklund, A.; Trimble, J. Beeswarm: The Bee Swarm Plot, an Alternative to Stripchart. 2021. Available online: https://rdrr.io/cran/beeswarm/ (accessed on 21 July 2022).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means. 2022. Available online: https://cran.r-project.org/web/packages/emmeans/emmeans.pdf (accessed on 21 July 2022).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Wang, H.; Guan, H.; Guyot, A.; Simmons, C.T.; Lockington, D.A. Quantifying sapwood width for three Australian native species using electrical resistivity tomography. Ecohydrology 2016, 9, 83–92. [Google Scholar] [CrossRef]
- Bowman, W.P.; Turnbull, M.H.; Tissue, D.T.; Whitehead, D.; Griffin, K.L. Sapwood temperature gradients between lower stems and the crown do not influence estimates of stand-level stem CO2 efflux. Tree Physiol. 2008, 28, 1553–1559. [Google Scholar] [CrossRef] [Green Version]
- Stockfors, J. Temperature variations and distribution of living cells within tree stems: Implications for stem respiration modeling and scale-up. Tree Physiol. 2000, 20, 1057–1062. [Google Scholar] [CrossRef]
- Robinson, K.W.; Morrison, P.R. The reaction to hot atmospheres of various species of Australian marsupial and placental animals. J. Cell. Comp. Physiol. 1957, 49, 455–478. [Google Scholar] [CrossRef]
- Goldingay, R.L.; Thomas, K.J. Tolerance to high temperature by arboreal mammals using nest boxes in southern Australia. J. Therm. Biol. 2021, 98, 102899. [Google Scholar] [CrossRef]
- Turner, J.M. Facultative hyperthermia during a heatwave delays injurious dehydration of an arboreal marsupial. J. Exp. Biol. 2020, 223, jeb219378. [Google Scholar] [CrossRef]
- O’Connell, C.; Keppel, G. Deep tree hollows: Important refuges from extreme temperatures. Wildl. Biol. 2016, 22, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Webb, J.K.; Shine, R. Out on a limb: Conservation implications of tree-hollow use by a threatened snake species (Hoplocephalus bungaroides: Serpentes, Elapidae). Biol. Conserv. 1997, 81, 21–33. [Google Scholar] [CrossRef]
- Beyer, G.L.; Goldingay, R.L.; Sharpe, D.J. The characteristics of squirrel glider (Petaurus norfolcensis) den trees in subtropical Australia. Aust. J. Zool. 2008, 56, 13–21. [Google Scholar] [CrossRef]
- Ellis, M.V.; Taylor, J.E.; Rhind, S.G. Creating entrances to tree cavities attracts hollow-dependent fauna: Proof of concept. Restor. Ecol. 2022, 30, e13713. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Callan, M.N.; Krix, D.; McLean, C.M.; Murray, B.R.; Webb, J.K. Thermal Profiles of Chainsaw Hollows and Natural Hollows during Extreme Heat Events. Biology 2023, 12, 361. https://doi.org/10.3390/biology12030361
Callan MN, Krix D, McLean CM, Murray BR, Webb JK. Thermal Profiles of Chainsaw Hollows and Natural Hollows during Extreme Heat Events. Biology. 2023; 12(3):361. https://doi.org/10.3390/biology12030361
Chicago/Turabian StyleCallan, Michael N., Dan Krix, Christopher M. McLean, Brad R. Murray, and Jonathan K. Webb. 2023. "Thermal Profiles of Chainsaw Hollows and Natural Hollows during Extreme Heat Events" Biology 12, no. 3: 361. https://doi.org/10.3390/biology12030361
APA StyleCallan, M. N., Krix, D., McLean, C. M., Murray, B. R., & Webb, J. K. (2023). Thermal Profiles of Chainsaw Hollows and Natural Hollows during Extreme Heat Events. Biology, 12(3), 361. https://doi.org/10.3390/biology12030361