
Tripeptide IRW Improves AMPK/eNOS Signaling Pathway via Activating ACE2 in the Aorta of High-Fat-Diet-Fed C57BL/6 Mice

Abstract
:Simple Summary
Abstract
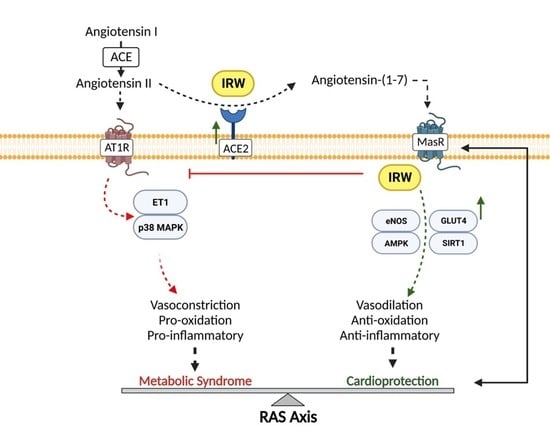
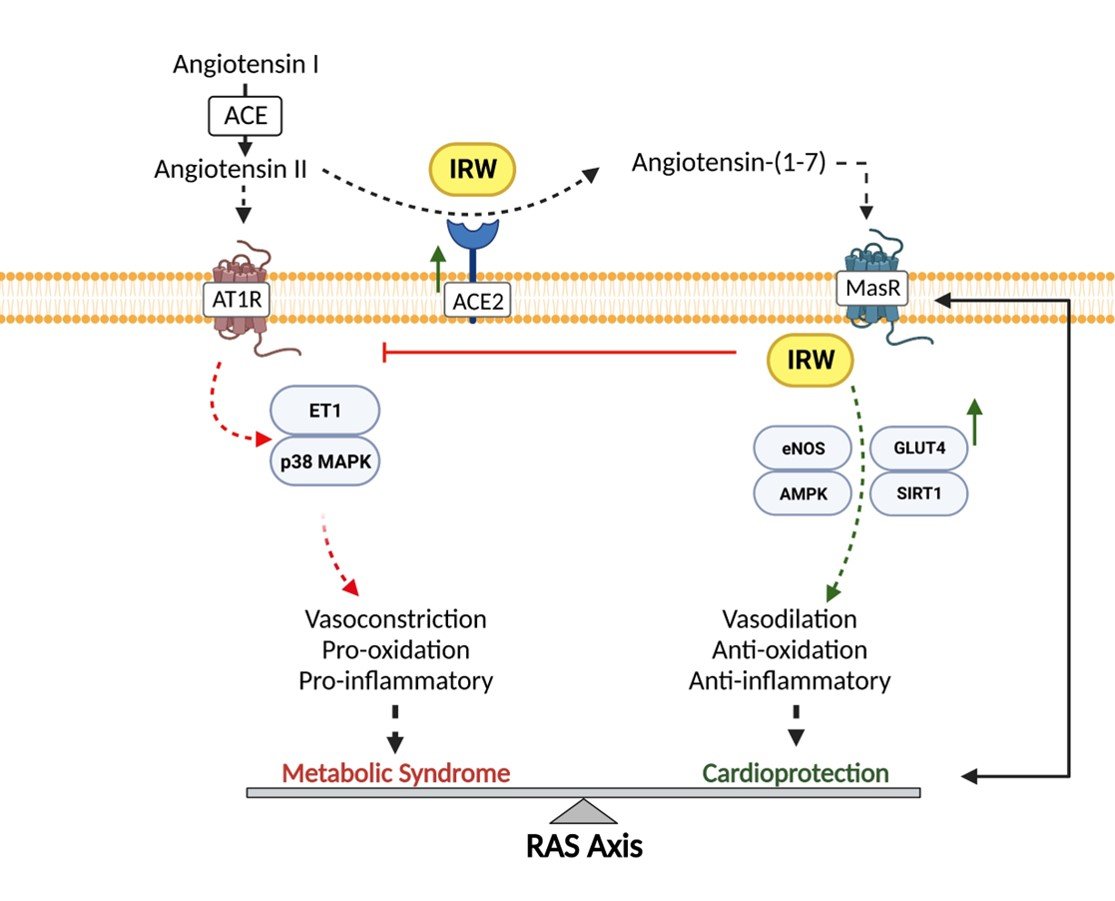{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animal Model Study
2.3. Tissue Collection
2.4. Protein Extraction and Western Blotting
2.5. Cell Culture
2.6. SiRNA Transfection
2.7. RT-PCR
2.8. Statistics
3. Results
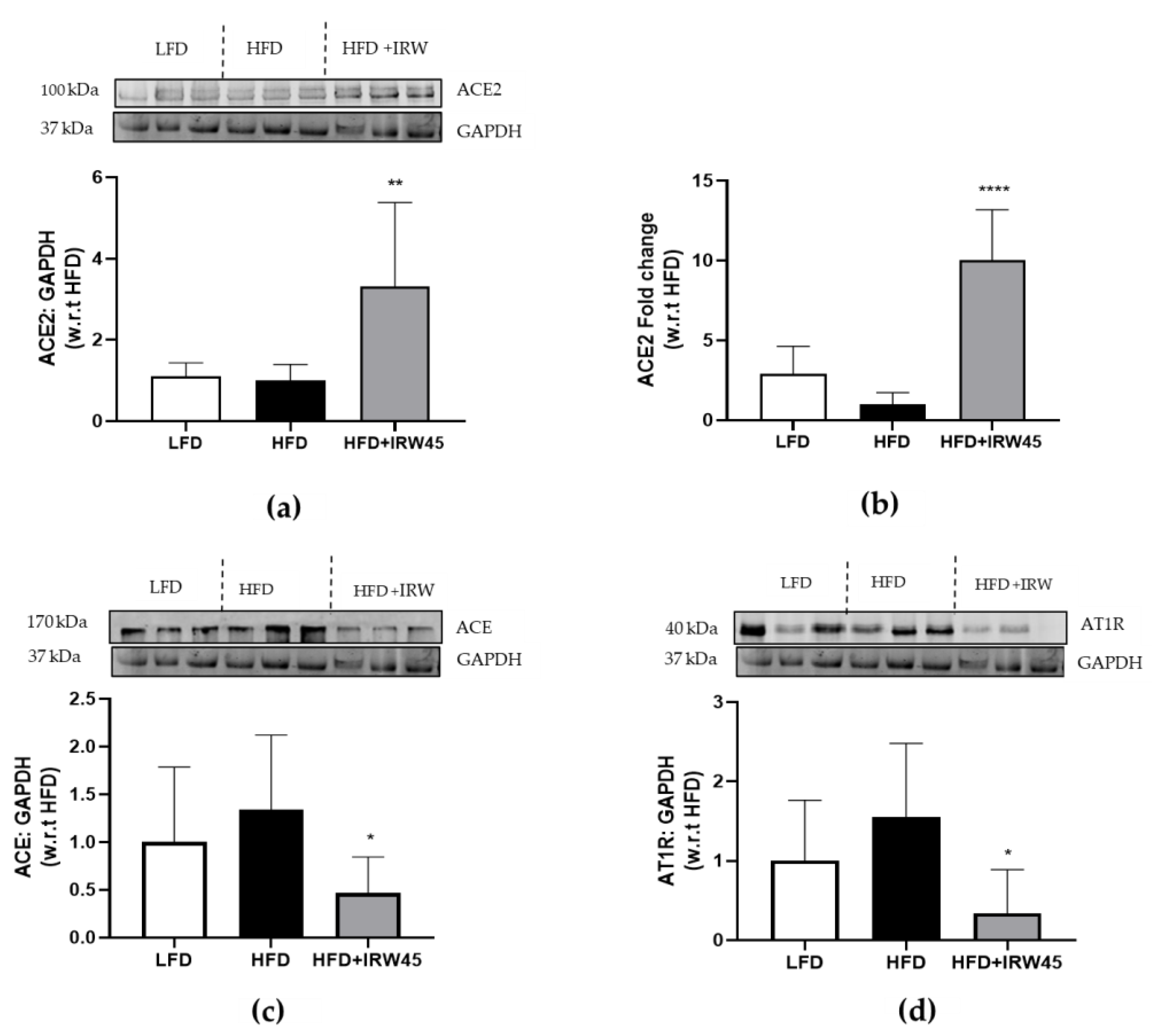
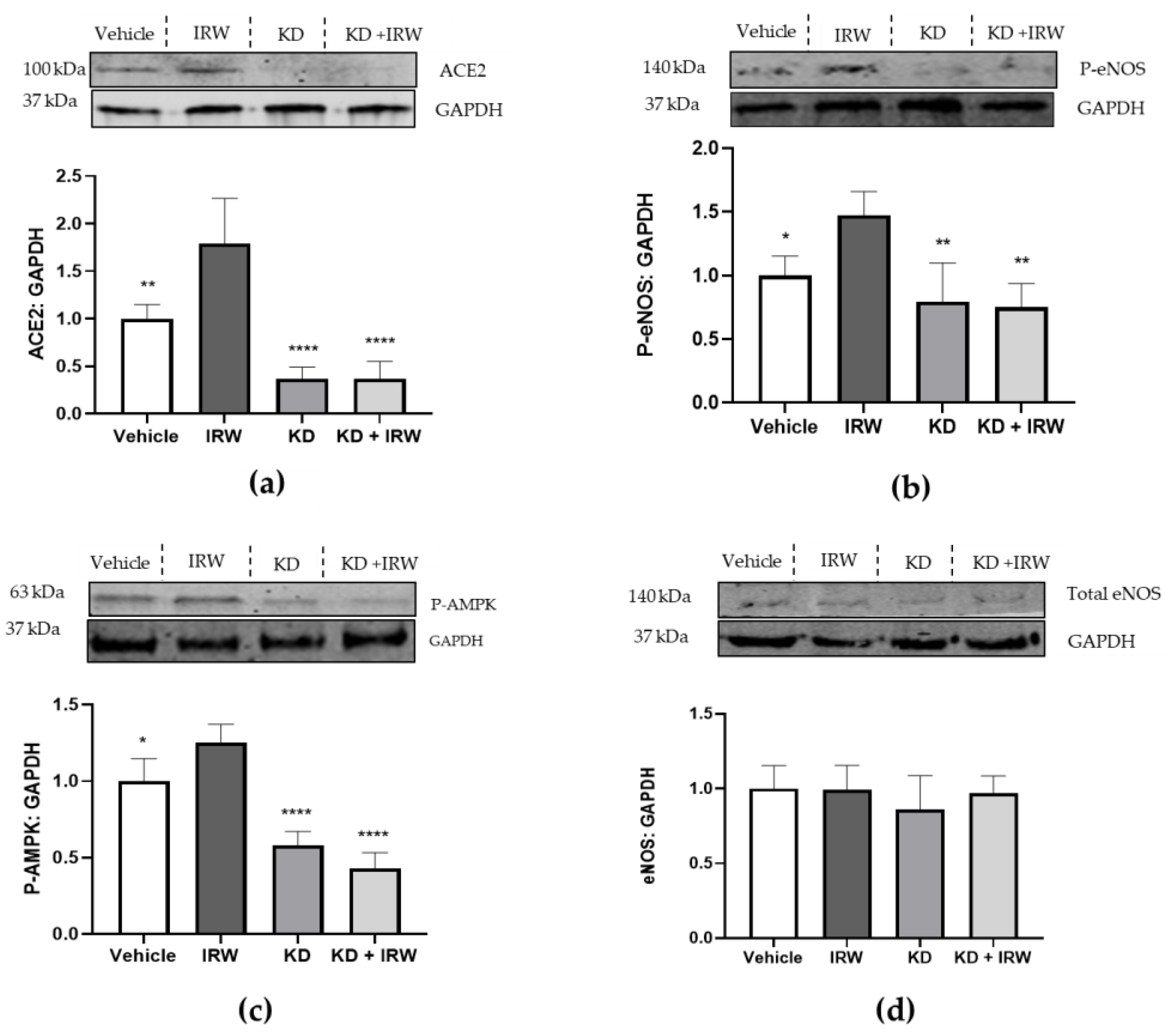
3.1. IRW Treatment Upregulated ACE2 and Diminished ACE and AT1R Expression in the Aorta
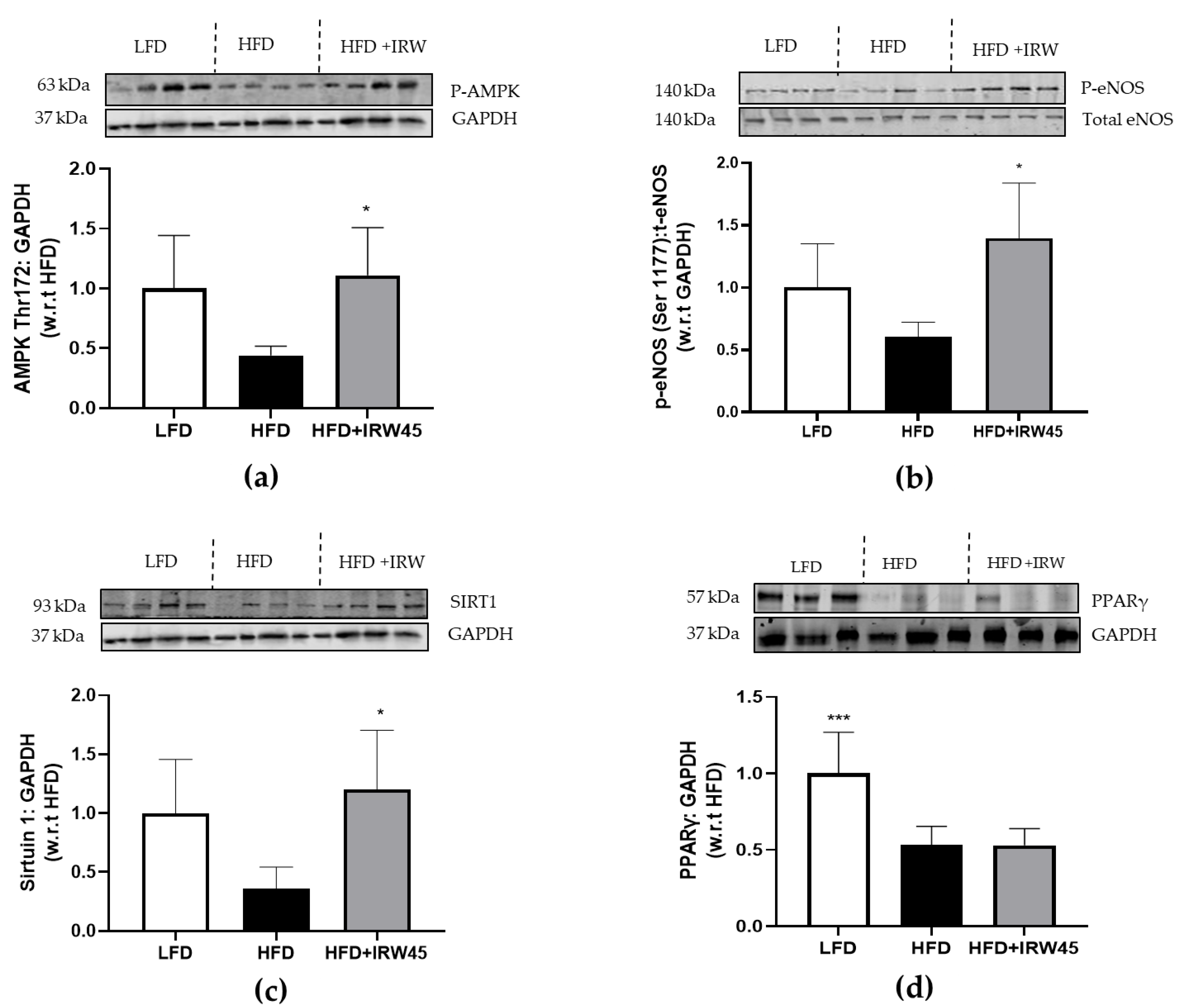
3.2. IRW Enhanced AMPK/SIRT1/eNOS Cascade in Aorta of HFD Mice via Aortic ACE2 Activation
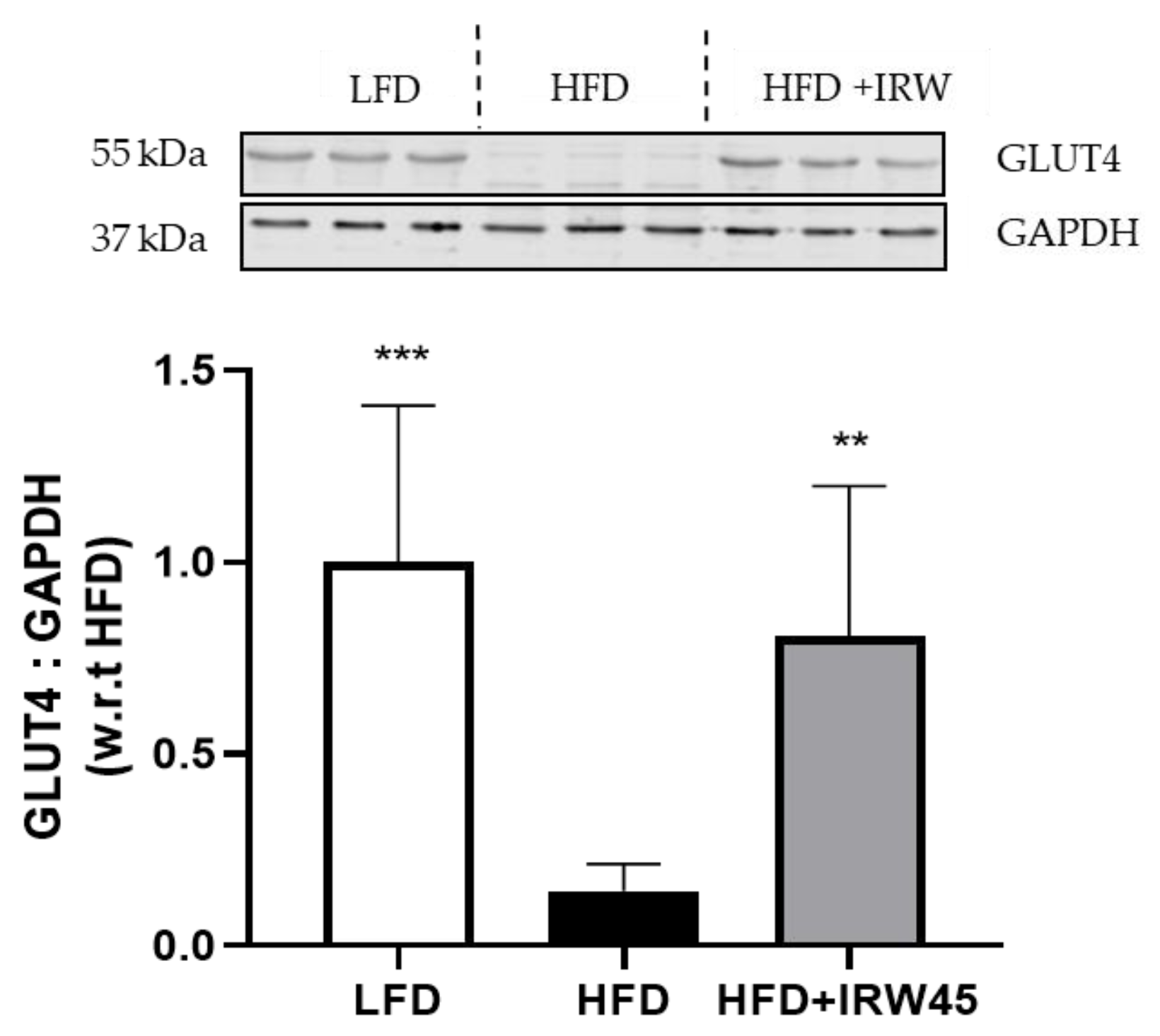
3.3. IRW Improved GLUT4 in Aorta of HFD Mice
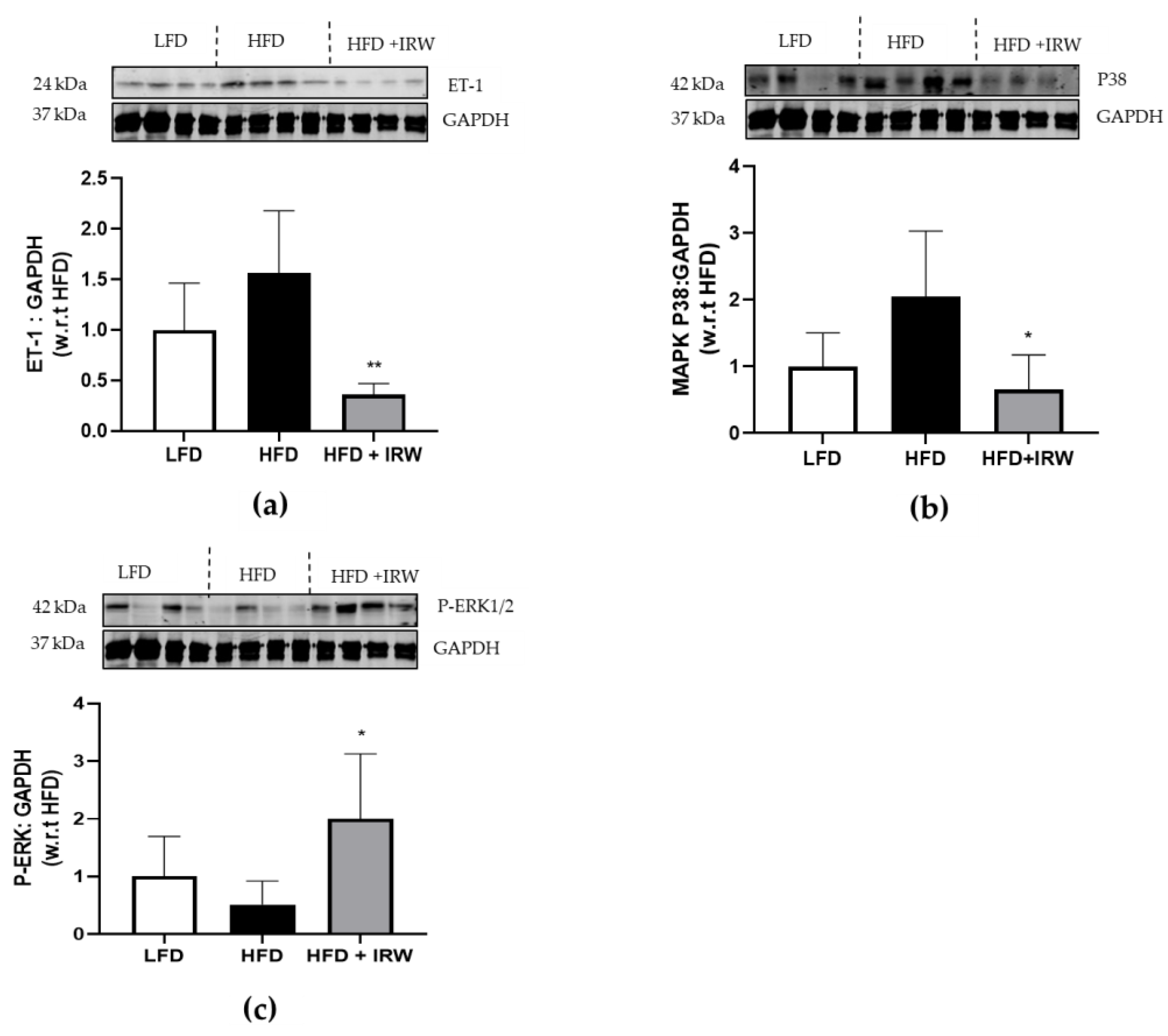
3.4. IRW Downregulated ET1/MAPK Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jahandideh, F.; Wu, J. A review on mechanisms of action of bioactive peptides against glucose intolerance and insulin resistance. Food Sci. Hum. Wellness. 2022, 11, 1441–1454. [Google Scholar] [CrossRef]
- Grundy, S.M. Metabolic syndrome update. Trends Cardiovasc. Med. 2016, 26, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Marcus, Y.; Shefer, G.; Stern, N. Adipose tissue renin–angiotensin–aldosterone system (RAAS) and progression of insulin resistance. Mol. Cell. Endocrinol. 2013, 378, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Chan, C.B.; Wu, J. Egg white ovotransferrin-derived ACE inhibitory peptide ameliorates angiotensin II-stimulated insulin resistance in skeletal muscle cells. Mol. Nutr. Food Res. 2018, 62, 1700602. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Fan, H.; Davidge, S.T.; Wu, J. Egg white–derived antihypertensive peptide IRW (Ile-Arg-Trp) reduces blood pressure in spontaneously hypertensive rats via the ACE2/ang (1–7)/mas receptor Axis. Mol. Nutr. Food Res. 2019, 63, 1900063. [Google Scholar] [CrossRef] [Green Version]
- Serfozo, P.; Wysocki, J.; Gulua, G.; Schulze, A.; Ye, M.; Liu, P.; Jin, J.; Bader, M.; Myöhänen, T.; García-Horsman, J.A. Ang II (angiotensin II) conversion to angiotensin-(1–7) in the circulation is POP (prolyloligopeptidase)-dependent and ACE2 (angiotensin-converting enzyme 2)-independent. Hypertension 2020, 75, 173–182. [Google Scholar] [CrossRef]
- Jahandideh, F.; Wu, J. Perspectives on the potential benefits of antihypertensive peptides towards metabolic syndrome. Int. J. Mol. Sci. 2020, 21, 2192. [Google Scholar] [CrossRef] [Green Version]
- Wu, J. A novel angiotensin converting enzyme 2 (ACE2) activating peptide: A reflection of 10 years of research on a small peptide Ile-Arg-Trp (IRW). J. Agric. Food Chem. 2020, 68, 14402–14408. [Google Scholar] [CrossRef]
- Li, S.; Liu, L.; He, G.; Wu, J. Molecular targets and mechanisms of bioactive peptides against metabolic syndromes. Food Funct. 2018, 9, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Bhullar, K.S.; Chakrabarti, S.; Davidge, S.T.; Wu, J. Egg white-derived tripeptide IRW (Ile-Arg-Trp) is an activator of angiotensin converting enzyme 2. J. Agric. Food Chem. 2018, 66, 11330–11336. [Google Scholar] [CrossRef]
- Liao, W.; Fan, H.; Wu, J. Egg white-derived antihypertensive peptide IRW (Ile-Arg-Trp) inhibits angiotensin II-stimulated migration of vascular smooth muscle cells via angiotensin type I receptor. J. Agric. Food Chem. 2018, 66, 5133–5138. [Google Scholar] [CrossRef] [PubMed]
- Atkins, K.B.; Johns, D.; Watts, S.; Webb, R.C.; Brosius III, F.C. Decreased vascular glucose transporter expression and glucose uptake in DOCA-salt hypertension. J. Hypertens. 2001, 19, 1581–1587. [Google Scholar] [CrossRef] [PubMed]
- Marcus, R.; England, R.; Nguyen, K.; Charron, M.J.; Briggs, J.P.; Brosius, F., 3rd. Altered renal expression of the insulin-responsive glucose transporter GLUT4 in experimental diabetes mellitus. Am. J. Physiol. Renal Physiol. 1994, 267, 816–824. [Google Scholar] [CrossRef]
- Park, J.L.; Loberg, R.D.; Duquaine, D.; Zhang, H.; Deo, B.K.; Ardanaz, N.; Coyle, J.; Atkins, K.B.; Schin, M.; Charron, M.J. GLUT4 facilitative glucose transporter specifically and differentially contributes to agonist-induced vascular reactivity in mouse aorta. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1596–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Campos Zani, S.C.; Son, M.; Bhullar, K.S.; Chan, C.B.; Wu, J. IRW (Isoleucine–Arginine–Tryptophan) Improves Glucose Tolerance in High Fat Diet Fed C57BL/6 Mice via Activation of Insulin Signaling and AMPK Pathways in Skeletal Muscle. Biomedicines 2022, 10, 1235. [Google Scholar] [CrossRef]
- van den Oever, I.A.; Raterman, H.G.; Nurmohamed, M.T.; Simsek, S. Endothelial dysfunction, inflammation, and apoptosis in diabetes mellitus. Mediators Inflamm. 2010, 2010, 792393. [Google Scholar] [CrossRef] [Green Version]
- van Sloten, T.T.; Henry, R.M.; Dekker, J.M.; Nijpels, G.; Unger, T.; Schram, M.T.; Stehouwer, C.D. Endothelial dysfunction plays a key role in increasing cardiovascular risk in type 2 diabetes: The Hoorn study. Hypertension 2014, 64, 1299–1305. [Google Scholar] [CrossRef] [Green Version]
- Elzinga, S.E.; Savelieff, M.G.; O’Brien, P.D.; Mendelson, F.E.; Hayes, J.M.; Feldman, E.L. Sex differences in insulin resistance, but not peripheral neuropathy, in a diet-induced prediabetes mouse model. Dis. Model. Mech. 2021, 14, dmm048909. [Google Scholar] [CrossRef]
- Majumder, K.; Chakrabarti, S.; Morton, J.S.; Panahi, S.; Kaufman, S.; Davidge, S.T.; Wu, J. Egg-derived tri-peptide IRW exerts antihypertensive effects in spontaneously hypertensive rats. PLoS ONE 2013, 8, e82829. [Google Scholar] [CrossRef] [Green Version]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L. High-carbohydrate, high-fat diet–induced metabolic syndrome and cardiovascular remodeling in rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624. [Google Scholar] [CrossRef]
- Majumder, K.; Chakrabarti, S.; Davidge, S.T.; Wu, J. Structure and activity study of egg protein ovotransferrin derived peptides (IRW and IQW) on endothelial inflammatory response and oxidative stress. J. Agric. Food Chem. 2013, 61, 2120–2129. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Chakrabarti, S.; Davidge, S.T.; Wu, J. Modulatory effects of egg white ovotransferrin-derived tripeptide IRW (Ile-Arg-Trp) on vascular smooth muscle cells against angiotensin II stimulation. J. Agric. Food Chem. 2016, 64, 7342–7347. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zou, M.-H. Molecular insights and therapeutic targets for diabetic endothelial dysfunction. Circulation 2009, 120, 1266–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruderman, N.B.; Xu, X.J.; Nelson, L.; Cacicedo, J.M.; Saha, A.K.; Lan, F.; Ido, Y. AMPK and SIRT1: A long-standing partnership? Am. J. Physiol. Endocrinol. 2010, 298, 751–760. [Google Scholar] [CrossRef]
- Haye, A.; Ansari, M.A.; Rahman, S.O.; Shamsi, Y.; Ahmed, D.; Sharma, M. Role of AMP-activated protein kinase on cardio-metabolic abnormalities in the development of diabetic cardiomyopathy: A molecular landscape. Eur. J. Pharmacol. 2020, 888, 173376. [Google Scholar] [CrossRef]
- Zhang, F.; Ren, X.; Zhao, M.; Zhou, B.; Han, Y. Angiotensin-(1–7) abrogates angiotensin II-induced proliferation, migration and inflammation in VSMCs through inactivation of ROS-mediated PI3K/Akt and MAPK/ERK signaling pathways. Sci. Rep. 2016, 6, 34621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouallegue, A.; Bou Daou, G.; Srivastava, A.K. Endothelin-1-induced signaling pathways in vascular smooth muscle cells. Curr. Vasc. Pharmacol. 2007, 5, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.D.; Oh, H.; Kim, M.-S. Higher intakes of fruits, vegetables, and multiple individual nutrients is associated with a lower risk of metabolic syndrome among adults with comorbidities. Nutr. Res. 2022, 99, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yarizadeh, H.; Setayesh, L.; Majidi, N.; Rasaei, N.; Mehranfar, S.; Ebrahimi, R.; Casazzza, K.; Mirzaei, K. Nutrient patterns and their relation to obesity and metabolic syndrome in Iranian overweight and obese adult women. Eat Weight Disord. 2022, 27, 1327–1337. [Google Scholar] [CrossRef]
- Prasad, H.; Ryan, D.A.; Celzo, M.F.; Stapleton, D. Metabolic syndrome: Definition and therapeutic implications. Postgrad. Med. 2012, 124, 21–30. [Google Scholar] [CrossRef]
- Frigolet, M.E.; Torres, N.; Tovar, A.R. The renin–angiotensin system in adipose tissue and its metabolic consequences during obesity. J. Nutr. Biochem. 2013, 24, 2003–2015. [Google Scholar] [CrossRef]
- Skov, J.; Persson, F.; Frøkiær, J.; Christiansen, J.S. Tissue renin–angiotensin systems: A unifying hypothesis of metabolic disease. Front. Endocrinol. 2014, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.-C.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-converting enzyme 2: SARS-CoV-2 receptor and regulator of the renin-angiotensin system: Celebrating the 20th anniversary of the discovery of ACE2. Circ. Res. 2020, 126, 1456–1474. [Google Scholar] [CrossRef]
- Li, L.; Yi-Ming, W.; Li, Z.-Z.; Zhao, L.; Yu, Y.-S.; Li, D.-J.; Xia, C.-Y.; Liu, J.-G.; Su, D.-F. Local RAS and inflammatory factors are involved in cardiovascular hypertrophy in spontaneously hypertensive rats. Pharmacol. Res. 2008, 58, 196–201. [Google Scholar] [CrossRef]
- Mackenzie, R.W.; Elliott, B.T. Akt/PKB activation and insulin signaling: A novel insulin signaling pathway in the treatment of type 2 diabetes. Diabetes Metab Syndr Obes. 2014, 7, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Ruderman, N.B.; Carling, D.; Prentki, M.; Cacicedo, J.M. AMPK, insulin resistance, and the metabolic syndrome. J. Clin. Investig. 2013, 123, 2764–2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbasnezhad, A.; Niazmand, S.; Mahmoudabady, M.; Rezaee, S.A.; Soukhtanloo, M.; Mosallanejad, R.; Hayatdavoudi, P. Nigella sativa L. seed regulated eNOS, VCAM-1 and LOX-1 genes expression and improved vasoreactivity in aorta of diabetic rat. J. Ethnopharmacol. 2019, 228, 142–147. [Google Scholar] [CrossRef] [PubMed]
- García-Prieto, C.F.; Hernández-Nuño, F.; Rio, D.D.; Ruiz-Hurtado, G.; Aránguez, I.; Ruiz-Gayo, M.; Somoza, B.; Fernández-Alfonso, M.S. High-fat diet induces endothelial dysfunction through a down-regulation of the endothelial AMPK–PI3K–Akt–eNOS pathway. Mol. Nutr. Food Res. 2015, 59, 520–532. [Google Scholar] [CrossRef]
- Ewart, M.-A.; Kennedy, S. AMPK and vasculoprotection. Pharmacol. Ther. 2011, 131, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Machha, A.; Schechter, A.N. Inorganic nitrate: A major player in the cardiovascular health benefits of vegetables? Nutr. Rev. 2012, 70, 367–372. [Google Scholar] [CrossRef]
- Triggle, C.R.; Ding, H. A review of endothelial dysfunction in diabetes: A focus on the contribution of a dysfunctional eNOS. J. Am. Soc. Hypertens. 2010, 4, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Harris, R.C. Role of endothelial nitric oxide synthase in diabetic nephropathy: Lessons from diabetic eNOS knockout mice. J. Diabetes Res. 2014, 2014, 590541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balakumar, P.; Kathuria, S. Submaximal PPARγ activation and endothelial dysfunction: New perspectives for the management of cardiovascular disorders. Br. J. Pharmacol. 2012, 166, 1981–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.-Z.; Cheng, Y.-W.; Jin, H.-Y.; Chang, Q.; Shang, Q.-H.; Xu, Y.-L.; Chen, L.-X.; Xu, R.; Song, B.; Zhong, J.-C. The sirtuin 6 prevents angiotensin II-mediated myocardial fibrosis and injury by targeting AMPK-ACE2 signaling. Oncotarget 2017, 8, 72302. [Google Scholar] [CrossRef] [Green Version]
- Ota, H.; Akishita, M.; Eto, M.; Iijima, K.; Kaneki, M.; Ouchi, Y. Sirt1 modulates premature senescence-like phenotype in human endothelial cells. J. Mol. Cell. Cardiol. 2007, 43, 571–579. [Google Scholar] [CrossRef]
- Mattagajasingh, I.; Kim, C.-S.; Naqvi, A.; Yamamori, T.; Hoffman, T.A.; Jung, S.-B.; DeRicco, J.; Kasuno, K.; Irani, K. SIRT1 promotes endothelium-dependent vascular relaxation by activating endothelial nitric oxide synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 14855–14860. [Google Scholar] [CrossRef] [Green Version]
- Karpe, P.A.; Tikoo, K. Heat shock prevents insulin resistance–induced vascular complications by augmenting angiotensin-(1-7) signaling. Diabetes 2014, 63, 1124–1139. [Google Scholar] [CrossRef] [Green Version]
- Jansson, P.A. Endothelial dysfunction in insulin resistance and type 2 diabetes. J. Intern. Med. 2007, 262, 173–183. [Google Scholar] [CrossRef]
- Ye, Y.; Zhong, X.; Li, N.; Pan, T. Protective effects of liraglutide on glomerular podocytes in obese mice by inhibiting the inflammatory factor TNF-α-mediated NF-κB and MAPK pathway. Obes. Res. Clin. Pract. 2019, 13, 385–390. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashkar, F.; Bhullar, K.S.; Jiang, X.; Wu, J. Tripeptide IRW Improves AMPK/eNOS Signaling Pathway via Activating ACE2 in the Aorta of High-Fat-Diet-Fed C57BL/6 Mice. Biology 2023, 12, 556. https://doi.org/10.3390/biology12040556
Ashkar F, Bhullar KS, Jiang X, Wu J. Tripeptide IRW Improves AMPK/eNOS Signaling Pathway via Activating ACE2 in the Aorta of High-Fat-Diet-Fed C57BL/6 Mice. Biology. 2023; 12(4):556. https://doi.org/10.3390/biology12040556
Chicago/Turabian StyleAshkar, Fatemeh, Khushwant S. Bhullar, Xu Jiang, and Jianping Wu. 2023. "Tripeptide IRW Improves AMPK/eNOS Signaling Pathway via Activating ACE2 in the Aorta of High-Fat-Diet-Fed C57BL/6 Mice" Biology 12, no. 4: 556. https://doi.org/10.3390/biology12040556
APA StyleAshkar, F., Bhullar, K. S., Jiang, X., & Wu, J. (2023). Tripeptide IRW Improves AMPK/eNOS Signaling Pathway via Activating ACE2 in the Aorta of High-Fat-Diet-Fed C57BL/6 Mice. Biology, 12(4), 556. https://doi.org/10.3390/biology12040556