
Factors Limiting Radial Growth of Conifers on Their Semiarid Borders across Kazakhstan
 , , , and
, , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
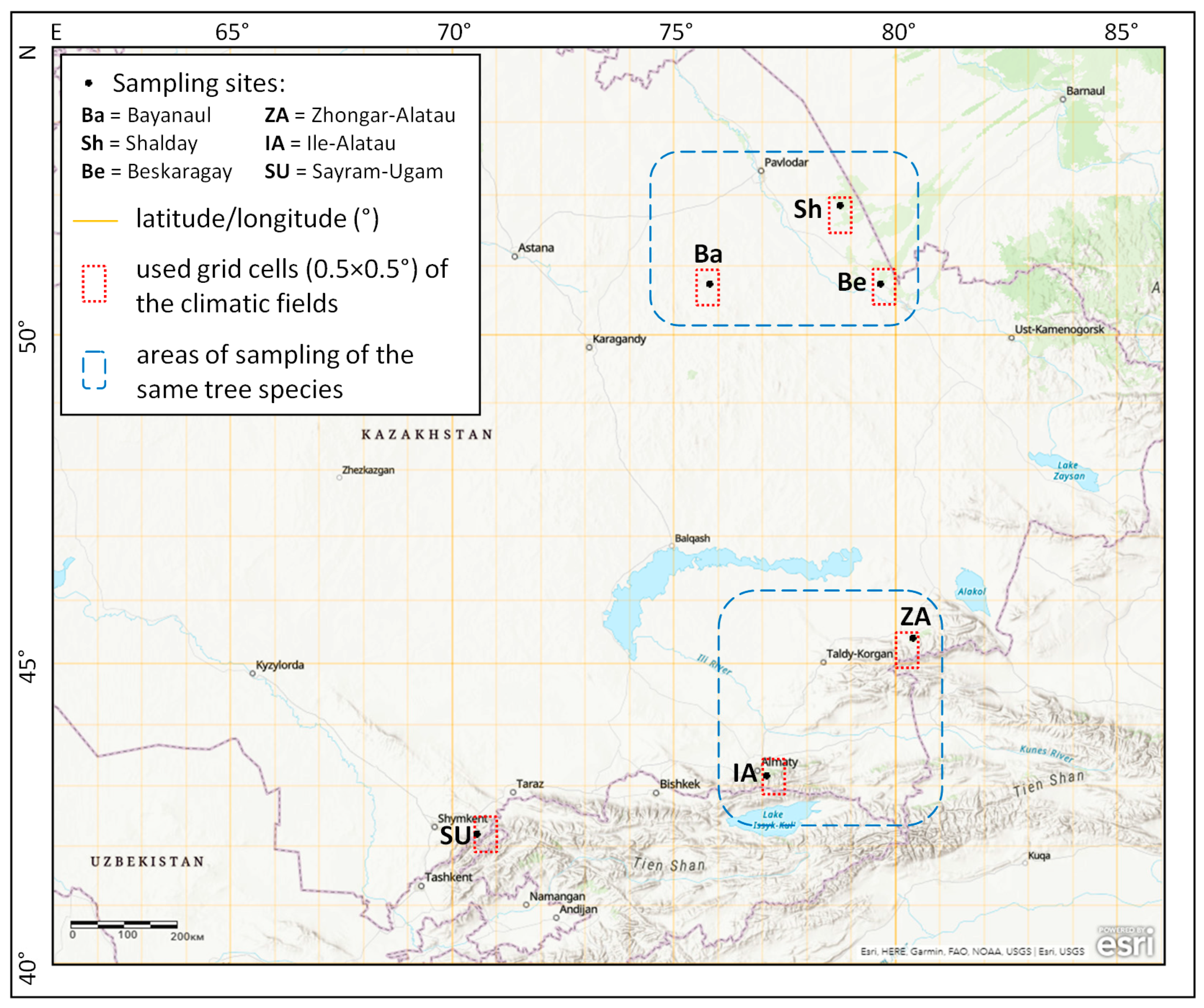
2.1. Study Region and Sampling Sites
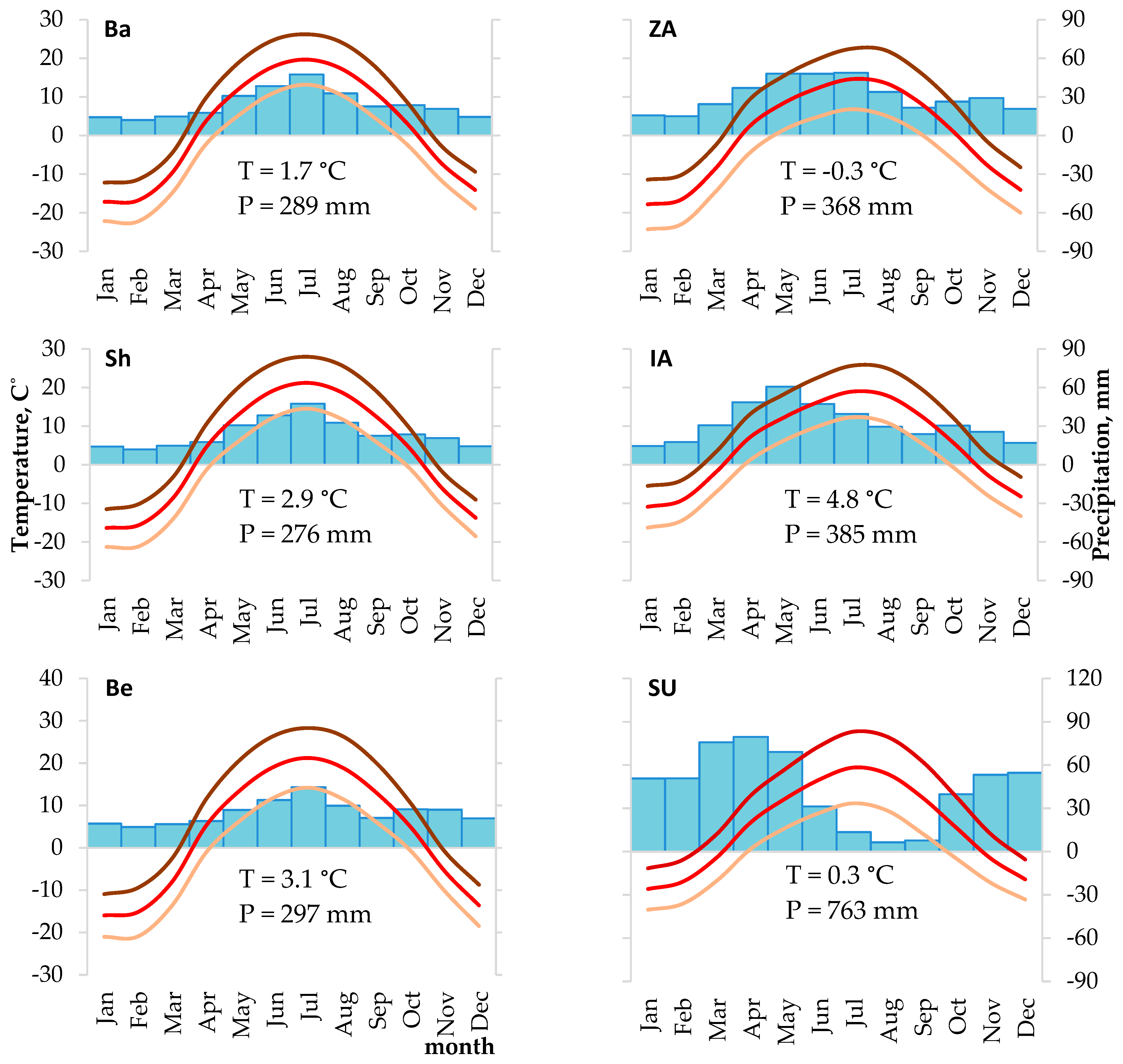
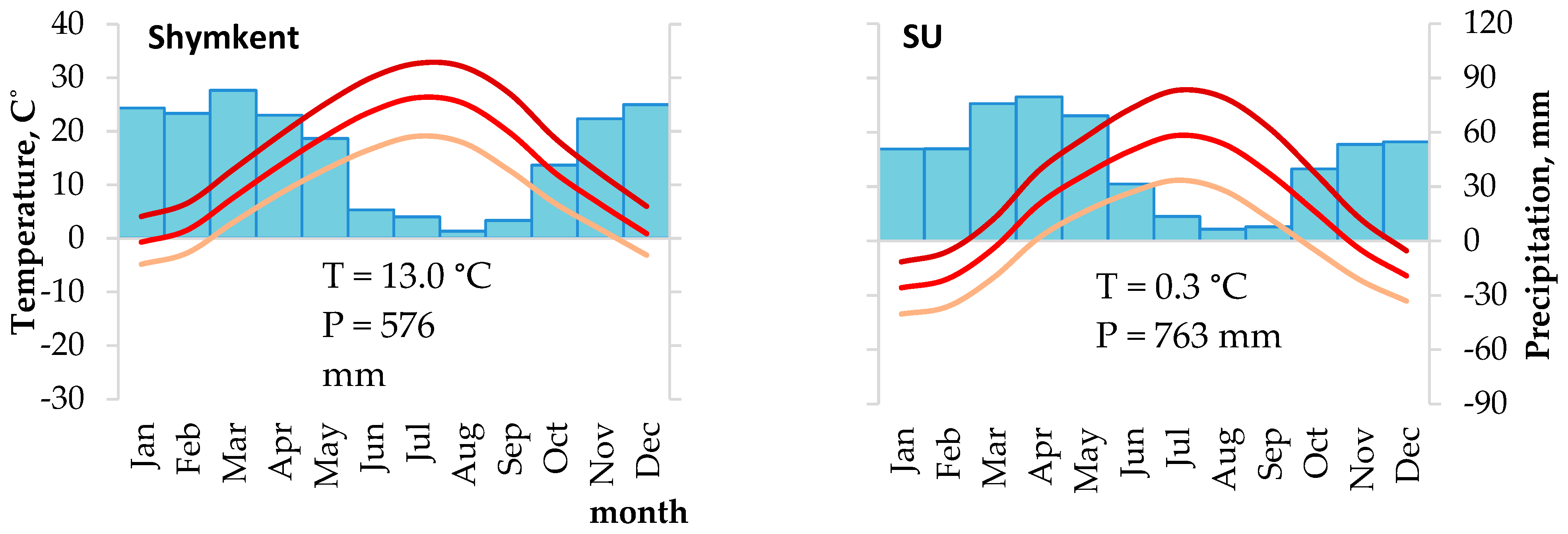
2.2. Data and Their Analysis
3. Results
3.1. Developed Chronologies and Their Statistic Characteristics
3.2. Climatic Response of Conifer Radial Growth
4. Discussion
4.1. Spatial Patterns in the Conifer Radial Growth
4.2. Conifer Growth Limitation by Climatic Factors in the Semiarid Territories of Kazakhstan
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Breshears, D.D.; Whicker, J.J.; Johansen, M.P.; Pinder, J.E. Wind and water erosion and transport in semi-arid shrubland, grassland and forest ecosystems: Quantifying dominance of horizontal wind-driven transport. Earth Surf. Process. Landf. 2003, 28, 1189–1209. [Google Scholar] [CrossRef]
- FAO. The State of the World’s Forests 2018—Forest Pathways to Sustainable Development; Food and Agriculture Organization: Rome, Italy, 2018. [Google Scholar]
- Jhariya, M.K.; Banerjee, A.; Meena, R.S.; Yadav, D.K. Sustainable Agriculture, Forest and Environmental Management; Springer: Singapore, 2019. [Google Scholar] [CrossRef]
- Verkerk, P.J.; Costanza, R.; Hetemäki, L.; Kubiszewski, I.; Leskinen, P.; Nabuurs, G.J.; Potocnik, J.; Palahi, M. Climate-smart forestry: The missing link. For. Policy Econ. 2020, 115, 102164. [Google Scholar] [CrossRef]
- Carrer, M.; Urbinati, C. Age-dependent tree-ring growth responses to climate in Larix decidua and Pinus cembra. Ecology 2004, 85, 730–740. [Google Scholar] [CrossRef]
- Chen, P.Y.; Welsh, C.; Hamann, A. Geographic variation in growth response of Douglas-fir to interannual climate variability and projected climate change. Glob. Chang. Biol. 2010, 16, 3374–3385. [Google Scholar] [CrossRef]
- Way, D.A.; Oren, R.A.M. Differential responses to changes in growth temperature between trees from different functional groups and biomes: A review and synthesis of data. Tree Physiol. 2010, 30, 669–688. [Google Scholar] [CrossRef] [PubMed]
- Babushkina, E.A.; Zhirnova, D.F.; Belokopytova, L.V.; Tychkov, I.I.; Vaganov, E.A.; Krutovsky, K.V. Response of four tree species to changing climate in a moisture-limited area of South Siberia. Forests 2019, 10, 999. [Google Scholar] [CrossRef]
- Menezes-Silva, P.E.; Loram-Lourenço, L.; Alves, R.D.F.B.; Sousa, L.F.; Almeida, S.E.D.S.; Farnese, F.S. Different ways to die in a changing world: Consequences of climate change for tree species performance and survival through an ecophysiological perspective. Ecol. Evol. 2019, 9, 11979–11999. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK, 2022; 3056p. [Google Scholar]
- Zhang, Q.; Ye, J.F.; Le, C.T.; Njenga, D.M.; Rabarijaona, N.R.; Omollo, W.O.; Lu, L.M.; Liu, B.; Chen, Z.D. New insights into the formation of biodiversity hotspots of the Kenyan flora. Divers. Distrib. 2022, 28, 2696–2711. [Google Scholar] [CrossRef]
- Hernandez, J.O.; Naeem, M.; Zaman, W. How does changing environment influence plant seed movements as populations of dispersal vectors decline? Plants 2023, 12, 1462. [Google Scholar] [CrossRef]
- Kirschbaum, M.U. Forest growth and species distribution in a changing climate. Tree Physiol. 2000, 20, 309–322. [Google Scholar] [CrossRef]
- Chazdon, R.L. Beyond deforestation: Restoring forests and ecosystem services on degraded lands. Science 2008, 320, 1458–1460. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Liu, H.; Park Williams, A.; Allen, C.D.; Guo, D.; Wu, X.; Anenkhonov, O.A.; Liang, E.; Sandanov, D.V.; Yin, Y.; Qi, Z.; et al. Rapid warming accelerates tree growth decline in semi-arid forests of Inner Asia. Glob. Chang. Biol. 2013, 19, 2500–2510. [Google Scholar] [CrossRef]
- McDowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.; Aukema, B.H.; Bond-Lamberty, B.; Chini, L.; Clark, J.S.; Dietze, M.; Grossiord, C.; Hanbury-Brown, A.; et al. Pervasive shifts in forest dynamics in a changing world. Science 2020, 368, eaaz9463. [Google Scholar] [CrossRef]
- Ge, W.; Deng, L.; Wang, F.; Han, J. Quantifying the contributions of human activities and climate change to vegetation net primary productivity dynamics in China from 2001 to 2016. Sci. Total Environ. 2021, 773, 145648. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Lioubimtseva, E.; Cole, R. Uncertainties of climate change in arid environments of Central Asia. Rev. Fish. Sci. 2006, 14, 29–49. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, C.; Hu, Q.; Tian, H. Temperature changes in Central Asia from 1979 to 2011 based on multiple datasets. J. Clim. 2014, 27, 1143–1167. [Google Scholar] [CrossRef]
- Seddon, A.W.; Macias-Fauria, M.; Long, P.R.; Benz, D.; Willis, K.J. Sensitivity of global terrestrial ecosystems to climate variability. Nature 2016, 531, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Liu, T.; Meng, F.; Duan, Y.; Bao, A.; Frankl, A.; De Maeyer, P. Spatiotemporal characteristics of future changes in precipitation and temperature in Central Asia. Int. J. Climatol. 2019, 39, 1571–1588. [Google Scholar] [CrossRef]
- Zhu, X.; Wei, Z.; Dong, W.; Ji, Z.; Wen, X.; Zheng, Z.; Yan, D.; Chen, D. Dynamical downscaling simulation and projection for mean and extreme temperature and precipitation over central Asia. Clim. Dyn. 2020, 54, 3279–3306. [Google Scholar] [CrossRef]
- Lioubimtseva, E.; Cole, R.; Adams, J.M.; Kapustin, G. Impacts of climate and land-cover changes in arid lands of Central Asia. J. Arid Environ. 2005, 62, 285–308. [Google Scholar] [CrossRef]
- Lioubimtseva, E.; Henebry, G.M. Climate and environmental change in arid Central Asia: Impacts, vulnerability, and adaptations. J. Arid Environ. 2009, 73, 963–977. [Google Scholar] [CrossRef]
- Siegfried, T.; Bernauer, T.; Guiennet, R.; Sellars, S.; Robertson, A.W.; Mankin, J.; Bauer-Gottwein, P.; Yakovlev, A. Will climate change exacerbate water stress in Central Asia? Clim. Chang. 2012, 112, 881–899. [Google Scholar] [CrossRef]
- Reyer, C.P.; Otto, I.M.; Adams, S.; Albrecht, T.; Baarsch, F.; Cartsburg, M.; Coumou, D.; Eden, A.; Ludi, E.; Marcus, R.; et al. Climate change impacts in Central Asia and their implications for development. Reg. Environ. Chang. 2017, 17, 1639–1650. [Google Scholar] [CrossRef]
- He, H.; Hamdi, R.; Cai, P.; Luo, G.; Ochege, F.U.; Zhang, M.; Termonia, P.; De Maeyer, P.; Li, C. Impacts of historical land use/cover change (1980–2015) on summer climate in the Aral Sea region. J. Geophys. Res. Atmos. 2021, 126, e2020JD032638. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Y.; Li, W.; Deng, H.; Fang, G. Potential impacts of climate change on vegetation dynamics in Central Asia. J. Geophys. Res. Atmos. 2015, 120, 12345–12356. [Google Scholar] [CrossRef]
- Yin, G.; Hu, Z.; Chen, X.; Tiyip, T. Vegetation dynamics and its response to climate change in Central Asia. J. Arid Land 2016, 8, 375–388. [Google Scholar] [CrossRef]
- Jiang, L.; Bao, A.; Guo, H.; Ndayisaba, F. Vegetation dynamics and responses to climate change and human activities in Central Asia. Sci. Total Environ. 2017, 599, 967–980. [Google Scholar] [CrossRef]
- Li, W.; Tojibaev, K.S.; Hisoriev, H.; Shomurodov, K.F.; Luo, M.; Feng, Y.; Ma, K. Mapping Asia plants: Current status of floristic information for Central Asian flora. Glob. Ecol. Conserv. 2020, 24, e01220. [Google Scholar] [CrossRef]
- Foggin, J.M.; Lechner, A.M.; Emslie-Smith, M.; Hughes, A.C.; Sternberg, T.; Dossani, R. Belt and road initiative in Central Asia: Anticipating socioecological challenges from large-scale infrastructure in a global biodiversity hotspot. Conserv. Lett. 2021, 14, e12819:1–e12819:15. [Google Scholar] [CrossRef]
- Kushlin, A.V.; Schillhorn-van-Veen, T.W.; Sutton, W. Kazakhstan: Forest Sector in Transition: The Resource, the Users and Sustainable Use; World Bank: Washington, DC, USA, 2004; 66p. [Google Scholar]
- Zubairov, B.; Lentschke, J.; Schröder, H. Dendroclimatology in Kazakhstan. Dendrochronologia 2019, 56, 125602. [Google Scholar] [CrossRef]
- Kirillov, V.; Pathak, A.; Kabanova, S.; Stikhareva, T.; Turumbayev, S.; Savazova, D.; Kerteshev, T.; Rakhimzhanov, A. Human resources for forestry in Kazakhstan: Current status, potential and problems. Folia For. Pol. 2022, 64, 226–244. [Google Scholar] [CrossRef]
- Meshkov, V.V.; Baizakov, S.B.; Yeger, A.V.; Orozumbekov, A. Forest rehabilitation in Kazakhstan. In Keep Asia Green. Volume IV “West and Central Asia”; Lee, D.K., Kleine, M., Eds.; IUFRO: Vienna, Austria, 2009; pp. 83–130. [Google Scholar]
- Yesserkepova, I. Kazakhstan. In Forests and Climate Change in Eastern Europe and Central Asia; Csaba, M., Ed.; FAO: Rome, Italy, 2010; pp. 63–75. [Google Scholar]
- Sehring, J. Forests in the Context of Climate Change in Kazakhstan; Justus Liebig University Giessen, Center for international Development and Environmental Research (ZEU): Giessen, Germany, 2012. [Google Scholar] [CrossRef]
- Andersson, L.; Ardfors, E. Evaluating Options for Implementing the Kazakhstan Forest Restoration Targets. Master’s Thesis, Chalmers University of Technology, Gothenburg, Sweden, 2021. [Google Scholar]
- Rudel, T.K.; Meyfroidt, P.; Chazdon, R.; Bongers, F.; Sloan, S.; Grau, H.R.; Van Holt, T.; Schneider, L. Whither the forest transition? Climate change, policy responses, and redistributed forests in the twenty-first century. Ambio 2020, 49, 74–84. [Google Scholar] [CrossRef]
- Cook, E.R.; Cole, J. On predicting the response of forests in eastern North America to future climatic change. Clim. Chang. 1991, 19, 271–282. [Google Scholar] [CrossRef]
- D’Arrigo, R.D.; Jacoby, G.C. Tree growth-climate relationships at the northern boreal forests tree line of North America: Evaluation of potential response to increasing carbon dioxide. Glob. Biochem. Cycles 1993, 7, 525–535. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Christopoulou, A.; Galanidis, A.; Michelaki, C.Z.; Dimitrakopoulos, P.G.; Fulé, P.Z.; Arianoutsou, M. Tree growth-climate relationships in a forest-plot network on Mediterranean mountains. Sci. Total Environ. 2017, 598, 393–403. [Google Scholar] [CrossRef]
- De Micco, V.; Carrer, M.; Rathgeber, C.B.; Camarero, J.J.; Voltas, J.; Cherubini, P.; Battipaglia, G. From xylogenesis to tree rings: Wood traits to investigate tree response to environmental changes. IAWA J. 2019, 40, 155–182. [Google Scholar] [CrossRef]
- Upadhyay, K.K.; Tripathi, S.K. Sustainable forest management under climate change: A dendrochronological approach. Environ. Ecol. 2019, 37, 998–1006. [Google Scholar]
- Farooq, I.; Shah, A.R.; Sahana, M.; Ehsan, M.A. Assessment of drought conditions over different climate zones of Kazakhstan using standardised precipitation evapotranspiration index. Earth Syst. Environ. 2023, 7, 283–296. [Google Scholar] [CrossRef]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Sciences; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990. [Google Scholar]
- Rinn, F. TSAP V3.5: Computer Program for Tree-Ring Analysis and Presentation; Frank Rinn Distribution: Heibelberg, Germany, 1996. [Google Scholar]
- Holmes, R.L. The dendrochronology program library. In International Tree-Ring Data Bank Program Library User’s Manual; Grissino-Mayer, H.D., Ed.; Laboratory of Tree-Ring Research, University of Arizona: Tucson, AZ, USA, 1992; pp. 40–74. [Google Scholar]
- Cook, E.R.; Krusic, P.J. A Tree-Ring Standardization Program Based on Detrending and Autoregressive Time Series Modeling, with Interactive Graphics (ARSTAN). Available online: http://www.ldeo.Columbia.edu/res/fac/trl/public/publicSoftware.html (accessed on 10 March 2023).
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Appl. Meteorol. Clim. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Fritts, H.C.; Smith, D.G.; Cardis, J.W.; Budelsky, C.A. Tree-ring characteristics along a vegetation gradient in northern Arizona. Ecology 1965, 46, 393–401. [Google Scholar] [CrossRef]
- Fritts, H.C. Growth-rings of trees: Their correlation with climate: Patterns of ring widths in trees in semiarid sites depend on climate-controlled physiological factors. Science 1966, 154, 973–979. [Google Scholar] [CrossRef]
- Benito-Garzon, M.; Alia, R.; Robson, T.M.; Zavala, M.A. Intra-specific variability and plasticity influence potential tree species distributions under climate change. Glob. Ecol. Biogeogr. 2011, 20, 766–778. [Google Scholar] [CrossRef]
- Santini, F.; Sin, E.; Shestakova, T.A.; Hereş, A.; Voltas, J. Phenotypic plasticity and intra-specific variation in water-use efficiency of Pinus sylvestris L. inferred from tree rings. In In Proceedings of the Tree Rings in Archaeology, Climatology and Ecology (TRACE 2017), Svetlogorsk, Russia, 16–21 May 2017. [Google Scholar] [CrossRef]
- Santini, F.; Ferrio, J.P.; Hereş, A.M.; Notivol, E.; Piqué, M.; Serrano, L.; Shestakova, T.A.; Sin, E.; Vericat, P.; Voltas, J. Scarce population genetic differentiation but substantial spatiotemporal phenotypic variation of water-use efficiency in Pinus sylvestris at its western distribution range. Eur. J. For. Res. 2018, 137, 863–878. [Google Scholar] [CrossRef]
- Pazouki, L.; Shanjani, P.S.; Fields, P.D.; Martins, K.; Suhhorutšenko, M.; Viinalass, H.; Niinemets, Ü. Large within-population genetic diversity of the widespread conifer Pinus sylvestris at its soil fertility limit characterized by nuclear and chloroplast microsatellite markers. Eur. J. For. Res. 2016, 135, 161–177. [Google Scholar] [CrossRef]
- Benito-Garzon, M.; Vizcaino-Palomar, N. Biogeographical patterns of local adaptation and plasticity of Mediterranean pines and their implications under climate change. In Pines and Their Mixed Forest Ecosystems in the Mediterranean Basin; Ne’eman, G., Osem, Y., Eds.; Springer: Cham, Germany, 2021; pp. 71–82. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D. An updated list of species used in tree-ring research. Tree-Ring Bull. 1993, 53, 17–43. [Google Scholar]
- Ali, G. Climate change and associated spatial heterogeneity of Pakistan: Empirical evidence using multidisciplinary approach. Sci. Total Environ. 2018, 634, 95–108. [Google Scholar] [CrossRef]
- Ljungqvist, F.C.; Piermattei, A.; Seim, A.; Krusic, P.J.; Büntgen, U.; He, M.; Kirdyanov, A.V.; Luterbacher, J.; Schneider, L.; Seftigen, K.; et al. Ranking of tree-ring based hydroclimate reconstructions of the past millennium. Quat. Sci. Rev. 2020, 230, 106074. [Google Scholar] [CrossRef]
- Fang, O.; Zhang, Q.B. Tree resilience to drought increases in the Tibetan Plateau. Glob. Chang. Biol. 2019, 25, 245–253. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Y.; Xiao, J.; Lyu, L. Species-specific coupling of tree-ring width and litter production in a temperate mixed forest. For. Ecol. Manag. 2022, 504, 119831. [Google Scholar] [CrossRef]
- Dorado-Linan, I.; Piovesan, G.; Martinez-Sancho, E.; Gea-Izquierdo, G.; Zang, C.; Canellas, I.; Castagneri, D.; Di Filippo, A.; Gutierrez, E.; Ewald, J.; et al. Geographical adaptation prevails over species-specific determinism in trees’ vulnerability to climate change at Mediterranean rear-edge forests. Glob. Chang. Biol. 2019, 25, 1296–1314. [Google Scholar] [CrossRef]
- Grießinger, J.; Bräuning, A.; Helle, G.; Schleser, G.H.; Hochreuther, P.; Meier, W.J.H.; Zhu, H. A dual stable isotope approach unravels common climate signals and species-specific responses to environmental change stored in multi-century tree-ring series from the Tibetan plateau. Geosciences 2019, 9, 151. [Google Scholar] [CrossRef]
- Tumajer, J.; Kaspar, J.; Kuzelova, H.; Shishov, V.V.; Tychkov, I.I.; Popkova, M.I.; Vaganov, E.A.; Treml, V. Forward modeling reveals multidecadal trends in cambial kinetics and phenology at treeline. Front. Plant Sci. 2021, 12, 613643. [Google Scholar] [CrossRef]
- Zhirnova, D.F.; Belokopytova, L.V.; Meko, D.M.; Babushkina, E.A.; Vaganov, E.A. Climate change and tree growth in the Khakass-Minusinsk Depression (South Siberia) impacted by large water reservoirs. Sci. Rep. 2021, 11, 14266. [Google Scholar] [CrossRef]
- Arzac, A.; de Quijano, D.D.; Khotcinskaia, K.I.; Tychkov, I.I.; Voronin, V.I.; Kirdyanov, A.V. The buffering effect of the Lake Baikal on climate impact on Pinus sylvestris L. radial growth. Agric. For. Meteorol. 2022, 313, 108764. [Google Scholar] [CrossRef]
- Brandes, A.F.D.N.; Albuquerque, R.P.; Moraes, L.F.D.D.; Barros, C.F. Annual tree rings in Piptadenia gonoacantha (Mart.) JF Macbr. in a restoration experiment in the Atlantic Forest: Potential for dendroecological research. Acta Bot. Bras. 2016, 30, 383–388. [Google Scholar] [CrossRef]
- Belokopytova, L.V.; Zhirnova, D.F.; Meko, D.M.; Babushkina, E.A.; Vaganov, E.A.; Krutovsky, K.V. Tree rings reveal the impact of soil temperature on larch growth in the forest-steppe of Siberia. Forests 2021, 12, 1765. [Google Scholar] [CrossRef]
- Carnicer, J.; Vives-Ingla, M.; Blanquer, L.; Mendez-Camps, X.; Rosell, C.; Sabate, S.; Gutierrez, E.; Sauras, T.; Penuelas, J.; Barbeta, A. Forest resilience to global warming is strongly modulated by local-scale topographic, microclimatic and biotic conditions. J. Ecol. 2021, 109, 3322–3339. [Google Scholar] [CrossRef]
- Agafonov, L.I.; Gurskaya, M.A.; Kukarskih, V.V.; Bubnov, M.O.; Devi, N.M.; Galimova, A.A. Insular pine forests of the Southern Urals and ribbon pine forests of the Altai as objects of dendroclimatic research. Russ. J. Ecol. 2021, 52, 349–357. [Google Scholar] [CrossRef]
- Demina, A.V.; Belokopytova, L.V.; Zhirnova, D.F.; Mehrotra, N.; Shah, S.K.; Babushkina, E.A.; Vaganov, E.A. Degree of connectivity in reconstructed precipitation dynamics and extremes for semiarid regions across South Siberia. Dendrochronologia 2022, 71, 125903. [Google Scholar] [CrossRef]
- Mukhamedshin, K.D.; Sartbaev, S.K. Archa—Tree of Life; Kaynar: Alma-Ata, Kazakhstan, 1981. (In Russian) [Google Scholar]
- Kopabayeva, A.; Mazarzhanova, K.; Kose, N.; Akkemik, U. Tree-ring chronologies of Pinus sylvestris from Burabai Region (Kazakhstan) and their response to climate change. Dendrobiology 2017, 78, 96–110. [Google Scholar] [CrossRef]
- Zubairov, B.; Heußner, K.-U.; Schröder, H. Searching for the best correlation between climate and tree rings in the Trans-Ili Alatau, Kazakhstan. Dendrobiology 2018, 79, 119–130. [Google Scholar] [CrossRef]
- Wang, T.; Ren, H.; Ma, K. Climatic signals in tree ring of Picea schrenkiana along an altitudinal gradient in the central Tianshan Mountains, northwestern China. Trees 2005, 19, 736–742. [Google Scholar] [CrossRef]
- Wang, H.Q.; Chen, F.; Ermenbaev, B.; Satylkanov, R. Comparison of drought-sensitive tree-ring records from the Tien Shan of Kyrgyzstan and Xinjiang (China) during the last six centuries. Adv. Clim. Chang. Res. 2017, 8, 18–25. [Google Scholar] [CrossRef]
- Li, J.; Gou, X.; Cook, E.R.; Chen, F. Tree-ring based drought reconstruction for the central Tien Shan area in northwest China. Geophys. Res. Lett. 2006, 33, L07715. [Google Scholar] [CrossRef]
- Chen, F.; Yuan, Y.; Davi, N.; Zhang, T. Upper Irtysh River flow since AD 1500 as reconstructed by tree rings, reveals the hydroclimatic signal of inner Asia. Clim. Chang. 2016, 139, 651–665. [Google Scholar] [CrossRef]
- Seim, A.; Tulyaganov, T.; Omurova, G.; Nikolyai, L.; Botman, E.; Linderholm, H.W. Dendroclimatological potential of three juniper species from the Turkestan range, northwestern Pamir-Alay Mountains, Uzbekistan. Trees 2016, 30, 733–748. [Google Scholar] [CrossRef]
- Huo, Y.; Gou, X.; Liu, W.; Li, J.; Zhang, F.; Fang, K. Climate-growth relationships of Schrenk spruce (Picea schrenkiana) along an altitudinal gradient in the western Tianshan mountains, northwest China. Trees 2017, 31, 429–439. [Google Scholar] [CrossRef]
- Peng, J.; Li, J.; Li, X.; Cui, J.; Peng, M. Climate-growth relationships of Chinese pine (Pinus tabulaeformis Carr.) at Mt. Shiren in climatic transition zone, central China. Biology 2022, 11, 753. [Google Scholar] [CrossRef]
- Crous, K.Y.; Uddling, J.; De Kauwe, M.G. Temperature responses of photosynthesis and respiration in evergreen trees from boreal to tropical latitudes. New Phytol. 2022, 234, 353–374. [Google Scholar] [CrossRef]
- Al Farsi, K.; Cameron, R.W.; Hitchmough, J.; Lupton, D. Assessing the effects of drought and temperature on the establishment of Juniperus seravschanica saplings in Northern Oman. Sibbaldia J. Bot. Gard. Hortic. 2016, 14, 37–53. [Google Scholar] [CrossRef]
- Rahimian Boogar, A.; Salehi, H.; Seyedabadi, E. Distribution and physiology of Juniperus seravschanica trees in the Genow—the southernmost and arid habitat of Iran. Water 2022, 14, 3508. [Google Scholar] [CrossRef]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef]
- MacLaren, C.A. Climate change drives decline of Juniperus seravschanica in Oman. J. Arid Environ. 2016, 128, 91–100. [Google Scholar] [CrossRef]
- Breshears, D.D.; Fontaine, J.B.; Ruthrof, K.X.; Field, J.P.; Feng, X.; Burger, J.R.; Law, D.J.; Kala, J.; Hardy, G.E.S.J. Underappreciated plant vulnerabilities to heat waves. New Phytol. 2021, 231, 32–39. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef]
- Schaberg, P.G.; Hennon, P.E.; D’amore, D.V.; Hawley, G.J. Influence of simulated snow cover on the cold tolerance and freezing injury of yellow-cedar seedlings. Glob. Chang. Biol. 2008, 14, 1282–1293. [Google Scholar] [CrossRef]
- Reinmann, A.B.; Susser, J.R.; Demaria, E.M.; Templer, P.H. Declines in northern forest tree growth following snowpack decline and soil freezing. Glob. Chang. Biol. 2019, 25, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yuan, Y.; Gou, X.; Zhang, T.; Zou, C.; Ji, C.; Fan, Z.; Qin, L.; Shang, H.; Li, X. Intra-annual radial growth of Schrenk spruce (Picea schrenkiana Fisch. et Mey) and its response to climate on the northern slopes of the Tianshan Mountains. Dendrochronologia 2016, 40, 36–42. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Manzanedo, R.D.; Zhang, F.; Pederson, N. Cambial phenology and xylogenesis of Juniperus przewalskii over a climatic gradient is influenced by both temperature and drought. Agric. For. Meteorol. 2018, 260, 165–175. [Google Scholar] [CrossRef]
- Deslauriers, A.; Fonti, P.; Rossi, S.; Rathgeber, C.B.; Gricar, J. Ecophysiology and plasticity of wood and phloem formation. In Dendroecology: Tree-Ring Analyses Applied to Ecological Studies; Amoroso, M., Daniels, L., Baker, P., Camarero, J., Eds.; Springer: Cham, Germany, 2017; pp. 13–33. [Google Scholar] [CrossRef]
- Ren, P.; Rossi, S.; Camarero, J.J.; Ellison, A.M.; Liang, E.; Penuelas, J. Critical temperature and precipitation thresholds for the onset of xylogenesis of Juniperus przewalskii in a semi-arid area of the north-eastern Tibetan Plateau. Ann. Bot. 2018, 121, 617–624. [Google Scholar] [CrossRef]
- Yu, B.; Li, X.; Zhao, P.; Huang, J. Comparison of intra-annual xylem and phloem formation of Picea crassifolia stands at two latitudes in Northwest China. Forests 2021, 12, 1445. [Google Scholar] [CrossRef]
- Babushkina, E.A.; Sitnikov, G.A.; Upadhyay, K.K.; Zhirnova, D.F.; Zelenov, G.K.; Vaganov, E.A.; Belokopytova, L.V. Seasonal growth of pine tree rings: Comparison of direct observations and simulation. Forests 2022, 13, 1978. [Google Scholar] [CrossRef]
- Chen, Y.; Rademacher, T.; Fonti, P.; Eckes-Shephard, A.H.; LeMoine, J.M.; Fonti, M.V.; Richardson, A.D.; Friend, A.D. Inter-annual and inter-species tree growth explained by phenology of xylogenesis. New Phytol. 2022, 235, 939–952. [Google Scholar] [CrossRef]
- Pompa-Garcia, M.; Camarero, J.J.; Colangelo, M. Different xylogenesis responses to atmospheric water demand contribute to species coexistence in a mixed pine–oak forest. J. For. Res. 2023, 34, 51–62. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Species 1 | Coordinates | ||
|---|---|---|---|---|
| Latitude (N) | Longitude (E) | Elevation, m a.s.l. | ||
| Bayanaul (Ba) | PISY | 50°49.58′ | 75°42.30′ | 555 |
| Shalday (Sh) | PISY | 51°55.33′ | 78°43.72′ | 168 |
| Beskaragay (Be) | PISY | 50°48.04′ | 79°40.70′ | 238 |
| Zhongar-Alatau (ZA) | PCSH | 45°27.04′ | 80°27.20′ | 1190 |
| Ile-Alatau (IA) | PCSH | 43°10.30′ | 77°00.94′ | 1377 |
| Sayram-Ugam (SU) | JUSE | 42°16.41′ | 70°38.24′ | 2000 |
| Statistics | Sampling Sites | |||||
|---|---|---|---|---|---|---|
| Ba | Sh | Be | ZA | IA | SU | |
| Sample | ||||||
| no. of trees/cores | 20/20 | 23/23 | 23/23 | 25/25 | 29/29 | 12/12 |
| cover period, years | 1852–2017 | 1916–2017 | 1827–2017 | 1938–2019 | 1896–2019 | 1913–2019 |
| total length, years | 166 | 102 | 191 | 82 | 124 | 107 |
| total no. of measured rings | 1438 | 2136 | 2382 | 1264 | 1893 | 814 |
| no. of missing rings | 1 | 1 | 4 | 0 | 0 | 0 |
| mean TRW, mm | 1.255 | 2.481 | 1.247 | 2.547 | 2.178 | 1.846 |
| Standard chronologies | ||||||
| SD | 0.464 | 0.339 | 0.401 | 0.175 | 0.194 | 0.175 |
| sens | 0.288 | 0.240 | 0.219 | 0.146 | 0.181 | 0.176 |
| r-bar * | 0.525 | 0.561 | 0.537 | 0.508 | 0.497 | 0.367 |
| ar−1 | 0.742 | 0.624 | 0.766 | 0.518 | 0.369 | 0.263 |
| EPS > 0.85, years | 1946–2017 | 1941–2017 | 1913–2017 | 1963–2019 | 1930–2019 | 1961–2019 |
| Sampling Sites | Ba | Sh | Be | ZA | IA |
|---|---|---|---|---|---|
| Sh | 0.42 | ||||
| Be | 0.19 | 0.50 | |||
| ZA | 0.11 | 0.13 | 0.00 | ||
| IA | 0.07 | 0.20 | 0.16 | 0.55 | |
| SU | 0.02 | −0.12 | −0.03 | 0.22 | 0.05 |
| Variable | Characteristics | Sampling Sites | |||||
|---|---|---|---|---|---|---|---|
| Ba | Sh | Be | ZA | IA | SU | ||
| prev_Tmax | months | Jul *–Sep * | Jul *–Sep * | Aug *–Oct * | Jun *–Aug * | Jun *–Aug * | Jul *–Aug * |
| r | −0.500 | −0.456 | −0.389 | −0.459 | −0.400 | −0.371 | |
| curr_Tmax | months | May–Aug | May–Aug | May–Aug | May–Aug | Jun–Jul | Jun–Jul |
| r | −0.376 | −0.387 | −0.165 | −0.440 | −0.180 | −0.326 | |
| prev_P | months | Jul *–Sep * | Jul *–Sep * | Jul *–Oct * | Jun *–Sep * | Jun *–Oct * | Jul *–Aug * |
| r | 0.466 | 0.413 | 0.169 | 0.461 | 0.493 | 0.210 | |
| curr_P | months | May–Jul | May–Jul | May–Jul | Apr–Aug | Apr–Jul | May–Jul |
| r | 0.400 | 0.315 | 0.142 | 0.385 | 0.194 | 0.255 | |
| year_P | months | Aug *–Jul | Jul *–Jun | Jul *–Jun | Jun *–May | Jun *–May | Jul *–Jun |
| r | 0.465 | 0.367 | 0.101 | 0.477 | 0.406 | 0.287 | |
| year_PDSI | months | Aug *–Jul | Jul *–Jun | Jul *–Jun | Jun *–May | Jun *–May | Jul *–Jun |
| r | 0.402 | 0.364 | 0.154 | 0.490 | 0.471 | 0.255 | |
| Pointer Years | Sampling Sites | |||||
|---|---|---|---|---|---|---|
| Ba | Sh | Be | ZA | IA | SU | |
| Positive (max TRW indices) | 1947, 1948, 1961, 1993, 1995 | 1979, 1993, 1994, 1995, 1996 | 1913, 1970, 1971, 1972, 1995 | 1970, 1993, 2003, 2004, 2005 | 1933, 1934, 1935, 1947, 1988 | 1970, 1972, 2002, 2005, 2006 |
| Negative (min TRW indices) | 1955, 1956, 1967, 1999, 2012 | 1952, 1953, 1955, 2003, 2012 | 1927, 1955, 1956, 1957, 1958 | 1977, 1996, 1998, 2008, 2015 | 1940, 1944, 1945, 1957, 2008 | 1963, 1980, 1984, 2014, 2019 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mapitov, N.B.; Belokopytova, L.V.; Zhirnova, D.F.; Abilova, S.B.; Ualiyeva, R.M.; Bitkeyeva, A.A.; Babushkina, E.A.; Vaganov, E.A. Factors Limiting Radial Growth of Conifers on Their Semiarid Borders across Kazakhstan. Biology 2023, 12, 604. https://doi.org/10.3390/biology12040604
Mapitov NB, Belokopytova LV, Zhirnova DF, Abilova SB, Ualiyeva RM, Bitkeyeva AA, Babushkina EA, Vaganov EA. Factors Limiting Radial Growth of Conifers on Their Semiarid Borders across Kazakhstan. Biology. 2023; 12(4):604. https://doi.org/10.3390/biology12040604
Chicago/Turabian StyleMapitov, Nariman B., Liliana V. Belokopytova, Dina F. Zhirnova, Sholpan B. Abilova, Rimma M. Ualiyeva, Aliya A. Bitkeyeva, Elena A. Babushkina, and Eugene A. Vaganov. 2023. "Factors Limiting Radial Growth of Conifers on Their Semiarid Borders across Kazakhstan" Biology 12, no. 4: 604. https://doi.org/10.3390/biology12040604
APA StyleMapitov, N. B., Belokopytova, L. V., Zhirnova, D. F., Abilova, S. B., Ualiyeva, R. M., Bitkeyeva, A. A., Babushkina, E. A., & Vaganov, E. A. (2023). Factors Limiting Radial Growth of Conifers on Their Semiarid Borders across Kazakhstan. Biology, 12(4), 604. https://doi.org/10.3390/biology12040604