
The Neurobiology of Love and Pair Bonding from Human and Animal Perspectives

Abstract
:Simple Summary
Abstract

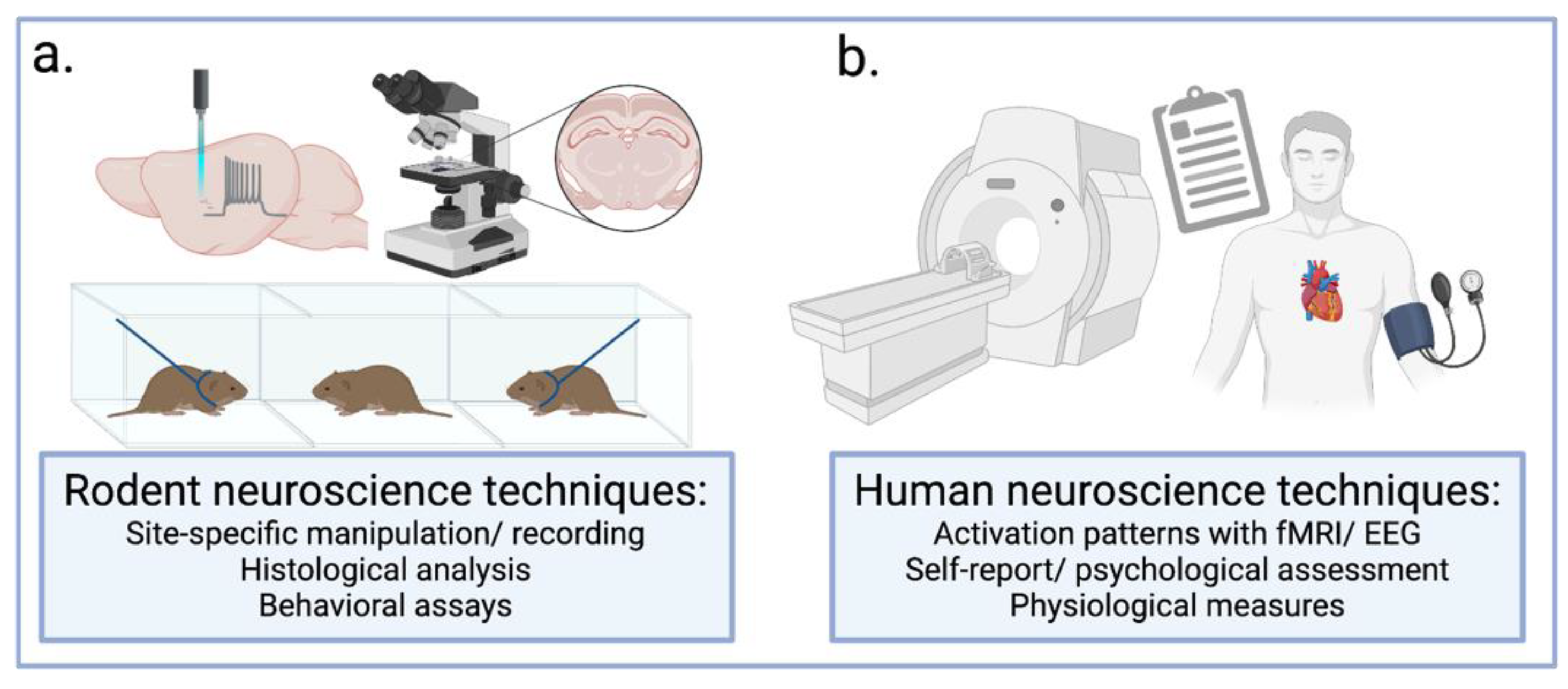
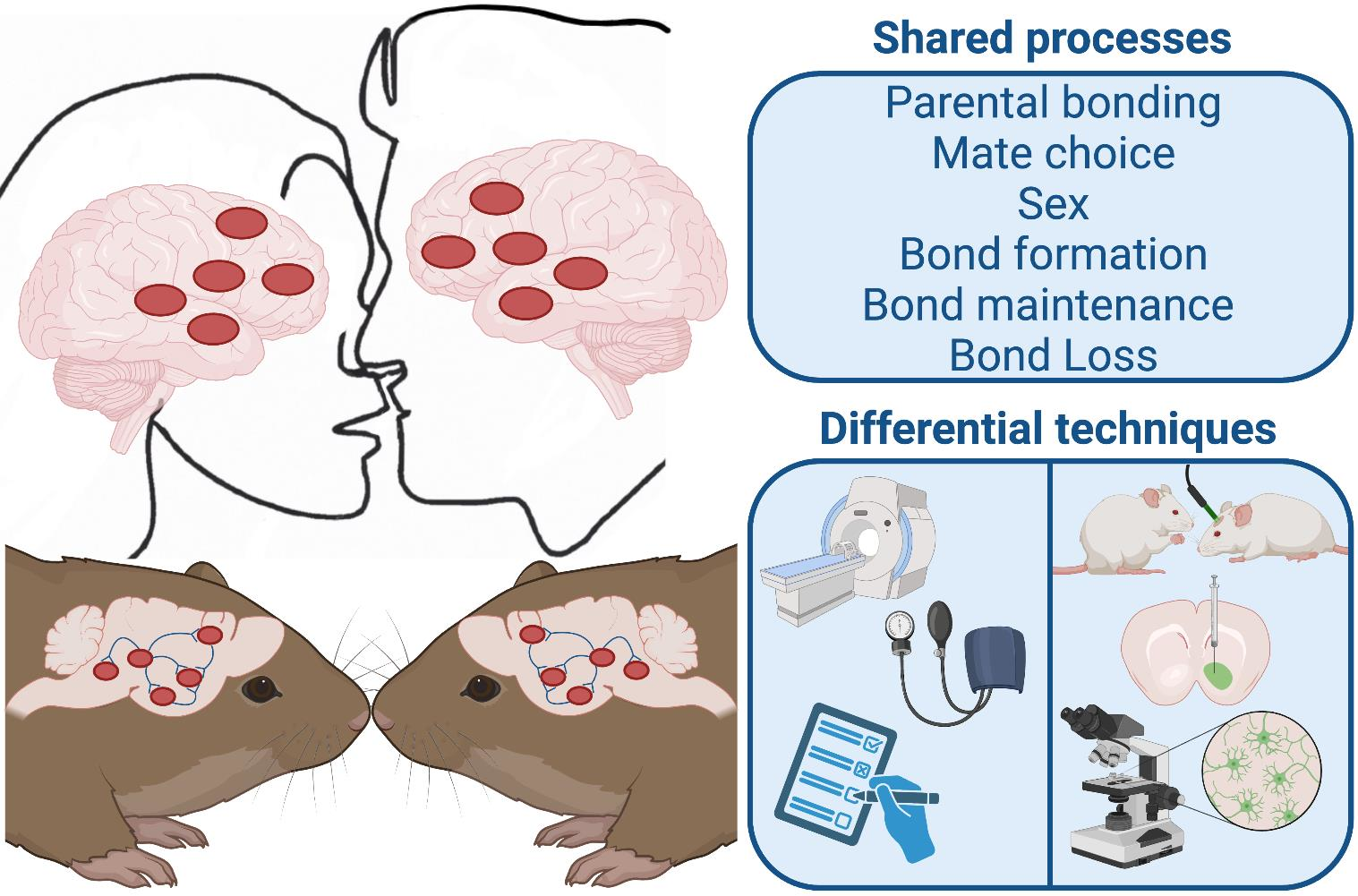{kind=link}
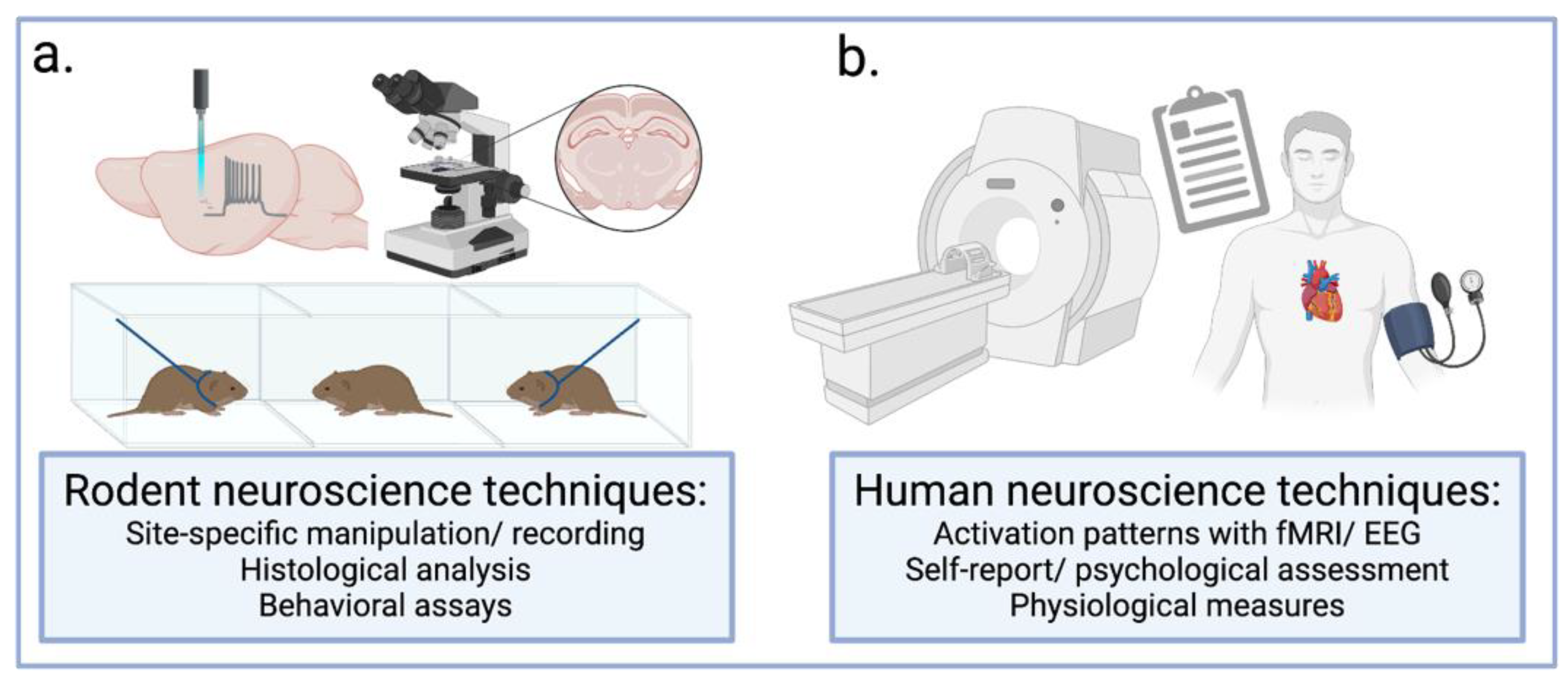{kind=link}
{kind=link}
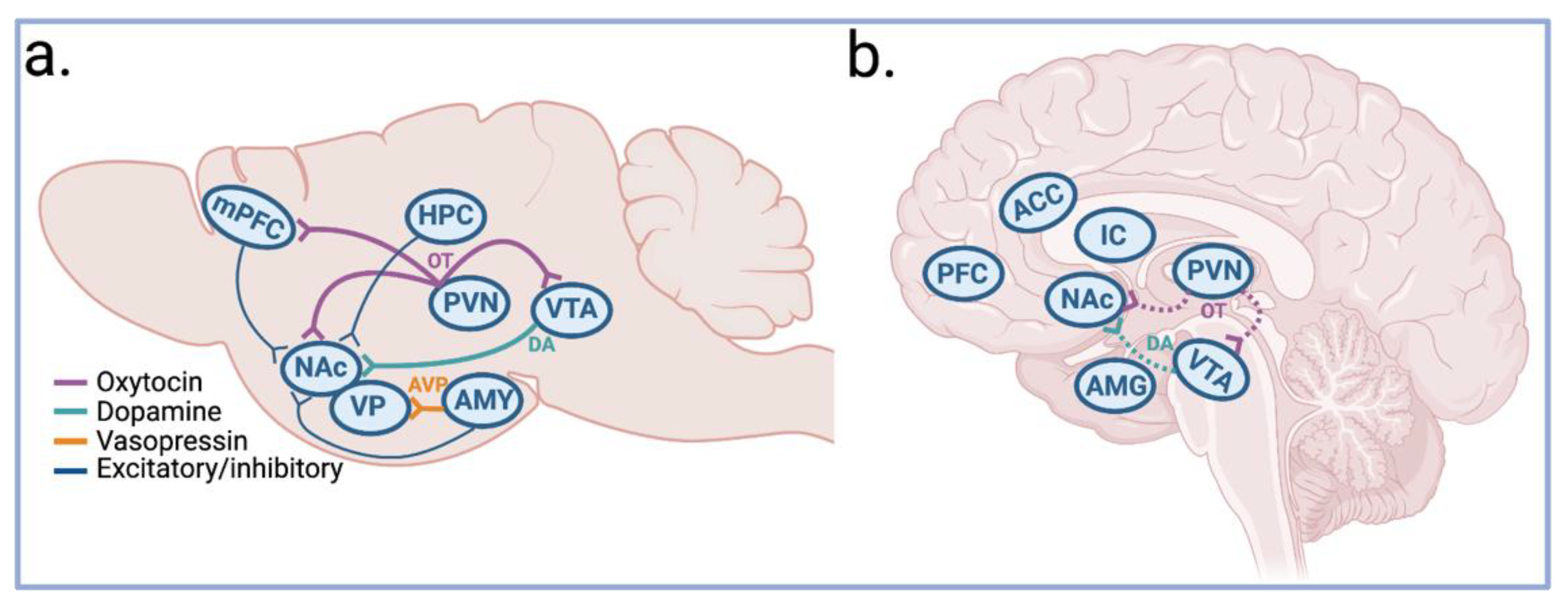{kind=link}
1. Introduction
“How little we know, how much to discover, what chemical forces flow from lover to lover”—Frank Sinatra
2. The Relationship between Parental Nurturing and Pair Bonding
2.1. Parental Bonding as the Evolutionary Antecedent to Pair Bonding
2.2. The Influence of Parental Care of Offspring in Later-Life Bonding
3. Stages of Bonding
3.1. Sexual Attraction and Mate Choice
3.2. Mating Activates OT, DA, and AVP Circuitry
3.3. Pair Bonding
3.4. Pair Bond Maintenance
3.4.1. Preventing Disruption of an Existing Pair Bond
3.4.2. Negative Reinforcement Drives Maintenance
3.5. The Healing Power of Love
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Y.C.; Boen, C.; Gerken, K.; Li, T.; Schorpp, K.; Harris, K.M. Social relationships and physiological determinants of longevity across the human life span. Proc. Natl. Acad. Sci. USA 2016, 113, 578–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, O.J.; Young, L.J. Oxytocin and Social Relationships: From Attachment to Bond Disruption. Curr. Top. Behav. Neurosci. 2018, 35, 97–117. [Google Scholar] [CrossRef] [PubMed]
- Young, L.; Alexander, B. The Chemistry between Us: Love, Sex, and the Science of Attraction; Current/Penguin Group: London, UK, 2012; 310p. [Google Scholar]
- Bales, K.L.; Ardekani, C.S.; Baxter, A.; Karaskiewicz, C.L.; Kuske, J.X.; Lau, A.R.; Savidge, L.E.; Sayler, K.R.; Witczak, L.R. What is a pair bond? Horm. Behav. 2021, 136, 105062. [Google Scholar] [CrossRef] [PubMed]
- Rilling, J.K.; Young, L.J. The biology of mammalian parenting and its effect on offspring social development. Science 2014, 345, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Marazziti, D.; Palermo, S.; Mucci, F. The Science of Love: State of the Art. Adv. Exp. Med. Biol. 2021, 1331, 249–254. [Google Scholar] [CrossRef]
- Fisher, H.E.; Aron, A.; Brown, L.L. Romantic love: A mammalian brain system for mate choice. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 2173–2186. [Google Scholar] [CrossRef] [Green Version]
- Bales, K.L.; Arias Del Razo, R.; Conklin, Q.A.; Hartman, S.; Mayer, H.S.; Rogers, F.D.; Simmons, T.C.; Smith, L.K.; Williams, A.; Williams, D.R.; et al. Titi Monkeys as a Novel Non-Human Primate Model for the Neurobiology of Pair Bonding. Yale J. Biol. Med. 2017, 90, 373–387. [Google Scholar]
- Walum, H.; Young, L.J. The neural mechanisms and circuitry of the pair bond. Nat. Rev. Neurosci. 2018, 19, 643–654. [Google Scholar] [CrossRef]
- Carter, C.S.; Perkeybile, A.M. The Monogamy Paradox: What Do Love and Sex Have to Do With It? Front. Ecol. Evol. 2018, 6, 202. [Google Scholar] [CrossRef] [Green Version]
- Lambert, C.T.; Sabol, A.C.; Solomon, N.G. Genetic Monogamy in Socially Monogamous Mammals Is Primarily Predicted by Multiple Life History Factors: A Meta-Analysis. Front. Ecol. Evol. 2018, 6, 139. [Google Scholar] [CrossRef] [Green Version]
- Lukas, D.; Clutton-Brock, T.H. The evolution of social monogamy in mammals. Science 2013, 341, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Jankowiak, W.R.; Fischer, E.F. A cross-cultural perspective on romantic love. Ethnology 1992, 31, 149–155. [Google Scholar] [CrossRef]
- Zablocki-Thomas, P.B.; Rogers, F.D.; Bales, K.L. Neuroimaging of human and non-human animal emotion and affect in the context of social relationships. Front. Behav. Neurosci. 2022, 16, 994504. [Google Scholar] [CrossRef] [PubMed]
- Keebaugh, A.C.; Barrett, C.E.; Laprairie, J.L.; Jenkins, J.J.; Young, L.J. RNAi knockdown of oxytocin receptor in the nucleus accumbens inhibits social attachment and parental care in monogamous female prairie voles. Soc. Neurosci. 2015, 10, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Hung, L.W.; Neuner, S.; Polepalli, J.S.; Beier, K.T.; Wright, M.; Walsh, J.J.; Lewis, E.M.; Luo, L.; Deisseroth, K.; Dölen, G.; et al. Gating of social reward by oxytocin in the ventral tegmental area. Science 2017, 357, 1406–1411. [Google Scholar] [CrossRef] [Green Version]
- Barrett, C.E.; Keebaugh, A.C.; Ahern, T.H.; Bass, C.E.; Terwilliger, E.F.; Young, L.J. Variation in vasopressin receptor (Avpr1a) expression creates diversity in behaviors related to monogamy in prairie voles. Horm. Behav. 2013, 63, 518–526. [Google Scholar] [CrossRef] [Green Version]
- Scheele, D.; Wille, A.; Kendrick, K.M.; Stoffel-Wagner, B.; Becker, B.; Güntürkün, O.; Maier, W.; Hurlemann, R. Oxytocin enhances brain reward system responses in men viewing the face of their female partner. Proc. Natl. Acad. Sci. USA 2013, 110, 20308–20313. [Google Scholar] [CrossRef] [Green Version]
- Walum, H.; Westberg, L.; Henningsson, S.; Neiderhiser, J.M.; Reiss, D.; Igl, W.; Ganiban, J.M.; Spotts, E.L.; Pedersen, N.L.; Eriksson, E.; et al. Genetic variation in the vasopressin receptor 1a gene (AVPR1A) associates with pair-bonding behavior in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 14153–14156. [Google Scholar] [CrossRef] [Green Version]
- Numan, M. Maternal Behavior: Neural Circuits, Stimulus Valence, and Motivational Processes. Parenting 2012, 12, 105–114. [Google Scholar] [CrossRef]
- Bridges, R.S. Neuroendocrine regulation of maternal behavior. Front. Neuroendocrinol. 2015, 36, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Terkel, J.; Rosenblatt, J.S. Maternal behavior induced by maternal blood plasma injected into virgin rats. J. Comp. Physiol. Psychol. 1968, 65, 479–482. [Google Scholar] [CrossRef] [PubMed]
- Rigney, N.; de Vries, G.J.; Petrulis, A.; Young, L.J. Oxytocin, Vasopressin, and Social Behavior: From Neural Circuits to Clinical Opportunities. Endocrinology 2022, 163, bqac111. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.J.; Christoffel, D.J.; Malenka, R.C. Neural circuits regulating prosocial behaviors. Neuropsychopharmacology 2023, 48, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.; Bakermans-Kranenburg, M.J. Oxytocin: A parenting hormone. Curr. Opin. Psychol. 2017, 15, 13–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, H.E.; Cole, C.D.; Smith, Y.; Neumann, I.D.; Landgraf, R.; Murphy, A.Z.; Young, L.J. Characterization of the oxytocin system regulating affiliative behavior in female prairie voles. Neuroscience 2009, 162, 892–903. [Google Scholar] [CrossRef] [Green Version]
- Marlin, B.J.; Mitre, M.; D’Amour, J.A.; Chao, M.V.; Froemke, R.C. Oxytocin enables maternal behaviour by balancing cortical inhibition. Nature 2015, 520, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Dwyer, C.M. Maternal behaviour and lamb survival: From neuroendocrinology to practical application. Animal 2014, 8, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Coria-Avila, G.A.; Herrera-Covarrubias, D.; García, L.I.; Toledo, R.; Hernández, M.E.; Paredes-Ramos, P.; Corona-Morales, A.A.; Manzo, J. Neurobiology of Maternal Behavior in Nonhuman Mammals: Acceptance, Recognition, Motivation, and Rejection. Animals 2022, 1, 3589. [Google Scholar] [CrossRef]
- Shahrokh, D.K.; Zhang, T.Y.; Diorio, J.; Gratton, A.; Meaney, M.J. Oxytocin-dopamine interactions mediate variations in maternal behavior in the rat. Endocrinology 2010, 151, 2276–2286. [Google Scholar] [CrossRef]
- Baik, J.H. Dopamine signaling in reward-related behaviors. Front. Neural Circuits 2013, 7, 152. [Google Scholar] [CrossRef] [Green Version]
- Akbari, E.M.; Shams, S.; Belay, H.T.; Kaiguo, M.; Razak, Z.; Kent, C.F.; Westwood, T.; Sokolowski, M.B.; Fleming, A.S. The effects of parity and maternal behavior on gene expression in the medial preoptic area and the medial amygdala in postpartum and virgin female rats: A microarray study. Behav. Neurosci. 2013, 127, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Stolzenberg, D.S.; Rissman, E.F. Oestrogen-independent, experience-induced maternal behaviour in female mice. J. Neuroendocrinol. 2011, 23, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, G.J.; Simpson, J.A.; Campbell, L.; Overall, N.C. Pair-bonding, romantic love, and evolution: The curious case of Homo sapiens. Perspect. Psychol. Sci. 2015, 10, 20–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghanti, M.A.; Edler, M.K.; Stephenson, A.R.; Munger, E.L.; Jacobs, B.; Hof, P.R.; Sherwood, C.C.; Holloway, R.L.; Lovejoy, C.O. A neurochemical hypothesis for the origin of hominids. Proc. Natl. Acad. Sci. USA 2018, 115, E1108–E1116. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Ma, H.; Xun, Y.; Hou, W.; Wang, L.; Zhang, X.; Ran, Y.; Yuan, W.; Guo, Q.; Zhang, J.; et al. Involvement of the dopamine system in paternal behavior induced by repeated pup exposure in virgin male ICR mice. Behav. Brain Res. 2021, 415, 113519. [Google Scholar] [CrossRef]
- He, Z.; Zhang, L.; Hou, W.; Zhang, X.; Young, L.J.; Li, L.; Liu, L.; Ma, H.; Xun, Y.; Lv, Z.; et al. Paraventricular Nucleus Oxytocin Subsystems Promote Active Paternal Behaviors in Mandarin Voles. J. Neurosci. 2021, 41, 6699–6713. [Google Scholar] [CrossRef]
- Weisman, O.; Zagoory-Sharon, O.; Feldman, R. Oxytocin administration to parent enhances infant physiological and behavioral readiness for social engagement. Biol. Psychiatry 2012, 72, 982–989. [Google Scholar] [CrossRef]
- Kim, P.; Rigo, P.; Mayes, L.C.; Feldman, R.; Leckman, J.F.; Swain, J.E. Neural plasticity in fathers of human infants. Soc. Neurosci. 2014, 9, 522–535. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Chen, X.; Mascaro, J.; Haroon, E.; Rilling, J.K. Intranasal oxytocin, but not vasopressin, augments neural responses to toddlers in human fathers. Horm. Behav. 2017, 93, 193–202. [Google Scholar] [CrossRef]
- Strathearn, L.; Li, J.; Fonagy, P.; Montague, P.R. What’s in a smile? Maternal brain responses to infant facial cues. Pediatrics 2008, 122, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Shih, H.C.; Kuo, M.E.; Wu, C.W.; Chao, Y.P.; Huang, H.W.; Huang, C.M. The Neurobiological Basis of Love: A Meta-Analysis of Human Functional Neuroimaging Studies of Maternal and Passionate Love. Brain Sci. 2022, 12, 830. [Google Scholar] [CrossRef] [PubMed]
- Atzil, S.; Touroutoglou, A.; Rudy, T.; Salcedo, S.; Feldman, R.; Hooker, J.M.; Dickerson, B.C.; Catana, C.; Barrett, L.F. Dopamine in the medial amygdala network mediates human bonding. Proc. Natl. Acad. Sci. USA 2017, 114, 2361–2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeevi, L.; Irani, M.; Catana, C.; Feldman Barrett, L.; Atzil, S. Maternal dopamine encodes affective signals of human infants. Soc. Cogn. Affect. Neurosci. 2022, 17, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Atzil, S.; Hendler, T.; Zagoory-Sharon, O.; Winetraub, Y.; Feldman, R. Synchrony and specificity in the maternal and the paternal brain: Relations to oxytocin and vasopressin. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 798–811. [Google Scholar] [CrossRef] [PubMed]
- Bartels, A.; Zeki, S. The neural correlates of maternal and romantic love. Neuroimage 2004, 21, 1155–1166. [Google Scholar] [CrossRef] [Green Version]
- Bartels, A.; Zeki, S. The neural basis of romantic love. Neuroreport 2000, 11, 3829–3834. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Ferris, C.F.; De Vries, G.J. Role of septal vasopressin innervation in paternal behavior in prairie voles (Microtus ochrogaster). Proc. Natl. Acad. Sci. USA 1994, 91, 400–404. [Google Scholar] [CrossRef] [Green Version]
- Bosch, O.J.; Neumann, I.D. Brain vasopressin is an important regulator of maternal behavior independent of dams’ trait anxiety. Proc. Natl. Acad. Sci. USA 2008, 105, 17139–17144. [Google Scholar] [CrossRef] [Green Version]
- Mayer, H.S.; Rosinger, Z.J.; Kruithof, V.B.; Mishra, S.; Black Owl, A.L., Sr.; Stolzenberg, D.S. Effects of maternal experience on pup-induced activation of maternal neural circuits in virgin mice. Horm. Behav. 2022, 141, 105129. [Google Scholar] [CrossRef]
- Rigney, N.; Zbib, A.; de Vries, G.J.; Petrulis, A. Knockdown of sexually differentiated vasopressin expression in the bed nucleus of the stria terminalis reduces social and sexual behaviour in male, but not female, mice. J. Neuroendocrinol. 2022, 34, e13083. [Google Scholar] [CrossRef]
- Arquilla, A.M.; Wilson, K.M.; Razak, K.A.; Saltzman, W. Fatherhood increases attraction to sensory stimuli from unrelated pups in male California mice, Peromyscus californicus. Anim. Behav. 2023, 198, 131–140. [Google Scholar] [CrossRef]
- Bowlby, J. Attachment; Basic Books: New York, NY, USA, 2008. [Google Scholar]
- Bowlby, J. The bowlby-ainsworth attachment theory. Behav. Brain Sci. 1979, 2, 637–638. [Google Scholar] [CrossRef]
- Ainsworth, M.D.S.; Blehar, M.C.; Waters, E.; Wall, S. Patterns of Attachment: A Psychological Study of the Strange Situation; Hillsdale; Lawlence Erlbaum Associates: Mahwah, NJ, USA, 1978. [Google Scholar]
- Delgado, E.; Serna, C.; Martínez, I.; Cruise, E. Parental Attachment and Peer Relationships in Adolescence: A Systematic Review. Int. J. Environ. Res. Public Health 2022, 19, 1064. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Verbeke, W.; Ein-Dor, T.; Vrtička, P. A functional neuro-anatomical model of human attachment (NAMA): Insights from first- and second-person social neuroscience. Cortex 2020, 126, 281–321. [Google Scholar] [CrossRef]
- Atzil, S.; Gao, W.; Fradkin, I.; Barrett, L.F. Growing a social brain. Nat. Hum. Behav. 2018, 2, 624–636. [Google Scholar] [CrossRef]
- Mikulincer, M.; Shaver, P.R.; Pereg, D. Attachment theory and affect regulation: The dynamics, development, and cognitive consequences of attachment-related strategies. Motiv. Emot. 2003, 27, 77–102. [Google Scholar] [CrossRef]
- River, L.M.; O’Reilly Treter, M.; Rhoades, G.K.; Narayan, A.J. Parent-Child Relationship Quality in the Family of Origin and Later Romantic Relationship Functioning: A Systematic Review. Fam. Process. 2022, 61, 259–277. [Google Scholar] [CrossRef]
- Fraley, R.C.; Roisman, G.I. The development of adult attachment styles: Four lessons. Curr. Opin. Psychol. 2019, 25, 26–30. [Google Scholar] [CrossRef]
- Kenkel, W.M.; Gustison, M.L.; Beery, A.K. A Neuroscientist’s Guide to the Vole. Curr. Protoc. 2021, 1, e175. [Google Scholar] [CrossRef]
- Perkeybile, A.M.; Griffin, L.L.; Bales, K.L. Natural variation in early parental care correlates with social behaviors in adolescent prairie voles (Microtus ochrogaster). Front. Behav. Neurosci. 2013, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Ahern, T.H.; Young, L.J. The impact of early life family structure on adult social attachment, alloparental behavior, and the neuropeptide systems regulating affiliative behaviors in the monogamous prairie vole (Microtus ochrogaster). Front. Behav. Neurosci. 2009, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Valera-Marín, G.; Young, L.J.; Camacho, F.; Paredes, R.G.; Rodríguez, V.M.; Díaz, N.F.; Portillo, W. Raised without a father: Monoparental care effects over development, sexual behavior, sexual reward, and pair bonding in prairie voles. Behav. Brain Res. 2021, 408, 113264. [Google Scholar] [CrossRef]
- Barrett, C.E.; Arambula, S.E.; Young, L.J. The oxytocin system promotes resilience to the effects of neonatal isolation on adult social attachment in female prairie voles. Transl. Psychiatry 2015, 5, e606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danoff, J.S.; Wroblewski, K.L.; Graves, A.J.; Quinn, G.C.; Perkeybile, A.M.; Kenkel, W.M.; Lillard, T.S.; Parikh, H.I.; Golino, H.F.; Gregory, S.G.; et al. Genetic, epigenetic, and environmental factors controlling oxytocin receptor gene expression. Clin. Epigenetics 2021, 13, 23. [Google Scholar] [CrossRef] [PubMed]
- Perkeybile, A.M.; Carter, C.S.; Wroblewski, K.L.; Puglia, M.H.; Kenkel, W.M.; Lillard, T.S.; Karaoli, T.; Gregory, S.G.; Mohammadi, N.; Epstein, L.; et al. Early nurture epigenetically tunes the oxytocin receptor. Psychoneuroendocrinology 2019, 99, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, T.X.; Nishitani, S.; Takiguchi, S.; Shimada, K.; Smith, A.K.; Tomoda, A. Oxytocin receptor DNA methylation and alterations of brain volumes in maltreated children. Neuropsychopharmacology 2019, 44, 2045–2053. [Google Scholar] [CrossRef]
- Ahern, T.H.; Olsen, S.; Tudino, R.; Beery, A.K. Natural variation in the oxytocin receptor gene and rearing interact to influence reproductive and nonreproductive social behavior and receptor binding. Psychoneuroendocrinology 2021, 128, 105209. [Google Scholar] [CrossRef]
- King, L.B.; Walum, H.; Inoue, K.; Eyrich, N.W.; Young, L.J. Variation in the Oxytocin Receptor Gene Predicts Brain Region-Specific Expression and Social Attachment. Biol. Psychiatry 2016, 80, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, G.G. Mate Choice: The Evolution of Sexual Decision Making from Microbes to Humans; Princeton University Press: Princeton, NJ, USA, 2017. [Google Scholar]
- Cooper, J.C.; Dunne, S.; Furey, T.; O’Doherty, J.P. Dorsomedial prefrontal cortex mediates rapid evaluations predicting the outcome of romantic interactions. J. Neurosci. 2012, 32, 15647–15656. [Google Scholar] [CrossRef] [Green Version]
- Fisher, H.; Aron, A.; Brown, L.L. Romantic love: An fMRI study of a neural mechanism for mate choice. J. Comp. Neurol. 2005, 493, 58–62. [Google Scholar] [CrossRef]
- Keyes, K.M.; Pratt, C.; Galea, S.; McLaughlin, K.A.; Koenen, K.C.; Shear, M.K. The burden of loss: Unexpected death of a loved one and psychiatric disorders across the life course in a national study. Am. J. Psychiatry 2014, 171, 864–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.H.; Yang, K.B.; Zhang, Y.Z.; Wu, C.F.; Wen, D.W.; Lv, J.W.; Zhu, G.L.; Du, X.J.; Chen, L.; Zhou, G.Q.; et al. Assessment of Modifiable Factors for the Association of Marital Status With Cancer-Specific Survival. JAMA Netw. Open 2021, 4, e2111813. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Lubetkin, E.I. Life expectancy and active life expectancy by marital status among older U.S. adults: Results from the U.S. Medicare Health Outcome Survey (HOS). SSM Popul. Health 2020, 12, 100642. [Google Scholar] [CrossRef] [PubMed]
- Newman, S.W. The medial extended amygdala in male reproductive behavior. A node in the mammalian social behavior network. Ann. N. Y. Acad. Sci. 1999, 877, 242–257. [Google Scholar] [CrossRef] [PubMed]
- Goodson, J.L. The vertebrate social behavior network: Evolutionary themes and variations. Horm. Behav. 2005, 48, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, L.A.; Hofmann, H.A. The vertebrate mesolimbic reward system and social behavior network: A comparative synthesis. J. Comp. Neurol. 2011, 519, 3599–3639. [Google Scholar] [CrossRef]
- Hiser, J.; Koenigs, M. The Multifaceted Role of the Ventromedial Prefrontal Cortex in Emotion, Decision Making, Social Cognition, and Psychopathology. Biol. Psychiatry 2018, 83, 638–647. [Google Scholar] [CrossRef]
- Bernhardt, B.C.; Singer, T. The neural basis of empathy. Annu. Rev. Neurosci. 2012, 35, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Rogers-Carter, M.M.; Christianson, J.P. An insular view of the social decision-making network. Neurosci. Biobehav. Rev. 2019, 103, 119–132. [Google Scholar] [CrossRef]
- Roth, T.S.; Samara, I.; Kret, M.E. Multimodal mate choice: Exploring the effects of sight, sound, and scent on partner choice in a speed-date paradigm. Evol. Hum. Behav. 2021, 42, 461–468. [Google Scholar] [CrossRef]
- Halliday, T. The study of mate choice. Mate Choice 1983, 1, 462. [Google Scholar]
- Ferkin, M.H. Odor Communication and Mate Choice in Rodents. Biology 2018, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Kavaliers, M.; Choleris, E.; Agmo, A.; Braun, W.J.; Colwell, D.D.; Muglia, L.J.; Ogawa, S.; Pfaff, D.W. Inadvertent social information and the avoidance of parasitized male mice: A role for oxytocin. Proc. Natl. Acad. Sci. USA 2006, 103, 4293–4298. [Google Scholar] [CrossRef] [Green Version]
- Oettl, L.L.; Ravi, N.; Schneider, M.; Scheller, M.F.; Schneider, P.; Mitre, M.; da Silva Gouveia, M.; Froemke, R.C.; Chao, M.V.; Young, W.S.; et al. Oxytocin Enhances Social Recognition by Modulating Cortical Control of Early Olfactory Processing. Neuron 2016, 90, 609–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuyama, T. Social memory engram in the hippocampus. Neurosci. Res. 2018, 129, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Lenschow, C.; Mendes, A.R.P.; Lima, S.Q. Hearing, touching, and multisensory integration during mate choice. Front. Neural Circuits 2022, 16, 943888. [Google Scholar] [CrossRef] [PubMed]
- Asaba, A.; Osakada, T.; Touhara, K.; Kato, M.; Mogi, K.; Kikusui, T. Male mice ultrasonic vocalizations enhance female sexual approach and hypothalamic kisspeptin neuron activity. Horm. Behav. 2017, 94, 53–60. [Google Scholar] [CrossRef]
- Neunuebel, J.P.; Taylor, A.L.; Arthur, B.J.; Egnor, S.E. Female mice ultrasonically interact with males during courtship displays. eLife 2015, 4, e06203. [Google Scholar] [CrossRef]
- Warren, M.R.; Clein, R.S.; Spurrier, M.S.; Roth, E.D.; Neunuebel, J.P. Ultrashort-range, high-frequency communication by female mice shapes social interactions. Sci. Rep. 2020, 10, 2637. [Google Scholar] [CrossRef] [Green Version]
- Bobrov, E.; Wolfe, J.; Rao, R.P.; Brecht, M. The representation of social facial touch in rat barrel cortex. Curr. Biol. 2014, 24, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Miao, W.; Ji, E.; Huang, S.; Jin, S.; Zhu, X.; Liu, M.Z.; Sun, Y.G.; Xu, F.; Yu, X. Social touch-like tactile stimulation activates a tachykinin 1-oxytocin pathway to promote social interactions. Neuron 2022, 110, 1051–1067.e7. [Google Scholar] [CrossRef] [PubMed]
- Elias, L.J.; Succi, I.K.; Schaffler, M.D.; Foster, W.; Gradwell, M.A.; Bohic, M.; Fushiki, A.; Upadhyay, A.; Ejoh, L.L.; Schwark, R.; et al. Touch neurons underlying dopaminergic pleasurable touch and sexual receptivity. Cell 2023, 186, 577–590.e16. [Google Scholar] [CrossRef] [PubMed]
- Ronald, K.L.; Zhang, X.; Morrison, M.V.; Miller, R.; Hurley, L.M. Male mice adjust courtship behavior in response to female multimodal signals. PLoS ONE 2020, 15, e0229302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, S.M.; Smith, A.L.; Goodman, M.M.; Bales, K.L. Selective localization of oxytocin receptors and vasopressin 1a receptors in the human brainstem. Soc. Neurosci. 2017, 12, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Freeman, S.M.; Inoue, K.; Smith, A.L.; Goodman, M.M.; Young, L.J. The neuroanatomical distribution of oxytocin receptor binding and mRNA in the male rhesus macaque (Macaca mulatta). Psychoneuroendocrinology 2014, 45, 128–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grace, S.A.; Rossell, S.L.; Heinrichs, M.; Kordsachia, C.; Labuschagne, I. Oxytocin and brain activity in humans: A systematic review and coordinate-based meta-analysis of functional MRI studies. Psychoneuroendocrinology 2018, 96, 6–24. [Google Scholar] [CrossRef] [PubMed]
- Little, A.C. Facial attractiveness. Wiley Interdiscip. Rev. Cogn. Sci. 2014, 5, 621–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, J.J.; Anglin, J.M. Facial attractiveness predicts longevity. Evol. Hum. Behav. 2003, 24, 351–356. [Google Scholar] [CrossRef]
- Pflüger, L.S.; Oberzaucher, E.; Katina, S.; Holzleitner, I.J.; Grammer, K. Cues to fertility: Perceived attractiveness and facial shape predict reproductive success. Evol. Hum. Behav. 2012, 33, 708–714. [Google Scholar] [CrossRef]
- Chatterjee, A.; Thomas, A.; Smith, S.E.; Aguirre, G.K. The neural response to facial attractiveness. Neuropsychology 2009, 23, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Pegors, T.K.; Kable, J.W.; Chatterjee, A.; Epstein, R.A. Common and unique representations in pFC for face and place attractiveness. J. Cogn. Neurosci. 2015, 27, 959–973. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.C.; Dunne, S.; Furey, T.; O’Doherty, J.P. The role of the posterior temporal and medial prefrontal cortices in mediating learning from romantic interest and rejection. Cereb. Cortex 2014, 24, 2502–2511. [Google Scholar] [CrossRef] [PubMed]
- Hsu, D.T.; Sankar, A.; Malik, M.A.; Langenecker, S.A.; Mickey, B.J.; Love, T.M. Common neural responses to romantic rejection and acceptance in healthy adults. Soc. Neurosci. 2020, 15, 571–583. [Google Scholar] [CrossRef]
- Kross, E.; Berman, M.G.; Mischel, W.; Smith, E.E.; Wager, T.D. Social rejection shares somatosensory representations with physical pain. Proc. Natl. Acad. Sci. USA 2011, 108, 6270–6275. [Google Scholar] [CrossRef] [Green Version]
- Uddin, L.Q. Salience processing and insular cortical function and dysfunction. Nat. Rev. Neurosci. 2015, 16, 55–61. [Google Scholar] [CrossRef]
- Burgos-Robles, A.; Gothard, K.M.; Monfils, M.H.; Morozov, A.; Vicentic, A. Conserved features of anterior cingulate networks support observational learning across species. Neurosci. Biobehav. Rev. 2019, 107, 215–228. [Google Scholar] [CrossRef]
- Sokhi, D.S.; Hunter, M.D.; Wilkinson, I.D.; Woodruff, P.W. Male and female voices activate distinct regions in the male brain. Neuroimage 2005, 27, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Andics, A.; McQueen, J.M.; Petersson, K.M.; Gál, V.; Rudas, G.; Vidnyánszky, Z. Neural mechanisms for voice recognition. Neuroimage 2010, 52, 1528–1540. [Google Scholar] [CrossRef] [PubMed]
- Suvilehto, J.T.; Glerean, E.; Dunbar, R.I.; Hari, R.; Nummenmaa, L. Topography of social touching depends on emotional bonds between humans. Proc. Natl. Acad. Sci. USA 2015, 112, 13811–13816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotz, S.A.; Kalberlah, C.; Bahlmann, J.; Friederici, A.D.; Haynes, J.D. Predicting vocal emotion expressions from the human brain. Hum. Brain Mapp. 2013, 34, 1971–1981. [Google Scholar] [CrossRef]
- Ebisch, S.J.; Ferri, F.; Gallese, V. Touching moments: Desire modulates the neural anticipation of active romantic caress. Front. Behav. Neurosci. 2014, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prochazkova, E.; Sjak-Shie, E.; Behrens, F.; Lindh, D.; Kret, M.E. Physiological synchrony is associated with attraction in a blind date setting. Nat. Hum. Behav. 2022, 6, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Laurent, R.; Toupance, B.; Chaix, R. Non-random mate choice in humans: Insights from a genome scan. Mol. Ecol. 2012, 21, 587–596. [Google Scholar] [CrossRef]
- Thomas, A.G.; Jonason, P.K.; Blackburn, J.D.; Kennair, L.E.O.; Lowe, R.; Malouff, J.; Stewart-Williams, S.; Sulikowski, D.; Li, N.P. Mate preference priorities in the East and West: A cross-cultural test of the mate preference priority model. J. Pers. 2020, 88, 606–620. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, J.M.; Hull, E.M. Dopamine, the medial preoptic area, and male sexual behavior. Physiol. Behav. 2005, 86, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Hull, E.M.; Dominguez, J.M. Neuroendocrine Regulation of Male Sexual Behavior. Compr. Physiol. 2019, 9, 1383–1410. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.V.; Manoli, D.S.; Fraser, E.J.; Coats, J.K.; Tollkuhn, J.; Honda, S.; Harada, N.; Shah, N.M. Estrogen masculinizes neural pathways and sex-specific behaviors. Cell 2009, 139, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Oti, T.; Satoh, K.; Uta, D.; Nagafuchi, J.; Tateishi, S.; Ueda, R.; Takanami, K.; Young, L.J.; Galione, A.; Morris, J.F.; et al. Oxytocin Influences Male Sexual Activity via Non-synaptic Axonal Release in the Spinal Cord. Curr. Biol. 2021, 31, 103–114.e5. [Google Scholar] [CrossRef]
- Waldherr, M.; Neumann, I.D. Centrally released oxytocin mediates mating-induced anxiolysis in male rats. Proc. Natl. Acad. Sci. USA 2007, 104, 16681–16684. [Google Scholar] [CrossRef] [Green Version]
- Johnson, Z.V.; Walum, H.; Xiao, Y.; Riefkohl, P.C.; Young, L.J. Oxytocin receptors modulate a social salience neural network in male prairie voles. Horm. Behav. 2017, 87, 16–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaus, J.G.; Damsma, G.; Wenkstern, D.; Fibiger, H.C. Sexual activity increases dopamine transmission in the nucleus accumbens and striatum of female rats. Brain Res. 1995, 693, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Amadei, E.A.; Johnson, Z.V.; Jun Kwon, Y.; Shpiner, A.C.; Saravanan, V.; Mays, W.D.; Ryan, S.J.; Walum, H.; Rainnie, D.G.; Young, L.J.; et al. Dynamic corticostriatal activity biases social bonding in monogamous female prairie voles. Nature 2017, 546, 297–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasper, E.R.; LaMarca, E.A.; Bocarsly, M.E.; Fasolino, M.; Opendak, M.; Gould, E. Sexual experience enhances cognitive flexibility and dendritic spine density in the medial prefrontal cortex. Neurobiol. Learn. Mem. 2015, 125, 73–79. [Google Scholar] [CrossRef]
- Spritzer, M.D.; Weinberg, A.; Viau, V.; Galea, L.A. Prior sexual experience increases hippocampal cell proliferation and decreases risk assessment behavior in response to acute predator odor stress in the male rat. Behav. Brain Res. 2009, 200, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagerer, S.; Klucken, T.; Wehrum, S.; Zimmermann, M.; Schienle, A.; Walter, B.; Vaitl, D.; Stark, R. Neural activation toward erotic stimuli in homosexual and heterosexual males. J. Sex. Med. 2011, 8, 3132–3143. [Google Scholar] [CrossRef] [PubMed]
- Karama, S.; Lecours, A.R.; Leroux, J.M.; Bourgouin, P.; Beaudoin, G.; Joubert, S.; Beauregard, M. Areas of brain activation in males and females during viewing of erotic film excerpts. Hum. Brain Mapp. 2002, 16, 1–13. [Google Scholar] [CrossRef]
- Ferretti, A.; Caulo, M.; Del Gratta, C.; Di Matteo, R.; Merla, A.; Montorsi, F.; Pizzella, V.; Pompa, P.; Rigatti, P.; Rossini, P.M.; et al. Dynamics of male sexual arousal: Distinct components of brain activation revealed by fMRI. Neuroimage 2005, 26, 1086–1096. [Google Scholar] [CrossRef]
- Leon-Carrion, J.; Martín-Rodríguez, J.F.; Damas-López, J.; Pourrezai, K.; Izzetoglu, K.; Barroso, Y.M.J.M.; Dominguez-Morales, M.R. Does dorsolateral prefrontal cortex (DLPFC) activation return to baseline when sexual stimuli cease? The role of DLPFC in visual sexual stimulation. Neurosci. Lett. 2007, 416, 55–60. [Google Scholar] [CrossRef]
- Komisaruk, B.R.; Whipple, B.; Crawford, A.; Liu, W.C.; Kalnin, A.; Mosier, K. Brain activation during vaginocervical self-stimulation and orgasm in women with complete spinal cord injury: fMRI evidence of mediation by the vagus nerves. Brain Res. 2004, 1024, 77–88. [Google Scholar] [CrossRef]
- Holstege, G.; Georgiadis, J.R.; Paans, A.M.; Meiners, L.C.; van der Graaf, F.H.; Reinders, A.A. Brain activation during human male ejaculation. J. Neurosci. 2003, 23, 9185–9193. [Google Scholar] [CrossRef]
- Wise, N.J.; Frangos, E.; Komisaruk, B.R. Brain Activity Unique to Orgasm in Women: An fMRI Analysis. J. Sex. Med. 2017, 14, 1380–1391. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, M.S.; Humbert, R.; Dixen, J.; Palmisano, G.; Greenleaf, W.; Davidson, J.M. Plasma oxytocin increases in the human sexual response. J. Clin. Endocrinol. Metab. 1987, 64, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.R.; Catania, K.C.; Carter, C.S. Development of partner preferences in female prairie voles (Microtus ochrogaster): The role of social and sexual experience. Horm. Behav. 1992, 26, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.M.; Wang, Z.; Olazábal, D.E.; Ren, X.; Terwilliger, E.F.; Young, L.J. Enhanced partner preference in a promiscuous species by manipulating the expression of a single gene. Nature 2004, 429, 754–757. [Google Scholar] [CrossRef]
- Insel, T.R.; Shapiro, L.E. Oxytocin receptor distribution reflects social organization in monogamous and polygamous voles. Proc. Natl. Acad. Sci. USA 1992, 89, 5981–5985. [Google Scholar] [CrossRef] [Green Version]
- Keebaugh, A.C.; Young, L.J. Increasing oxytocin receptor expression in the nucleus accumbens of pre-pubertal female prairie voles enhances alloparental responsiveness and partner preference formation as adults. Horm. Behav. 2011, 60, 498–504. [Google Scholar] [CrossRef] [Green Version]
- Aragona, B.J.; Liu, Y.; Yu, Y.J.; Curtis, J.T.; Detwiler, J.M.; Insel, T.R.; Wang, Z. Nucleus accumbens dopamine differentially mediates the formation and maintenance of monogamous pair bonds. Nat. Neurosci. 2006, 9, 133–139. [Google Scholar] [CrossRef]
- Aragona, B.J.; Liu, Y.; Curtis, J.T.; Stephan, F.K.; Wang, Z. A critical role for nucleus accumbens dopamine in partner-preference formation in male prairie voles. J. Neurosci. 2003, 23, 3483–3490. [Google Scholar] [CrossRef]
- Gingrich, B.; Liu, Y.; Cascio, C.; Wang, Z.; Insel, T.R. Dopamine D2 receptors in the nucleus accumbens are important for social attachment in female prairie voles (Microtus ochrogaster). Behav. Neurosci. 2000, 114, 173–183. [Google Scholar] [CrossRef]
- Donaldson, Z.R.; Spiegel, L.; Young, L.J. Central vasopressin V1a receptor activation is independently necessary for both partner preference formation and expression in socially monogamous male prairie voles. Behav. Neurosci. 2010, 124, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Phelps, S.M.; Young, L.J. Extraordinary diversity in vasopressin (V1a) receptor distributions among wild prairie voles (Microtus ochrogaster): Patterns of variation and covariation. J. Comp. Neurol. 2003, 466, 564–576. [Google Scholar] [CrossRef]
- Pitkow, L.J.; Sharer, C.A.; Ren, X.; Insel, T.R.; Terwilliger, E.F.; Young, L.J. Facilitation of affiliation and pair-bond formation by vasopressin receptor gene transfer into the ventral forebrain of a monogamous vole. J. Neurosci. 2001, 21, 7392–7396. [Google Scholar] [CrossRef] [Green Version]
- Grebe, N.M.; Sharma, A.; Freeman, S.M.; Palumbo, M.C.; Patisaul, H.B.; Bales, K.L.; Drea, C.M. Neural correlates of mating system diversity: Oxytocin and vasopressin receptor distributions in monogamous and non-monogamous Eulemur. Sci. Rep. 2021, 11, 3746. [Google Scholar] [CrossRef]
- Turner, L.M.; Young, A.R.; Römpler, H.; Schöneberg, T.; Phelps, S.M.; Hoekstra, H.E. Monogamy evolves through multiple mechanisms: Evidence from V1aR in deer mice. Mol. Biol. Evol. 2010, 27, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Monin, J.K.; Goktas, S.O.; Kershaw, T.; DeWan, A. Associations between spouses’ oxytocin receptor gene polymorphism, attachment security, and marital satisfaction. PLoS ONE 2019, 14, e0213083. [Google Scholar] [CrossRef] [PubMed]
- Ueda, R.; Abe, N. Neural Representations of the Committed Romantic Partner in the Nucleus Accumbens. Psychol. Sci. 2021, 32, 1884–1895. [Google Scholar] [CrossRef] [PubMed]
- Ophir, A.G.; Phelps, S.M.; Sorin, A.B.; Wolff, J.O. Social but not genetic monogamy is associated with greater breeding success in prairie voles. Anim. Behav. 2008, 75, 1143–1154. [Google Scholar] [CrossRef]
- Rice, M.A.; Restrepo, L.F.; Ophir, A.G. When to Cheat: Modeling Dynamics of Paternity and Promiscuity in Socially Monogamous Prairie Voles (Microtus ochrogaster). Front. Ecol. Evol. 2018, 6, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okhovat, M.; Berrio, A.; Wallace, G.; Ophir, A.G.; Phelps, S.M. Sexual fidelity trade-offs promote regulatory variation in the prairie vole brain. Science 2015, 350, 1371–1374. [Google Scholar] [CrossRef]
- Gobrogge, K.L.; Liu, Y.; Young, L.J.; Wang, Z. Anterior hypothalamic vasopressin regulates pair-bonding and drug-induced aggression in a monogamous rodent. Proc. Natl. Acad. Sci. USA 2009, 106, 19144–19149. [Google Scholar] [CrossRef] [Green Version]
- Holley, A.; Bellevue, S.; Vosberg, D.; Wenzel, K.; Roorda, S., Jr.; Pfaus, J.G. The role of oxytocin and vasopressin in conditioned mate guarding behavior in the female rat. Physiol. Behav. 2015, 144, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Resendez, S.L.; Kuhnmuench, M.; Krzywosinski, T.; Aragona, B.J. κ-Opioid receptors within the nucleus accumbens shell mediate pair bond maintenance. J. Neurosci. 2012, 32, 6771–6784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resendez, S.L.; Keyes, P.C.; Day, J.J.; Hambro, C.; Austin, C.J.; Maina, F.K.; Eidson, L.N.; Porter-Stransky, K.A.; Nevárez, N.; McLean, J.W.; et al. Dopamine and opioid systems interact within the nucleus accumbens to maintain monogamous pair bonds. eLife 2016, 5, e15325. [Google Scholar] [CrossRef]
- Maninger, N.; Mendoza, S.P.; Williams, D.R.; Mason, W.A.; Cherry, S.R.; Rowland, D.J.; Schaefer, T.; Bales, K.L. Imaging, Behavior and Endocrine Analysis of “Jealousy” in a Monogamous Primate. Front. Ecol. Evol. 2017, 5, 66. [Google Scholar] [CrossRef] [Green Version]
- Mercier, F.; Witczak, L.R.; Bales, K.L. Coppery titi monkey (Plecturocebus cupreus) pairs display coordinated behaviors in response to a simulated intruder. Am. J. Primatol. 2020, 82, e23141. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Luo, L.; Li, J.; Xu, L.; Zhou, F.; Gao, Z.; Becker, B.; Kendrick, K.M. A dimensional approach to jealousy reveals enhanced fronto-striatal, insula and limbic responses to angry faces. Brain Struct. Funct. 2019, 224, 3201–3212. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Nishimura, F.; Kakiuchi, C.; Kasai, K.; Kimura, M.; Haruno, M. Interactive effects of OXTR and GAD1 on envy-associated behaviors and neural responses. PLoS ONE 2019, 14, e0210493. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.L.; Berkman, E.T.; Karremans, J.C.; Lieberman, M.D. Incidental regulation of attraction: The neural basis of the derogation of attractive alternatives in romantic relationships. Cogn. Emot. 2011, 25, 490–505. [Google Scholar] [CrossRef] [Green Version]
- Ueda, R.; Yanagisawa, K.; Ashida, H.; Abe, N. Executive control and faithfulness: Only long-term romantic relationships require prefrontal control. Exp. Brain Res. 2018, 236, 821–828. [Google Scholar] [CrossRef]
- Scheele, D.; Striepens, N.; Güntürkün, O.; Deutschländer, S.; Maier, W.; Kendrick, K.M.; Hurlemann, R. Oxytocin modulates social distance between males and females. J. Neurosci. 2012, 32, 16074–16079. [Google Scholar] [CrossRef] [Green Version]
- McNeal, N.; Scotti, M.A.; Wardwell, J.; Chandler, D.L.; Bates, S.L.; Larocca, M.; Trahanas, D.M.; Grippo, A.J. Disruption of social bonds induces behavioral and physiological dysregulation in male and female prairie voles. Auton. Neurosci. 2014, 180, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, O.J.; Nair, H.P.; Ahern, T.H.; Neumann, I.D.; Young, L.J. The CRF system mediates increased passive stress-coping behavior following the loss of a bonded partner in a monogamous rodent. Neuropsychopharmacology 2009, 34, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Osako, Y.; Hidaka, C.; Nishihara, M.; Young, L.J.; Mitsui, S.; Yuri, K. Separation from a bonded partner alters neural response to inflammatory pain in monogamous rodents. Behav. Brain Res. 2022, 418, 113650. [Google Scholar] [CrossRef] [PubMed]
- Reul, J.M.; Holsboer, F. On the role of corticotropin-releasing hormone receptors in anxiety and depression. Dialogues Clin. Neurosci. 2002, 4, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Bosch, O.J.; Dabrowska, J.; Modi, M.E.; Johnson, Z.V.; Keebaugh, A.C.; Barrett, C.E.; Ahern, T.H.; Guo, J.; Grinevich, V.; Rainnie, D.G.; et al. Oxytocin in the nucleus accumbens shell reverses CRFR2-evoked passive stress-coping after partner loss in monogamous male prairie voles. Psychoneuroendocrinology 2016, 64, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Pohl, T.T.; Young, L.J.; Bosch, O.J. Lost connections: Oxytocin and the neural, physiological, and behavioral consequences of disrupted relationships. Int. J. Psychophysiol. 2019, 136, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.; Tokunaga, R.S. Romantic Partner Monitoring After Breakups: Attachment, Dependence, Distress, and Post-Dissolution Online Surveillance via Social Networking Sites. Cyberpsychol. Behav. Soc. Netw. 2015, 18, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Carey, I.M.; Shah, S.M.; DeWilde, S.; Harris, T.; Victor, C.R.; Cook, D.G. Increased risk of acute cardiovascular events after partner bereavement: A matched cohort study. JAMA Intern. Med. 2014, 174, 598–605. [Google Scholar] [CrossRef] [Green Version]
- Gerra, G.; Monti, D.; Panerai, A.E.; Sacerdote, P.; Anderlini, R.; Avanzini, P.; Zaimovic, A.; Brambilla, F.; Franceschi, C. Long-term immune-endocrine effects of bereavement: Relationships with anxiety levels and mood. Psychiatry Res. 2003, 121, 145–158. [Google Scholar] [CrossRef]
- O’Connor, M.F.; Wellisch, D.K.; Stanton, A.L.; Eisenberger, N.I.; Irwin, M.R.; Lieberman, M.D. Craving love? Enduring grief activates brain’s reward center. Neuroimage 2008, 42, 969–972. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.L.R.; Davila, C.D.; Shah, M.; Wheeler, D.S.; Ziccardi, M.R.; Banerji, S.; Figueredo, V.M. Marital status and living condition as predictors of mortality and readmissions among African Americans with heart failure. Int. J. Cardiol. 2016, 222, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Naderi, A.; Soltanmaohammadi, E.; Kaza, V.; Barlow, S.; Chatzistamou, I.; Kiaris, H. Persistent effects of pair bonding in lung cancer cell growth in monogamous Peromyscus californicus. eLife 2021, 10, e64711. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Liu, S.; Bai, X.; Gao, Y.; Liu, G.; Wang, X.; Liu, D.; Li, T.; Hao, A.; Wang, Z. Oxytocin inhibits lipopolysaccharide-induced inflammation in microglial cells and attenuates microglial activation in lipopolysaccharide-treated mice. J. Neuroinflamm. 2016, 13, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouin, J.P.; Carter, C.S.; Pournajafi-Nazarloo, H.; Glaser, R.; Malarkey, W.B.; Loving, T.J.; Stowell, J.; Kiecolt-Glaser, J.K. Marital behavior, oxytocin, vasopressin, and wound healing. Psychoneuroendocrinology 2010, 35, 1082–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordt, E.A.; Smith, C.J.; Demarest, T.G.; Bilbo, S.D.; Kingsbury, M.A. Mitochondria, Oxytocin, and Vasopressin: Unfolding the Inflammatory Protein Response. Neurotox. Res. 2019, 36, 239–256. [Google Scholar] [CrossRef]
- Cuneo, M.G.; Szeto, A.; Schrepf, A.; Kinner, E.M.; Schachner, B.I.; Ahmed, R.; Thaker, P.H.; Goodheart, M.; Bender, D.; Cole, S.W.; et al. Oxytocin in the tumor microenvironment is associated with lower inflammation and longer survival in advanced epithelial ovarian cancer patients. Psychoneuroendocrinology 2019, 106, 244–251. [Google Scholar] [CrossRef]
- Kramer, K.L.; Schacht, R.; Bell, A. Adult sex ratios and partner scarcity among hunter-gatherers: Implications for dispersal patterns and the evolution of human sociality. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160316. [Google Scholar] [CrossRef] [Green Version]
- Schacht, R.; Kramer, K.L. Are we monogamous? A review of the evolution of pair-bonding in humans and its contemporary variation cross-culturally. Front. Ecol. Evol. 2019, 7, 230. [Google Scholar] [CrossRef] [Green Version]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blumenthal, S.A.; Young, L.J. The Neurobiology of Love and Pair Bonding from Human and Animal Perspectives. Biology 2023, 12, 844. https://doi.org/10.3390/biology12060844
Blumenthal SA, Young LJ. The Neurobiology of Love and Pair Bonding from Human and Animal Perspectives. Biology. 2023; 12(6):844. https://doi.org/10.3390/biology12060844
Chicago/Turabian StyleBlumenthal, Sarah A., and Larry J. Young. 2023. "The Neurobiology of Love and Pair Bonding from Human and Animal Perspectives" Biology 12, no. 6: 844. https://doi.org/10.3390/biology12060844
APA StyleBlumenthal, S. A., & Young, L. J. (2023). The Neurobiology of Love and Pair Bonding from Human and Animal Perspectives. Biology, 12(6), 844. https://doi.org/10.3390/biology12060844