


White LED Lighting Increases the Root Productivity of Panax ginseng C. A. Meyer in a Hydroponic Cultivation System of a Plant Factory

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
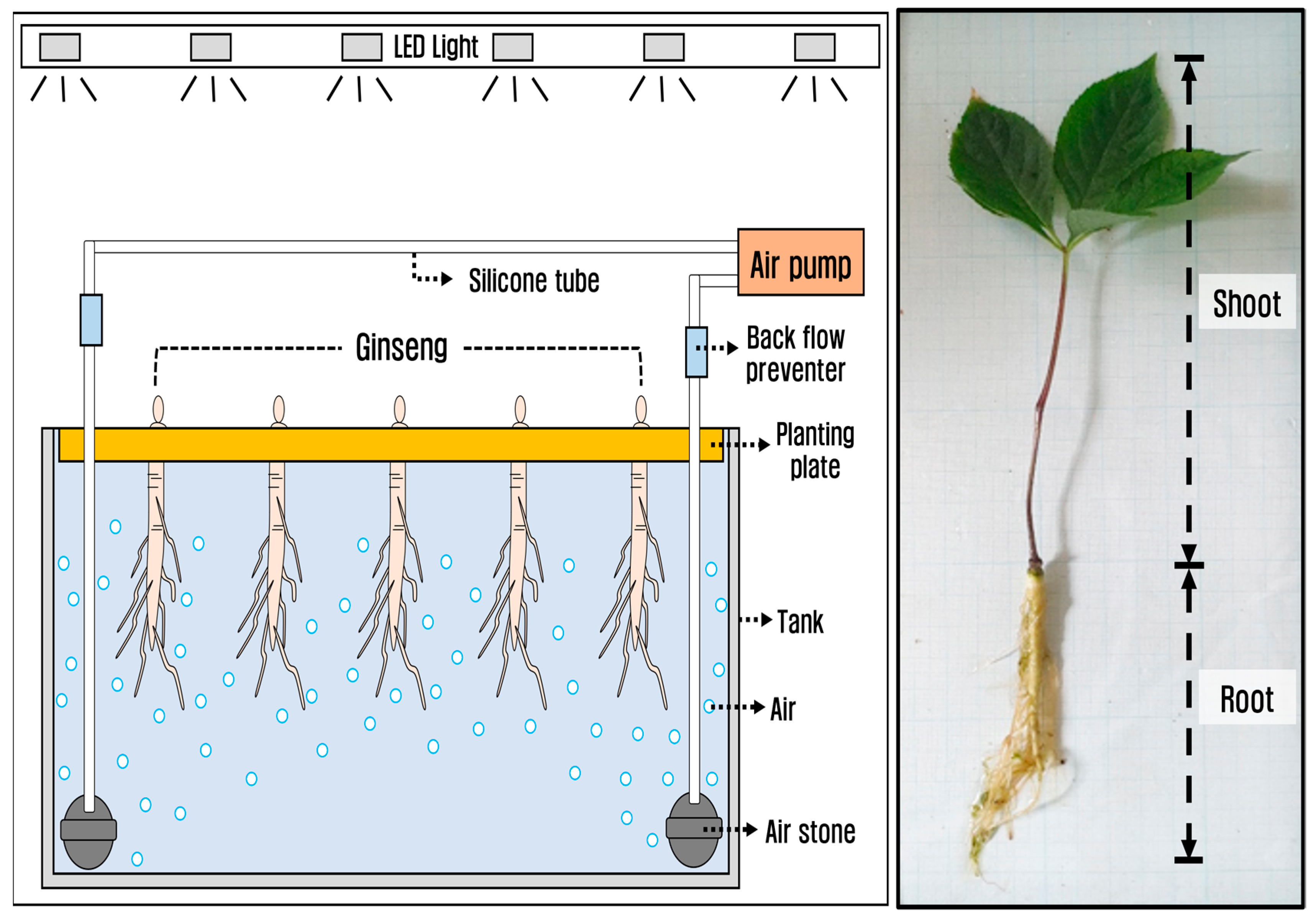
2.1. Planting and Cultivation of P. ginseng
2.2. Light Environment Conditions
2.3. Measurement Parameters
2.4. Data Analysis
3. Results
3.1. Physiological Responses
3.2. Growth Responses
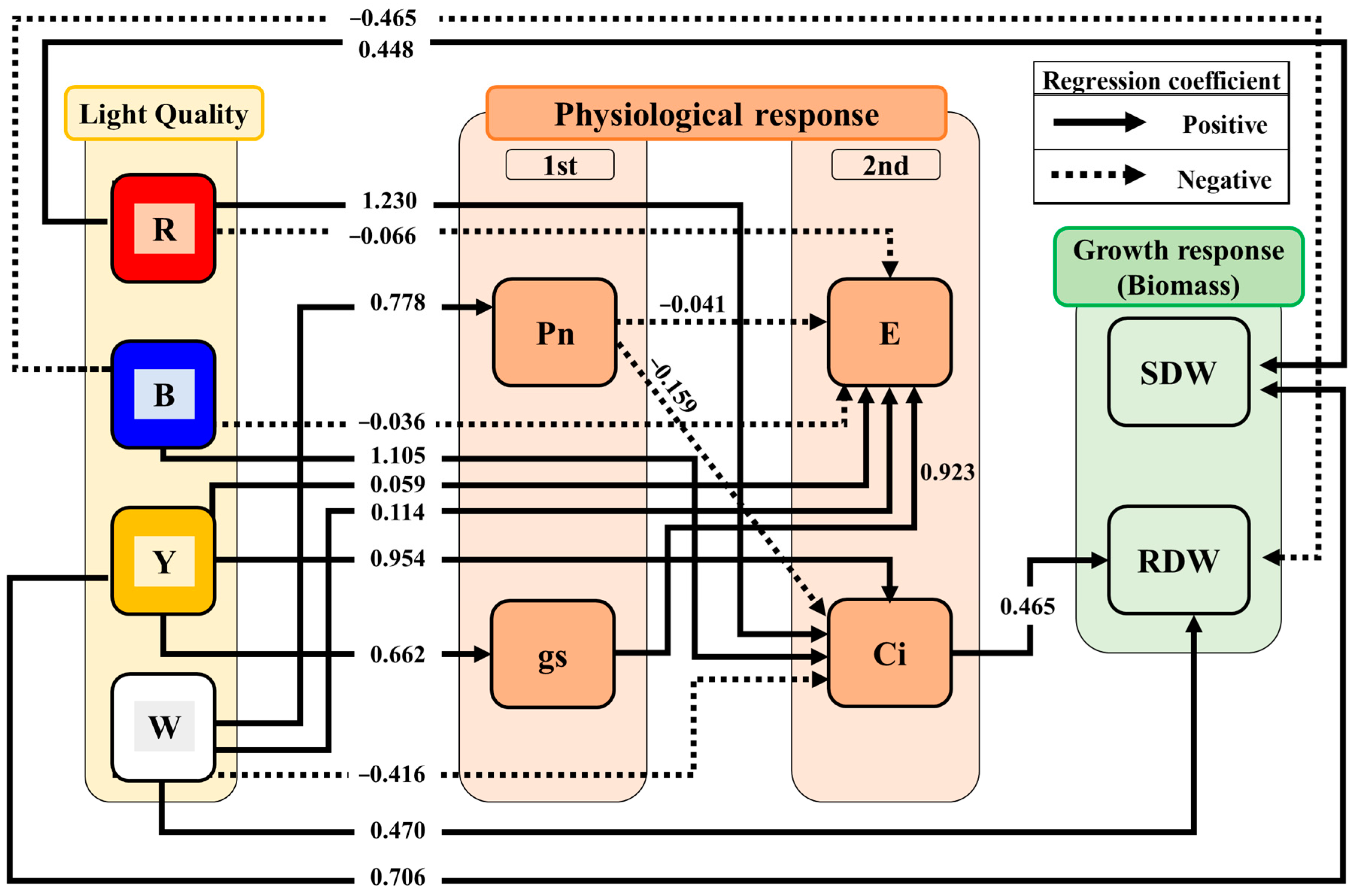
3.3. Pathways Affecting Physiological and Growth Responses
4. Discussion
4.1. The Role of Light Spectra in Physiological Responses
4.2. The Role of Light Spectra in Growth Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Neff, M.M.; Chory, J. Genetic interactions between phytochrome A, phytochrome B, and cryptochrome 1 during Arabidopsis development. Plant Physiol. 1998, 118, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Christie, J.M.; Blackwood, L.; Petersen, J.; Sullivan, S. Plant flavoprotein photoreceptors. Plant Cell Physiol. 2015, 56, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Demarsy, E.; Fankhauser, C. Higher plants use LOV to perceive blue light. Curr. Opin. Plant Biol. 2009, 12, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Nájera, C.; Gallegos-Cedillo, V.M.; Ros, M.; Pascual, J.A. Role of Spectrum-Light on Productivity, and Plant Quality over Vertical Farming Systems: Bibliometric Analysis. Horticulturae 2023, 9, 63. [Google Scholar] [CrossRef]
- Folta, K.M.; Maruhnich, S.A. Green light: A signal to slow down or stop. J. Exp. Bot. 2007, 58, 3099–3111. [Google Scholar] [CrossRef]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Good Information Company. Future New Growth Industry: Trends in Technology Development and Application Cases for Core Areas of Smart Agriculture/Smart Farming; Good Information Company: Hwaseong-si, Republic of Korea, 2022; pp. 250–251. [Google Scholar]
- Dorais, M. The use of supplemental lighting for vegetable crop production: Light intensity, crop response, nutrition, crop management, cultural practices. In Proceedings of the Canadian Greenhouse Conference, Toronto, ON, Canada, 9 October 2003; Available online: https://www.agrireseau.net/legumesdeserre/documents/cgc-dorais2003fin2.pdf (accessed on 8 May 2023).
- Kozai, T.; Niu, G.; Takagaki, M. Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production; Academic Press: London, UK, 2020. [Google Scholar] [CrossRef]
- Van Delden, S.H.; SharathKumar, M.; Butturini, M.; Graamans, L.J.A.; Heuvelink, E.; Kacira, M.; Kaiser, E.; Klamer, R.S.; Klerkx, L.; Kootstra, G.; et al. Current status and future challenges in implementing and upscaling vertical farming systems. Nat. Food 2021, 2, 944–956. [Google Scholar] [CrossRef]
- Barbosa, G.L.; Almeida Gadelha, F.D.; Kublik, N.; Proctor, A.; Reichelm, L.; Weissinger, E.; Wohlleb, G.M.; Halden, R.U. Comparison of land, water, and energy requirements of lettuce grown using hydroponic vs. conventional agricultural methods. Int. J. Environ Res. Public Health 2015, 12, 6879–6891. [Google Scholar] [CrossRef]
- Graamans, L.; Baeza, E.; Van Den Dobbelsteen, A.; Tsafaras, I.; Stanghellini, C. Plant factories versus greenhouses: Comparison of resource use efficiency. Agric. Syst. 2018, 160, 31–43. [Google Scholar] [CrossRef]
- Yoon, C.G.; Choi, H.K. A study on the various light source radiation conditions and use of LED illumination for plant factory. KIIEE 2011, 25, 14–22. [Google Scholar] [CrossRef]
- Kozai, T.; Niu, G. Plant Factory as a Resource-Efficient Closed Plant Production System; Academic Press: London, UK, 2016; pp. 387–393. [Google Scholar] [CrossRef]
- Seo, S.M. Eco-Physiological Characteristics and Ingredient Differences of Forest Ginseng in Various Forest Environments. Ph.D. Thesis, University of Seoul, Seoul, Republic of Korean, 2010. [Google Scholar]
- Park, C.K.; Jeon, B.S.; Yang, J.W. The chemical components of korean ginseng. Food Ind. Nutr. 2003, 8, 10–23. [Google Scholar]
- Han, J.H.; Park, S.J.; Ahn, C.N.; Wee, J.J.; Kim, K.Y.; Park, S.H. Nutritional Composition, Ginsenoside Content and Fundermental Safety Evaluation with Leaf and Stem Extract of Panax ginseng. J. Korean Soc. Food Sci. Nutr. 2004, 33, 778–784. [Google Scholar] [CrossRef]
- Hwang, S.H.; Kim, S.C.; Seong, J.A.; Lee, H.Y.; Cho, D.Y.; Kim, M.J.; Jung, J.G.; Jeong, E.H.; Son, K.H.; Cho, K.M. Comparison of ginsenoside contents and antioxidant activity according to the size of ginseng sprout has produced in a plant factory. J. Appl. Biol. Chem. 2021, 64, 253–261. [Google Scholar] [CrossRef]
- Baeg, I.H.; So, S.H. The world ginseng market and the ginseng (Korea). J. Ginseng Res. 2013, 37, 1–7. [Google Scholar] [CrossRef]
- Shin, S.M.; Park, M.S.; Lee, H.S.; Lee, S.G.; Lee, H.U.; Kim, T.H.; Kim, H.J. Global Trends in Research on Wild-Simulated Ginseng: Quo Vadis? Forests 2021, 12, 664. [Google Scholar] [CrossRef]
- Souther, S.; McGraw, J.B. Vulnerability of wild American ginseng to an extreme early spring temperature fluctuation. Popul. Ecol. 2011, 53, 119–129. [Google Scholar] [CrossRef]
- Hyun, D.Y.; Hwang, J.K.; Choi, S.Y.; Jo, J.S. Photosynthetic Characteristics of Panax ginseng C. A. Meyer-I. Photosynthetic Response to Changes of Light Intensity and Leaf Temperature. J. Ginseng Res. 1993, 17, 240–245. [Google Scholar]
- Hyun, D.Y.; You, N.H. Effect of leaf temperature on light response and respiration in Panax ginseng C. A. Meyer. KJCS 1996, 41, 725–728. [Google Scholar]
- Jang, I.B.; Do, G.R.; Hwang, H.S.; Suh, S.J.; Yu, J.; Jang, I.B.; Moon, J.W.; Chun, C.H. Morphological development and photosynthetic acclimation of Panax ginseng seedlings to irradiation by light-emitting diodes (LEDs). Hortic. Environ. Biotechnol. 2021, 62, 571–579. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Pearcy, R.W. The importance of sunflecks for forest understory plants. BioScience 1991, 41, 760–766. [Google Scholar] [CrossRef]
- Marcelis, L.F.M.; Heuvelink, E.; Goudriaan, J. Modelling biomass production and yield of horticultural crops: A review. Sci. Hortic. 1998, 74, 83–111. [Google Scholar] [CrossRef]
- Yeochon Association for Ecological Research. Modern Ecology Experiment; Kyomunsa: Seoul, Republic of Korea, 2005; pp. 264–268. [Google Scholar]
- Hu, L.T.; Bentler, P.M. Cutoff criteria for fit indexes in covariance structure analysis: Conventional criteria versus new alternatives. Struct. Equ. Model. 1999, 6, 1–55. [Google Scholar] [CrossRef]
- Smith, M.S.; McMillan, B.F. A Primer of Model Fit Indices in Structural Equation Modeling; Southwest Educational Research Association: New Orleans, LA, USA, 2001. [Google Scholar]
- West, S.G.; Taylor, A.B.; Wu, W. Model fit and model selection in structural equation modeling. Handb. Struct. Equ. Model. 2012, 1, 209–231. [Google Scholar]
- Zhou, C.; Zhang, Y.; Liu, W.; Zha, L.; Shao, M.; Li, B. Light quality affected the growth and root organic carbon and autotoxin secretions of hydroponic lettuce. Plants 2020, 9, 1542. [Google Scholar] [CrossRef]
- Meinzer, F.C. Stomatal control of transpiration. Trends Ecol. Evol. 1993, 8, 289–294. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Huang, S.; Ding, M.; Roelfsema, M.R.G.; Dreyer, I.; Scherzer, S.; Al-Rasheid, K.A.; Gao, S.; Nagel, G.; Hedrich, R.; Konrad, K.R. Optogenetic control of the guard cell membrane potential and stomatal movement by the light-gated anion channel Gt ACR1. Sci. Adv. 2021, 7, eabg4619. [Google Scholar] [CrossRef]
- Yavari, N.; Tripathi, R.; Wu, B.S.; MacPherson, S.; Singh, J.; Lefsrud, M. The effect of light quality on plant physiology, photosynthetic, and stress response in Arabidopsis thaliana leaves. PLoS ONE 2021, 16, e0247380. [Google Scholar] [CrossRef]
- Noe, S.M.; Giersch, C. A simple dynamic model of photosynthesis in oak leaves: Coupling leaf conductance and photosynthetic carbon fixation by a variable intracellular CO2 pool. Funct. Plant Biol. 2004, 31, 1195–1204. [Google Scholar] [CrossRef]
- Kong, S.W.; Chung, H.Y.; Chang, M.Y.; Fang, W. The contribution of different spectral sections to increase fresh weight of boston lettuce. HortScience 2015, 50, 1006–1010. [Google Scholar] [CrossRef]
- Oh, H.E.; Yoon, A.; Park, Y.G. Red Light enhances the antioxidant properties and growth of Rubus hongnoensis. Plants 2021, 10, 2589. [Google Scholar] [CrossRef]
- Mortensen, L.M.; Strømme, E. Effects of light quality on some greenhouse crops. Sci. Hortic. 1987, 33, 27–36. [Google Scholar] [CrossRef]
- Huston, M.; Smith, T. Plant succession: Life history and competition. Am. Nat. 1987, 130, 168–198. [Google Scholar] [CrossRef]
- Wei, H.; Chen, X.; Chen, G.; Zhao, H. Foliar nutrient and carbohydrate in Aralia elata can be modified by understory light quality in forests with different structures at Northeast China. Ann For Res. 2019, 62, 125–137. [Google Scholar] [CrossRef]
- Givnish, T.J. Adaptation to sun and shade: A whole-plant perspective. Funct. Plant Biol. 1988, 15, 63–92. [Google Scholar] [CrossRef]
- Tilman, D. Plant Strategies and the Dynamics and Structure of Plant Communities; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Wu, B.S.; Mansoori, M.; Trumpler, K.; Addo, P.W.; MacPherson, S.; Lefsrud, M. Effect of amber (595 nm) light supplemented with narrow blue (430 nm) light on tomato biomass. Plants 2023, 12, 2457. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, J.M.; Kim, E.J.; You, Y.H. The effects of LED light quality on ecophysiological and growth responses of Epilobium hirsutum L., a Korean endangered plant, in a smart farm facility. J. Ecol. Environ. 2022, 46, 16. [Google Scholar] [CrossRef]
- Kuronuma, T.; WANG, Q.; Ando, M.; Watanabe, H. Effects of Different Light Intensities on the Growth and Accumulation of Photosynthetic Products in Panax ginseng C. A. Meyers. Environ. Control Biol. 2020, 58, 131–135. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Description | Units |
|---|---|---|
| R | Red light | μmol m−2 s−1 |
| B | Blue light | μmol m−2 s−1 |
| Y | Yellow light | μmol m−2 s−1 |
| W | White light | μmol m−2 s−1 |
| Pn | Net photosynthetic rate | μmol m−2 s−1 |
| E | Transpiration rate | μmol m−2 s−1 |
| Ci | Intercellular CO2 partial pressure | vpm |
| gs | Stomatal conductance | mol m-m−2 s−1 |
| SDW | Shoot dry weight | g |
| RDW | Root dry weight | g |
| S/R ratio | Shoot dry weight to root dry weight ratio | - |
| Light Source | Spectral Ratio (%) | PPFD (μmol m−2 s−1) | |||
|---|---|---|---|---|---|
| Blue (400–500 nm) | Green (500–560 nm) | Yellow (560–630 nm) | Red (630–700 nm) | ||
| RB | 29.9 | 0.4 | 4.1 | 65.6 | 98.8 ± 9.3 |
| RBW | 19.7 | 9.5 | 16.6 | 54.2 | 115.3 ± 12.6 |
| B | 98.5 | 1 | 0.2 | 0.3 | 112.8 ± 22.4 |
| Y | 0.5 | 1.2 | 97.4 | 0.9 | 27.7 ± 4.0 |
| R | 0.3 | 0.2 | 4.8 | 94.7 | 114.4 ± 20.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-H.; Park, J.-H.; Kim, E.-J.; Lee, J.-M.; Park, J.-W.; Kim, Y.-S.; Kim, G.-R.; Lee, J.-S.; Lee, E.-P.; You, Y.-H. White LED Lighting Increases the Root Productivity of Panax ginseng C. A. Meyer in a Hydroponic Cultivation System of a Plant Factory. Biology 2023, 12, 1052. https://doi.org/10.3390/biology12081052
Kim S-H, Park J-H, Kim E-J, Lee J-M, Park J-W, Kim Y-S, Kim G-R, Lee J-S, Lee E-P, You Y-H. White LED Lighting Increases the Root Productivity of Panax ginseng C. A. Meyer in a Hydroponic Cultivation System of a Plant Factory. Biology. 2023; 12(8):1052. https://doi.org/10.3390/biology12081052
Chicago/Turabian StyleKim, Se-Hee, Jae-Hoon Park, Eui-Joo Kim, Jung-Min Lee, Ji-Won Park, Yoon-Seo Kim, Gyu-Ri Kim, Ju-Seon Lee, Eung-Pill Lee, and Young-Han You. 2023. "White LED Lighting Increases the Root Productivity of Panax ginseng C. A. Meyer in a Hydroponic Cultivation System of a Plant Factory" Biology 12, no. 8: 1052. https://doi.org/10.3390/biology12081052
APA StyleKim, S.-H., Park, J.-H., Kim, E.-J., Lee, J.-M., Park, J.-W., Kim, Y.-S., Kim, G.-R., Lee, J.-S., Lee, E.-P., & You, Y.-H. (2023). White LED Lighting Increases the Root Productivity of Panax ginseng C. A. Meyer in a Hydroponic Cultivation System of a Plant Factory. Biology, 12(8), 1052. https://doi.org/10.3390/biology12081052