
Role of the Alteration in Calcium Homeostasis in Cell Death Induced by Clostridioides difficile Toxin A and Toxin B

Abstract
:Simple Summary
Abstract
1. Introduction
2. C. difficile
2.1. General Characteristics of CDI
2.2. Molecular Structure of Tcds
2.3. Tcd Receptors
2.4. Cellular Uptake of Tcds and Tcd-Induced Cellular Effects
3. Mechanisms of Tcd-Induced Cytotoxicity
4. Alteration in Ca2+ Homeostasis by TcdA and TcdB
5. Comparison between PFT and Tcd Properties
5.1. Formation of Pore/Ca2+ Channels on the Plasma Membrane
5.2. Activation of Ca2+ Channels on the Plasma Membrane
5.3. Activation of Ca2+ Channels in the Endoplasmic Reticulum
5.4. Activation of Ca2+ Channels in Lysosomes
5.5. Activation of Ca2+-Dependent Cytoplasmic Proteins
5.6. Alteration in Ca2+ Homeostasis and Cell Death
5.6.1. Necrosis
5.6.2. Apoptosis
5.6.3. Pyroptosis
5.6.4. Comparison between Aerolysin and Tcds
6. Repair Mechanisms of the Influx of Ca2+
- Ectocytosis: this is a process of elimination towards the outside of the membrane areas that contain the pores by formation of microvesicles, and it takes place by two modalities [114,115,116]. The first is based on the activity of the annexins, which—activated by the high concentration of cytosolic Ca2+—migrate to the plasma membrane where the pores have formed [117]. Here, annexins establish a firm bond with Ca2+, contributing to the formation of a strong interaction with the plasma membrane. The latter forms membrane folds that are subsequently expelled, detaching from the membrane, which then rapidly reseals the externalized microarea called “microvesicles (MVs)” [117,118,119]. The second modality is realized through the activity of the endosomal sorting complex required for transport (ESCRT) [177,178].
- Multivesicular bodies: this is one of the membrane repair mechanisms that provokes a profound alteration in Ca2+ homeostasis, with an increase in the cytoplasmic concentration creating MVs that have two main purposes [118,119]. The first is to seal the parts of the membrane in which the pores have formed, mainly responsible for the influx of Ca2+ by ectocytosis; the second is to surround the cell with MVs derived from the membrane containing the receptors for the PFTs. Therefore, the PFTs are bound before they can reach the cell membrane. Despite the fact that Tcds have been convincingly shown to alter Ca2+ homeostasis, there are no direct studies on MV formation in response to Tcds. However, an interesting study on platelets demonstrated that a pathway that regulates the formation of MVs is blocked by TcdB [179]. This could explain why Tcds, despite the induction of Ca2+ increase, at the same time prevent the cell to form MVs as a protective response. In particular, the induction of a Ca2+ influx in platelets leads to the formation of MVs through the Rac1 and p21-activated kinase 1/2 (PAK1/2) activation pathway with the involvement of the activation of calpains, which in turn activate caspases [179]. By inhibiting Rac1, TcdB blocks the formation of MVs. Therefore, if TcdB induces Ca2+ influx, the cell’s response should be to generate MVs, but this cannot happen because TcdB inhibits Rac1 [179].
- This aspect is relevant for Tcd pathogenesis, because if the MV response also has a protective role [179], TcdB could have chosen a very sophisticated strategy to prevent the cell from protecting itself from TcdB by MVs.
7. Destruction of Intercellular Junctions (IJs)
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oren, A.; Rupnik, M. Clostridium difficile and Clostridioides difficile: Two validly published and correct names. Anaerobe 2018, 52, 125–126. [Google Scholar] [CrossRef]
- Lawson, P.A.; Citron, D.M.; Tyrrell, K.L.; Finegold, S.M. Reclassification of Clostridium difficile as Clostridioides difficile (Hall and O’Toole 1935) Prévot 1938. Anaerobe 2016, 40, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Balsells, E.; Shi, T.; Leese, C.; Lyell, I.; Burrows, J.; Wiuff, C.; Campbell, H.; Kyaw, M.H.; Nair, H. Global burden of Clostridium difficile infections: A systematic review and meta-analysis. J. Glob. Health 2019, 9, 010407. [Google Scholar] [CrossRef] [PubMed]
- Magill, S.S.; O’Leary, E.; Janelle, S.J.; Thompson, D.L.; Dumyati, G.; Nadle, J. Emerging Infections Program Hospital Prevalence Survey Team. Changes in Prevalence of Health Care–Associated Infections in U.S. Hospitals. N. Engl. J. Med. 2018, 379, 1732–1744. [Google Scholar] [CrossRef] [PubMed]
- Finn, E.; Andersson, F.L.; Madin-Warburton, M. Burden of Clostridioides difficile infection (CDI)—A systematic review of the epidemiology of primary and recurrent CDI. BMC Infect. Dis. 2021, 21, 456. [Google Scholar] [CrossRef] [PubMed]
- Guh, A.Y.; Kutty, P.K. Clostridioides difficile Infection. Ann. Intern. Med. 2018, 169, ITC49–ITC64. [Google Scholar] [CrossRef]
- Czepiel, J.; Dróżdż, M.; Pituch, H.; Kuijper, E.J.; Perucki, W.; Mielimonka, A.; Goldman, S.; Wultańska, D.; Garlicki, A.; Biesiada, G. Clostridium difficile infection: Review. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Aktories, K.; Schwan, C.; Jank, T. Clostridium difficile Toxin Biology. Annu. Rev. Microbiol. 2017, 71, 281–307. [Google Scholar] [CrossRef]
- Chandrasekaran, R.; Lacy, D.B. The role of toxins in Clostridium difficile infection. FEMS Microbiol. Rev. 2017, 41, 723–750. [Google Scholar] [CrossRef] [Green Version]
- Di Bella, S.; Ascenzi, P.; Siarakas, S.; Petrosillo, N.; di Masi, A. Clostridium difficile Toxins A and B: Insights into Pathogenic Properties and Extraintestinal Effects. Toxins 2016, 8, 134. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Savidge, T.; Feng, H. The Enterotoxicity of Clostridium difficile Toxins. Toxins 2010, 2, 1848–1880. [Google Scholar] [CrossRef] [Green Version]
- Pruitt, R.N.; Lacy, D.B. Toward a structural understanding of Clostridium difficile toxins A and B. Front. Cell. Infect. Microbiol. 2012, 2, 28. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Luo, Y.; Grinspan, A.M. Epidemiology of community-acquired and recurrent Clostridioides difficile infection. Therap. Adv. Gastroenterol. 2021, 14, 175628482110162. [Google Scholar] [CrossRef]
- Boeriu, A.; Roman, A.; Fofiu, C.; Dobru, D. The Current Knowledge on Clostridioides difficile Infection in Patients with Inflammatory Bowel Diseases. Pathogens 2022, 11, 819. [Google Scholar] [CrossRef]
- Bassotti, G.; Fruganti, A.; Maconi, G.; Marconi, P.; Fettucciari, K. Clostridioides difficile Infection in Patients with Inflammatory Bowel Disease May be Favoured by the Effects of Proinflammatory Cytokines on the Enteroglial Network. J. Inflamm. Res. 2021, 14, 7443–7453. [Google Scholar] [CrossRef]
- Dalal, R.S.; Allegretti, J.R. Diagnosis and management of Clostridioides difficile infection in patients with inflammatory bowel disease. Curr. Opin. Gastroenterol. 2021, 37, 336–343. [Google Scholar] [CrossRef]
- Khanna, S. Management of Clostridioides difficile infection in patients with inflammatory bowel disease. Intest. Res. 2021, 19, 265–274. [Google Scholar] [CrossRef]
- Badilla-Lobo, A.; Rodríguez, C. Microbiological features, epidemiology, and clinical presentation of Clostridioides difficile strains from MLST Clade 2: A narrative review. Anaerobe 2021, 69, 102355. [Google Scholar] [CrossRef]
- Hunt, J.J.; Ballard, J.D. Variations in Virulence and Molecular Biology among Emerging Strains of Clostridium difficile. Microbiol. Mol. Biol. Rev. 2013, 77, 567–581. [Google Scholar] [CrossRef] [Green Version]
- Kachrimanidou, M.; Tzika, E.; Filioussis, G. Clostridioides (Clostridium) difficile in food-producing animals, horses and household pets: A comprehensive review. Microorganisms 2019, 7, 667. [Google Scholar] [CrossRef] [Green Version]
- Fettucciari, K.; Marconi, P.; Marchegiani, A.; Fruganti, A.; Spaterna, A.; Bassotti, G. Invisible steps for a global endemy: Molecular strategies adopted by Clostridioides difficile. Therap. Adv. Gastroenterol. 2021, 14, 175628482110327. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.C.; Knight, D.R.; Riley, T.V. Clostridium difficile and One Health. Clin. Microbiol. Infect. 2020, 26, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Comparcini, D.; Simonetti, V.; Segala, F.V.; Di Gennaro, F.; Bavaro, D.F.; Pompeo, M.A.; Saracino, A.; Cicolini, G. Nurses’ Knowledge, Attitudes and Practices on the Management of Clostridioides difficile Infection: A Cross-Sectional Study. Antibiotics 2023, 12, 529. [Google Scholar] [CrossRef] [PubMed]
- Raeisi, H.; Azimirad, M.; Nabavi-Rad, A.; Asadzadeh Aghdaei, H.; Yadegar, A.; Zali, M.R. Application of recombinant antibodies for treatment of Clostridioides difficile infection: Current status and future perspective. Front. Immunol. 2022, 13, 972930. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, D.P.; Wilcox, M.H. Antibodies for treatment of Clostridium difficile infection. Clin. Vaccine Immunol. 2014, 21, 913–923. [Google Scholar] [CrossRef]
- Razim, A.; Górska, S.; Gamian, A. Non-Toxin-Based Clostridioides difficile Vaccination Approaches. Pathogens 2023, 12, 235. [Google Scholar] [CrossRef]
- Leuzzi, R.; Adamo, R.; Scarselli, M. Vaccines against Clostridium difficile comprising recombinant toxins. Hum. Vaccin. Immunother. 2014, 10, 1466–1477. [Google Scholar] [CrossRef] [Green Version]
- Bakken, J.S.; Borody, T.; Brandt, L.J.; Brill, J.V.; Demarco, D.C.; Franzos, M.A.; Kelly, C.; Khoruts, A.; Louie, T.; Martinelli, L.P.; et al. Treating Clostridium difficile infection with fecal microbiota transplantation. Clin. Gastroenterol. Hepatol. 2011, 9, 1044–1049. [Google Scholar] [CrossRef] [Green Version]
- Baunwall, S.M.D.; Lee, M.M.; Eriksen, M.K.; Mullish, B.H.; Marchesi, J.R.; Dahlerup, J.F.; Hvas, C.L. Faecal microbiota transplantation for recurrent Clostridioides difficile infection: An updated systematic review and meta-analysis. EClinicalMedicine 2020, 29–30, 100642. [Google Scholar] [CrossRef]
- Liubakka, A.; Vaughn, B.P. Clostridium difficile infection and fecal microbiota transplant. AACN Adv. Crit. Care 2016, 27, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Weiss, K. Toxin-binding treatment for Clostridium difficile: A review including reports of studies with tolevamer. Int. J. Antimicrob. Agents 2009, 33, 4–7. [Google Scholar] [CrossRef]
- Le Lay, C.; Dridi, L.; Bergeron, M.G.; Ouellette, M.; Fliss, I. Nisin is an effective inhibitor of Clostridium difficile vegetative cells and spore germination. J. Med. Microbiol. 2016, 65, 169–175. [Google Scholar] [CrossRef]
- Sorg, J.A.; Sonenshein, A.L. Inhibiting the initiation of Clostridium difficile spore germination using analogs of chenodeoxycholic acid, a bile acid. J. Bacteriol. 2010, 192, 4983–4990. [Google Scholar] [CrossRef] [Green Version]
- Gerding, D.N.; Sambol, S.P.; Johnson, S. Non-Toxigenic Clostridioides (Formerly Clostridium) difficile for Prevention of C. difficile Infection: From Bench to Bedside Back to Bench and Back to Bedside. Front. Microbiol. 2018, 9, 1700. [Google Scholar] [CrossRef]
- Heuler, J.; Fortier, L.C.; Sun, X. Clostridioides difficile phage biology and application. FEMS Microbiol. Rev. 2021, 45, fuab012. [Google Scholar] [CrossRef]
- Nale, J.Y.; Thanki, A.M.; Rashid, S.J.; Shan, J.; Vinner, G.K.; Dowah, A.S.A.; Cheng, J.K.J.; Sicheritz-Pontén, T.; Clokie, M.R.J. Diversity, Dynamics and Therapeutic Application of Clostridioides difficile Bacteriophages. Viruses 2022, 14, 2772. [Google Scholar] [CrossRef]
- Ivarsson, M.E.; Durantie, E.; Huberli, C.; Huwiler, S.; Hegde, C.; Friedman, J.; Altamura, F.; Lu, J.; Verdu, E.F.; Bercik, P.; et al. Small-Molecule Allosteric Triggers of Clostridium difficile Toxin B Auto-proteolysis as a Therapeutic Strategy. Cell Chem. Biol. 2019, 26, 17–26.e13. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yang, H. Non-antibiotic therapy for Clostridioides difficile infection: A review. Crit. Rev. Clin. Lab. Sci. 2019, 56, 493–509. [Google Scholar] [CrossRef]
- Jarmo, O.; Veli-Jukka, A.; Eero, M. Treatment of Clostridioides (Clostridium) difficile infection. Ann. Med. 2020, 52, 12–20. [Google Scholar] [CrossRef]
- Bainum, T.B.; Reveles, K.R.; Hall, R.G.; Cornell, K.; Alvarez, C.A. Controversies in the Prevention and Treatment of Clostridioides difficile Infection in Adults: A Narrative Review. Microorganisms 2023, 11, 387. [Google Scholar] [CrossRef]
- Singh, T.; Bedi, P.; Bumrah, K.; Singh, J.; Rai, M.; Seelam, S. Updates in Treatment of Recurrent Clostridium difficile Infection. J. Clin. Med. Res. 2019, 11, 465–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Córdova, P.; Mora-Uribe, P.; Reyes-Ramírez, R.; Cofré-Araneda, G.; Orozco-Aguilar, J.; Brito-Silva, C.; Mendoza-León, M.J.; Kuehne, S.A.; Minton, N.P.; Pizarro-Guajardo, M.; et al. Entry of spores into intestinal epithelial cells contributes to recurrence of Clostridioides difficile infection. Nat. Commun. 2021, 12, 1140. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Sabja, D.; Shen, A.; Sorg, J.A. Clostridium difficile spore biology: Sporulation, germination, and spore structural proteins. Trends Microbiol. 2014, 22, 406–416. [Google Scholar] [CrossRef] [Green Version]
- Kirk, J.A.; Banerji, O.; Fagan, R.P. Characteristics of the Clostridium difficile cell envelope and its importance in therapeutics. Microb. Biotechnol. 2017, 10, 76–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjuwon-Foster, B.R.; Tamayo, R. Phase variation of Clostridium difficile virulence factors. Gut Microbes 2018, 9, 76–83. [Google Scholar] [CrossRef] [Green Version]
- Fettucciari, K.; Ponsini, P.; Gioè, D.; Macchioni, L.; Palumbo, C.; Antonelli, E.; Coaccioli, S.; Villanacci, V.; Corazzi, L.; Marconi, P.; et al. Enteric glial cells are susceptible to Clostridium difficile toxin B. Cell. Mol. Life Sci. 2017, 74, 1527–1551. [Google Scholar] [CrossRef]
- Fettucciari, K.; Marguerie, F.; Fruganti, A.; Marchegiani, A.; Spaterna, A.; Brancorsini, S.; Marconi, P.; Bassotti, G. Clostridioides difficile toxin B alone and with pro-inflammatory cytokines induces apoptosis in enteric glial cells by activating three different signalling pathways mediated by caspases, calpains and cathepsin B. Cell. Mol. Life Sci. 2022, 79, 442. [Google Scholar] [CrossRef]
- Orrell, K.E.; Melnyk, R.A. Large Clostridial Toxins: Mechanisms and Roles in Disease. Microbiol. Mol. Biol. Rev. 2021, 85, e0006421. [Google Scholar] [CrossRef]
- Riegler, M.; Sedivy, R.; Pothoulakis, C.; Hamilton, G.; Zacherl, J.; Bischof, G.; Cosentini, E.; Feil, W.; Schiessel, R.; LaMont, J.T.; et al. Clostridium difficile toxin B is more potent than toxin A in damaging human colonic epithelium in vitro. J. Clin. Investig. 1995, 95, 2004–2011. [Google Scholar] [CrossRef] [Green Version]
- El Hadidy, N.; Uversky, V.N.; Sun, X. On the Potential Significance of the Intrinsically Disordered Regions in the Clostridiodes difficile Toxins A and B. Curr. Protein Pept. Sci. 2022, 23, 192–209. [Google Scholar] [CrossRef]
- Orrell, K.E.; Zhang, Z.; Sugiman-Marangos, S.N.; Melnyk, R.A. Clostridium difficile toxins A and B: Receptors, pores, and translocation into cells. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 461–473. [Google Scholar] [CrossRef]
- Pothoulakis, C.; Gilbert, R.J.; Cladaras, C.; Castagliuolo, I.; Semenza, G.; Hitti, Y.; Montcrief, J.S.; Linevsky, J.; Kelly, C.P.; Nikulasson, S.; et al. Rabbit sucrase-isomaltase contains a functional intestinal receptor for Clostridium difficile toxin A. J. Clin. Investig. 1996, 98, 641–649. [Google Scholar] [CrossRef] [Green Version]
- Na, X.; Kim, H.; Moyer, M.P.; Pothoulakis, C.; LaMont, J.T. gp96 is a human colonocyte plasma membrane binding protein for Clostridium difficile toxin A. Infect. Immun. 2008, 76, 2862–2871. [Google Scholar] [CrossRef] [Green Version]
- Yuan, P.; Zhang, H.; Cai, C.; Zhu, S.; Zhou, Y.; Yang, X.; He, R.; Li, C.; Guo, S.; Li, S.; et al. Chondroitin sulfate proteoglycan 4 functions as the cellular receptor for Clostridium difficile toxin B. Cell Res. 2015, 25, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Tao, L.; Zhang, J.; Meraner, P.; Tovaglieri, A.; Wu, X.; Gerhard, R.; Zhang, X.; Stallcup, W.B.; Miao, J.; He, X.; et al. Frizzled proteins are colonic epithelial receptors for C. difficile toxin B. Nature 2016, 538, 350–355. [Google Scholar] [CrossRef] [Green Version]
- LaFrance, M.E.; Farrow, M.A.; Chandrasekaran, R.; Sheng, J.; Rubin, D.H.; Lacy, D.B. Identification of an epithelial cell receptor responsible for Clostridium difficile TcdB-induced cytotoxicity. Proc. Natl. Acad. Sci. USA 2015, 112, 7073–7078. [Google Scholar] [CrossRef]
- Terada, N.; Ohno, N.; Murata, S.; Katoh, R.; Stallcup, W.B.; Ohno, S. Immunohistochemical study of NG2 chondroitin sulfate proteoglycan expression in the small and large intestines. Histochem. Cell. Biol. 2006, 126, 483–490. [Google Scholar] [CrossRef]
- Henkel, D.; Tatge, H.; Schöttelndreier, D.; Tao, L.; Dong, M.; Gerhard, R. Receptor Binding Domains of TcdB from Clostridioides difficile for Chondroitin Sulfate Proteoglycan-4 and Frizzled Proteins Are Functionally Independent and Additive. Toxins 2020, 12, 736. [Google Scholar] [CrossRef]
- Bassotti, G.; Fruganti, A.; Stracci, F.; Marconi, P.; Fettucciari, K. Cytotoxic synergism of Clostridioides difficile toxin B with proinflammatory cytokines in subjects with inflammatory bowel diseases. World J. Gastroenterol. 2023, 29, 582–596. [Google Scholar] [CrossRef]
- Fettucciari, K.; Fruganti, A.; Stracci, F.; Spaterna, A.; Marconi, P.; Bassotti, G. Clostridioides difficile Toxin B Induced Senescence: A New Pathologic Player for Colorectal Cancer? Int. J. Mol. Sci. 2023, 24, 8155. [Google Scholar] [CrossRef]
- D’Auria, K.M.; Bloom, M.J.; Reyes, Y.; Gray, M.C.; Van Opstal, E.J.; Papin, J.A.; Hewlett, E.L. High temporal resolution of glucosyltransferase dependent and independent effects of Clostridium difficile toxins across multiple cell types. BMC Microbiol. 2015, 15, 7. [Google Scholar] [CrossRef] [PubMed]
- Chumbler, N.M.; Farrow, M.A.; Lapierre, L.A.; Franklin, J.L.; Haslam, D.; Goldenring, J.R.; Lacy, D.B. Clostridium difficile Toxin B Causes Epithelial Cell Necrosis through an Autoprocessing-Independent Mechanism. PLoS Pathog. 2012, 8, 1003072. [Google Scholar] [CrossRef]
- Chumbler, N.M.; Farrow, M.A.; Lapierre, L.A.; Franklin, J.L.; Borden, D. Clostridium difficile Toxins TcdA and TcdB Cause Colonic Tissue. Infect. Immun. 2016, 84, 2871–2877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrow, M.A.; Chumbler, N.M.; Lapierre, L.A.; Franklin, J.L.; Rutherford, S.A.; Goldenring, J.R.; Lacy, D.B. Clostridium difficile toxin B-induced necrosis is mediated by the host epithelial cell NADPH oxidase complex. Proc. Natl. Acad. Sci. USA 2013, 110, 18674–18679. [Google Scholar] [CrossRef] [PubMed]
- Wohlan, K.; Goy, S.; Olling, A.; Srivaratharajan, S.; Tatge, H.; Genth, H.; Gerhard, R. Pyknotic cell death induced by Clostridium difficile TcdB: Chromatin condensation and nuclear blister are induced independently of the glucosyltransferase activity. Cell. Microbiol. 2014, 16, 1678–1692. [Google Scholar] [CrossRef]
- Brito, G.A.C.; Fujji, J.; Carneiro-Filho, B.A.; Lima, A.A.M.; Obrig, T.; Guerrant, R.L. Mechanism of Clostridium difficile toxin A-induced apoptosis in T84 cells. J. Infect. Dis. 2002, 186, 1438–1447. [Google Scholar] [CrossRef] [Green Version]
- Mullan, N.; Hughes, K.R.; Mahida, Y.R. Primary human colonic myofibroblasts are resistant to Clostridium difficile toxin A-induced, but not toxin B-induced, cell death. Infect. Immun. 2011, 79, 1623–1630. [Google Scholar] [CrossRef] [Green Version]
- Neunlist, M.; Barouk, J.; Michel, K.; Just, I.; Oreshkova, T.; Schemann, M.; Galmiche, J.P. Toxin B of Clostridium difficile activates human VIP submucosal neurons, in part via an IL-1β-dependent pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G1049–G1055. [Google Scholar] [CrossRef] [Green Version]
- Mahida, Y.R.; Galvin, A.; Makh, S.; Hyde, S.; Sanfilippo, L.; Borriello, S.P.; Sewell, H.F. Effect of Clostridium difficile toxin A on human colonic lamina propria cells: Early loss of macrophages followed by T-cell apoptosis. Infect. Immun. 1998, 66, 5462–5469. [Google Scholar] [CrossRef]
- Modi, N.; Gulati, N.; Solomon, K.; Monaghan, T.; Robins, A.; Sewell, H.F.; Mahida, Y.R. Differential binding and internalization of Clostridium difficile toxin A by human peripheral blood monocytes, neutrophils and lymphocytes. Scand. J. Immunol. 2011, 74, 264–271. [Google Scholar] [CrossRef]
- Solomon, K.; Webb, J.; Ali, N.; Robins, R.A.; Mahida, Y.R. Monocytes are highly sensitive to Clostridium difficile toxin A-induced apoptotic and nonapoptotic cell death. Infect. Immun. 2005, 73, 1625–1634. [Google Scholar] [CrossRef] [Green Version]
- Linseman, D.A.; Laessig, T.; Meintzer, M.K.; McClure, M.; Barth, H.; Aktories, K.; Heidenreich, K.A. An Essential Role for Rac/Cdc42 GTPases in Cerebellar Granule Neuron Survival. J. Biol. Chem. 2001, 276, 39123–39131. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Hong, J.; Yoon, I.N.; Kang, J.K.; Hwang, J.S.; Kim, H. Clostridium difficile toxin a induces reactive oxygen species production and p38 MAPK activation to exert cellular toxicity in neuronal cells. J. Microbiol. Biotechnol. 2017, 27, 1163–1170. [Google Scholar] [CrossRef] [Green Version]
- Florin, I. Isolation of a fibroblast mutant resistant to Clostridium difficile toxins A and B. Microb. Pathog. 1991, 11, 337–346. [Google Scholar] [CrossRef]
- Grossmann, E.M.; Longo, W.E.; Kaminski, D.L.; Smith, G.S.; Murphy, C.E.; Durham, R.L.; Shapiro, M.J.; Norman, J.G.; Mazuski, J.E. Clostridium difficile toxin: Cytoskeletal changes and lactate dehydrogenase release in hepatocytes. J. Surg. Res. 2000, 88, 165–172. [Google Scholar] [CrossRef]
- Krijnen, P.A.J.; Sipkens, J.A.; Molling, J.W.; Rauwerda, J.A.; Stehouwer, C.D.A.; Muller, A.; Paulus, W.J.; van Nieuw Amerongen, G.P.; Hack, C.E.; Verhoeven, A.J.; et al. Inhibition of Rho-ROCK signaling induces apoptotic and non-apoptotic PS exposure in cardiomyocytes via inhibition of flippase. J. Mol. Cell. Cardiol. 2010, 49, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.V.S.; Shin, J.H.; Goldbeck, S.M.; Bolick, D.T.; Mesquita, F.S.; Loureiro, A.V.; Rodrigues-Jesus, M.J.; Brito, G.A.C.; Warren, C.A. Adenosine receptors differentially mediate enteric glial cell death induced by Clostridioides difficile Toxins A and B. Front. Immunol. 2023, 13, 956326. [Google Scholar] [CrossRef]
- Costa, D.V.S.; Bon-Frauches, A.C.; Silva, A.M.H.P.; Lima-Júnior, R.C.P.; Martins, C.S.; Leitão, R.F.C.; Freitas, G.B.; Castelucci, P.; Bolick, D.T.; Guerrant, R.L.; et al. 5-Fluorouracil Induces Enteric Neuron Death and Glial Activation During Intestinal Mucositis via a S100B-RAGE-NFκB-Dependent Pathway. Sci. Rep. 2019, 9, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowardin, C.A.; Jackman, B.M.; Noor, Z.; Burgess, S.L.; Feig, A.L.; Petri, W.A. Glucosylation drives the innate inflammatory response to Clostridium difficile toxin A. Infect. Immun. 2016, 84, 2317–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fettucciari, K.; Macchioni, L.; Davidescu, M.; Scarpelli, P.; Palumbo, C.; Corazzi, L.; Marchegiani, A.; Cerquetella, M.; Spaterna, A.; Marconi, P.; et al. Clostridium difficile toxin B induces senescence in enteric glial cells: A potential new mechanism of Clostridium difficile pathogenesis. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1945–1958. [Google Scholar] [CrossRef]
- Christopher Peritore-Galve, F.; Shupe, J.A.; Cave, R.J.; Childress, K.O.; Kay Washington, M.; Kuehne, S.A.; Borden Lacy, D. Glucosyltransferase-dependent and independent effects of Clostridioides difficile toxins during infection. PLoS Pathog. 2022, 18, e1010323. [Google Scholar] [CrossRef]
- Farrow, M.A.; Chumber, N.M.; Bloch, S.C.; King, M.; Moton-Melancon, K.; Shupe, J.; Washington, M.K.; Spiller, B.W.; Lacy, D.B. Small Molecule Inhibitor Screen Reveals Calcium Channel Signaling as a Mechanistic Mediator of Clostridium difficile TcdB-Induced Necrosis. ACS Chem. Biol. 2020, 15, 1212–1221. [Google Scholar] [CrossRef]
- He, D.; Hagen, S.J.; Pothoulakis, C.; Chen, M.; Medina, N.D.; Warny, M.; LaMont, J.T. Clostridium difficile toxin A causes early damage to mitochondria in cultured cells. Gastroenterology 2000, 119, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kokkotou, E.; Na, X.; Rhee, S.H.; Moyer, M.P.; Pothoulakis, C.; Lamont, J.T. Clostridium difficile toxin A-induced colonocyte apoptosis involves p53-dependent p21(WAF1/CIP1) induction via p38 mitogen-activated protein kinase. Gastroenterology 2005, 129, 1875–1888. [Google Scholar] [CrossRef]
- Carneiro, B.A.; Fujii, J.; Brito, G.A.C.; Alcantara, C.; Oriá, R.B.; Lima, A.A.M.; Obrig, T.; Guerrant, R.L. Caspase and bid involvement in Clostridium difficile toxin A-induced apoptosis and modulation of toxin A effects by glutamine and alanyl-glutamine in vivo and in vitro. Infect. Immun. 2006, 74, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Nottrott, S.; Schoentaube, J.; Genth, H.; Just, I.; Gerhard, R. Clostridium difficile toxin A-induced apoptosis is p53-independent but depends on glucosylation of Rho GTPases. Apoptosis 2007, 12, 1443–1453. [Google Scholar] [CrossRef]
- Gerhard, R.; Nottrott, S.; Schoentaube, J.; Tatge, H.; Oiling, A.; Just, I. Glucosylation of Rho GTPases by Clostridium difficile toxin A triggers apoptosis in intestinal epithelial cells. J. Med. Microbiol. 2008, 57, 765–770. [Google Scholar] [CrossRef] [Green Version]
- Hippenstiel, S.; Schmeck, B.; N’Guessan, P.D.; Seybold, J.; Krüll, M.; Preissner, K.; Eichel-Streiber, C.V.; Suttorp, N. Rho protein inactivation induced apoptosis of cultured human endothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 283, L830–L838. [Google Scholar] [CrossRef]
- Qa’Dan, M.; Ramsey, M.; Daniel, J.; Spyres, L.M.; Safiejko-Mroczka, B.; Ortiz-Leduc, W.; Ballard, J.D. Clostridium difficile toxin B activates dual caspase-dependent and caspase-independent apoptosis in intoxicated cells. Cell. Microbiol. 2002, 4, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Le, S.S.; Loucks, F.A.; Udo, H.; Richardson-Burns, S.; Phelps, R.A.; Bouchard, R.J.; Barth, H.; Aktories, K.; Tyler, K.L.; Kandel, E.R.; et al. Inhibition of Rac GTPase triggers a c-Jun- and Bim-dependent mitochondrial apoptotic cascade in cerebellar granule neurons. J. Neurochem. 2005, 94, 1025–1039. [Google Scholar] [CrossRef] [Green Version]
- Matarrese, P.; Falzano, L.; Fabbri, A.; Gambardella, L.; Frank, C.; Geny, B.; Popoff, M.R.; Malorni, W.; Fiorentini, C. Clostridium difficile toxin B causes apoptosis in epithelial cells by thrilling mitochondria: Involvement of ATP-sensitive mitochondrial potassium channels. J. Biol. Chem. 2007, 282, 9029–9041. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.; Dreger, S.; Gerhard, R.; Barth, H.; Just, I.; Genth, H. Difference in the cytotoxic effects of toxin B from Clostridium difficile strain VPI 10463 and toxin B from variant Clostridium difficile strain 1470. Infect. Immun. 2007, 75, 801–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Park, M.; Tam, J.; Auger, A.; Beilhartz, G.L.; Lacy, D.B.; Melnyk, R.A. Translocation domain mutations affecting cellular toxicity identify the Clostridium difficile toxin B pore. Proc. Natl. Acad. Sci. USA 2014, 111, 3721–3726. [Google Scholar] [CrossRef] [PubMed]
- Matte, I.; Lane, D.; Côté, É.; Asselin, A.É.; Fortier, L.C.; Asselin, C.; Piché, A. Antiapoptotic proteins Bcl-2 and Bcl-XL inhibit Clostridium difficile toxin A-induced cell death in human epithelial cells. Infect. Immun. 2009, 77, 5400–5410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufmann, S.H.; Hengartner, M.O. Programmed cell death: Alive and well in the new millennium. Trends Cell Biol. 2001, 11, 526–534. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Bidère, N.; Senik, A. Caspase-independent apoptotic pathways in T lymphocytes: A minireview. Apoptosis 2001, 6, 371–375. [Google Scholar] [CrossRef]
- Jäättelä, M.; Tschopp, J. Caspase-independent cell death in T lymphocytes. Nat. Immunol. 2003, 4, 416–423. [Google Scholar] [CrossRef]
- Mathiasen, I.S.; Jäättelä, M. Triggering caspase-independent cell death to combat cancer. Trends Mol. Med. 2002, 8, 212–220. [Google Scholar] [CrossRef]
- Momeni, H.R. Role of calpain in apoptosis. Cell J. 2011, 13, 65–72. [Google Scholar]
- Aits, S.; Jäättelä, M. Lysosomal cell death at a glance. J. Cell Sci. 2013, 126, 1905–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macchioni, L.; Davidescu, M.; Fettucciari, K.; Petricciuolo, M.; Gatticchi, L.; Gioè, D.; Villanacci, V.; Bellini, M.; Marconi, P.; Roberti, R.; et al. Enteric glial cells counteract Clostridium difficile Toxin B through a NADPH oxidase/ROS/JNK/caspase-3 axis, without involving mitochondrial pathways. Sci. Rep. 2017, 7, 45569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frädrich, C.; Beer, L.A.; Gerhard, R. Reactive oxygen species as additional determinants for cytotoxicity of Clostridium difficile toxins A and B. Toxins 2016, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stieglitz, F.; Gerhard, R.; Hönig, R.; Giehl, K.; Pich, A. TcdB of Clostridioides difficile Mediates RAS-Dependent Necrosis in Epithelial Cells. Int. J. Mol. Sci. 2022, 23, 4258. [Google Scholar] [CrossRef] [PubMed]
- Warny, M.; Kelly, C.P. Monocytic cell necrosis is mediated by potassium depletion and caspase- like proteases. Am. J. Physiol. Cell Physiol. 1999, 276, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Donald, R.G.K.; Flint, M.; Kalyan, N.; Johnson, E.; Witko, S.E.; Kotash, C.; Zhao, P.; Megati, S.; Yurgelonis, I.; Lee, P.K.; et al. A novel approach to generate a recombinant toxoid vaccine against Clostridium difficile. Microbiology 2013, 159, 1254–1266. [Google Scholar] [CrossRef]
- Ng, J.; Hirota, S.A.; Gross, O.; Li, Y.; Ulke-Lemee, A.; Potentier, M.S.; Schenck, L.P.; Vilaysane, A.; Seamone, M.E.; Feng, H.; et al. Clostridium difficile toxin-induced inflammation and intestinal injury are mediated by the inflammasome. Gastroenterology 2010, 139, 542–552.e3. [Google Scholar] [CrossRef]
- Liu, Y.H.; Chang, Y.C.; Chen, L.K.; Su, P.A.; Ko, W.C.; Tsai, Y.S.; Chen, Y.H.; Lai, H.C.; Wu, C.Y.; Hung, Y.P.; et al. The ATP-P2X7 signaling axis is an essential sentinel for intracellular Clostridium difficile pathogen-induced inflammasome activation. Front. Cell. Infect. Microbiol. 2018, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Gao, W.; Shao, F. Pyroptosis: Gasdermin-Mediated Programmed Necrotic Cell Death. Trends Biochem. Sci. 2017, 42, 245–254. [Google Scholar] [CrossRef]
- Xu, H.; Yang, J.; Gao, W.; Li, L.; Li, P.; Zhang, L.; Gong, Y.N.; Peng, X.; Xi, J.J.; Chen, S.; et al. Innate immune sensing of bacterial modifications of Rho GTPases by the Pyrin inflammasome. Nature 2014, 513, 237–241. [Google Scholar] [CrossRef]
- Russo, H.M.; Rathkey, J.; Boyd-Tressler, A.; Katsnelson, M.A.; Abbott, D.W.; Dubyak, G.R. Active Caspase-1 Induces Plasma Membrane Pores That Precede Pyroptotic Lysis and Are Blocked by Lanthanides. J. Immunol. 2016, 197, 1353–1367. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Ockinger, J.; Yu, J.; Byles, V.; McColl, A.; Hofer, A.M.; Horng, T. Critical role for calcium mobilization in activation of the NLRP3 inflammasome. Proc. Natl. Acad. Sci. USA 2012, 109, 11282–11287. [Google Scholar] [CrossRef] [PubMed]
- Peraro, M.D.; Van Der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef]
- Bouillot, S.; Reboud, E.; Huber, P. Functional consequences of calcium influx promoted by bacterial pore-forming Toxins. Toxins 2018, 10, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran Van Nhieu, G.; Dupont, G.; Combettes, L. Ca2+ signals triggered by bacterial pathogens and microdomains. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1838–1845. [Google Scholar] [CrossRef] [PubMed]
- Babiychuk, E.B.; Draeger, A. Defying death: Cellular survival strategies following plasmalemmal injury by bacterial toxins. Semin. Cell Dev. Biol. 2015, 45, 39–47. [Google Scholar] [CrossRef]
- Koerdt, S.N.; Ashraf, A.P.K.; Gerke, V. Annexins and plasma membrane repair. Curr. Top. Membr. 2019, 84, 43–65. [Google Scholar] [CrossRef] [PubMed]
- Latifkar, A.; Hur, Y.H.; Sanchez, J.C.; Cerione, R.A.; Antonyak, M.A. New insights into extracellular vesicle biogenesis and function. J. Cell Sci. 2019, 132, jcs222406. [Google Scholar] [CrossRef] [Green Version]
- Panizza, E.; Cerione, R.A.; Antonyak, M.A. Exosomes as Sentinels against Bacterial Pathogens. Dev. Cell 2020, 53, 138–139. [Google Scholar] [CrossRef] [PubMed]
- Los, F.C.O.; Randis, T.M.; Aroian, R.V.; Ratner, A.J. Role of Pore-Forming Toxins in Bacterial Infectious Diseases. Microbiol. Mol. Biol. Rev. 2013, 77, 173–207. [Google Scholar] [CrossRef] [Green Version]
- Orrell, K.E.; Mansfield, M.J.; Doxey, A.C.; Melnyk, R.A. The C. difficile toxin B membrane translocation machinery is an evolutionarily conserved protein delivery apparatus. Nat. Commun. 2020, 11, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giesemann, T.; Jank, T.; Gerhard, R.; Maier, E.; Just, I.; Benz, R.; Aktories, K. Cholesterol-dependent pore formation of Clostridium difficile toxin A. J. Biol. Chem. 2006, 281, 10808–10815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klepka, C.; Sandmann, M.; Tatge, H.; Mangan, M.; Arens, A.; Henkel, D.; Gerhard, R. Impairment of lysosomal function by Clostridioides difficile TcdB. Mol. Microbiol. 2022, 117, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Giugliano, L.G.; Drasar, B.S. The influence of drugs on the response of a cell culture preparation to bacterial toxins. J. Med. Microbiol. 1984, 17, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Caspar, M.; Florin, I.; Thelestam, M. Calcium and calmodulin in cellular intoxication with Clostridium difficile toxin B. J. Cell. Physiol. 1987, 132, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Fiorentini, C.; Donelli, G.; Nicotera, P.; Thelestam, M. Clostridium difficile toxin A elicits Ca2+-independent cytotoxic effects in cultured normal rat intestinal crypt cells. Infect. Immun. 1993, 61, 3988–3993. [Google Scholar] [CrossRef]
- Fiorentini, C.; Thelestam, M. Clostridium difficile toxin A and its effects on cells. Toxicon 1991, 29, 543–567. [Google Scholar] [CrossRef] [PubMed]
- Pothoulakis, C.; Sullivan, R.; Melnick, D.A.; Triadafilopoulos, G.; Gadenne, A.S.; Meshulam, T.; LaMont, J.T. Clostridium difficile toxin A stimulates intracellular calcium release and chemotactic response in human granulocytes. J. Clin. Investig. 1988, 81, 1741–1745. [Google Scholar] [CrossRef] [Green Version]
- Triadafilopoulos, G.; Pothoulakis, C.; O’Brien, M.J.; LaMont, J.T. Differential effects of Clostridium difficile toxins A and B on rabbit ileum. Gastroenterology 1987, 93, 273–279. [Google Scholar] [CrossRef]
- Ciesielski-Treska, J.; Ulrich, G.; Rihn, B.; Aunis, D. Mechanism of action of Clostridium difficile toxin B: Role of external medium and cytoskeletal organization in intoxicated cells. Eur. J. Cell Biol. 1989, 48, 191–202. [Google Scholar]
- Lima, A.A.M.; Lyerly, D.M.; Wilkins, T.D.; Innes, D.J.; Guerrant, R.L. Effects of Clostridium difficile toxins A and B in rabbit small and large intestine in vivo and on cultured cells in vitro. Infect. Immun. 1988, 56, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.S.; Musch, M.W.; Sugi, K.; Walsh-Reitz, M.M.; Ropeleski, M.J.; Hendrickson, B.A.; Pothoulakis, C.; Lamont, J.T.; Chang, E.B. Protective role of HSP72 against Clostridium difficile toxin A-induced intestinal epithelial cell dysfunction. Am. J. Physiol. Cell Physiol. 2003, 284, C1073–C1082. [Google Scholar] [CrossRef]
- Gilbert, R.J.; Pothoulakis, C.; Lamont, J.T.; Yakubovich, M. Clostridium difficile toxin B activates calcium influx required for actin disassembly during cytotoxicity. Am. J. Physiol. Gastrointest. Liver Physiol. 1995, 268, G487–G495. [Google Scholar] [CrossRef]
- Jefferson, K.K.; Smith, M.F.; Bobak, D.A. Roles of Intracellular Calcium and NF-κB in the Clostridium difficile Toxin A-Induced Up-Regulation and Secretion of IL-8 from Human Monocytes. J. Immunol. 1999, 163, 5183–5191. [Google Scholar] [CrossRef]
- Warny, M.; Keates, A.C.; Keates, S.; Castagliuolo, I.; Zacks, J.K.; Aboudola, S.; Qamar, A.; Pothoulakis, C.; LaMont, J.T.; Kelly, C.P. p38 MAP kinase activation by Clostridium difficile toxin A mediates monocyte necrosis, IL-8 production, and enteritis. J. Clin. Investig. 2000, 105, 1147–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.M.; Lee, J.Y.; Yoon, Y.M.; Oh, Y.K.; Youn, J.; Kim, Y.J. NF-κB activation pathway is essential for the chemokine expression in intestinal epithelial cells stimulated with Clostridium difficile toxin A. Scand. J. Immunol. 2006, 63, 453–460. [Google Scholar] [CrossRef]
- Demarest, S.J.; Salbato, J.; Elia, M.; Zhong, J.; Morrow, T.; Holland, T.; Kline, K.; Woodnutt, G.; Kimmel, B.E.; Hansen, G. Structural characterization of the cell wall binding domains of Clostridium difficile toxins A and B; evidence that Ca2+ plays a role in toxin A cell surface association. J. Mol. Biol. 2005, 346, 1197–1206. [Google Scholar] [CrossRef]
- Popoff, M.R. Clostridial pore-forming toxins: Powerful virulence factors. Anaerobe 2014, 30, 220–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goy, S.D.; Olling, A.; Neumann, D.; Pich, A.; Gerhard, R. Human neutrophils are activated by a peptide fragment of Clostridium difficile toxin B presumably via formyl peptide receptor. Cell. Microbiol. 2015, 17, 893–909. [Google Scholar] [CrossRef]
- Jiang, Y.; Xu, L.; Yu, L.; Xu, X.; Feng, C.; Li, J. NOX4 inhibition protects enteric glial cells against Clostridium difficile toxin B toxicity via attenuating oxidative and Endoplasmic reticulum stresses. Free Radic. Res. 2019, 53, 932–940. [Google Scholar] [CrossRef]
- Sarri, E.; Böckmann, I.; Kempter, U.; Valeva, A.; Von Eichel-Streiber, C.; Weichel, O.; Klein, J. Regulation of phospholipase D activity in synaptosomes permeabilized with Staphylococcus aureus α-toxin. FEBS Lett. 1998, 440, 287–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djouder, N.; Prepens, U.; Aktories, K.; Cavalié, A. Inhibition of calcium release-activated calcium current by Rac/Cdc42- inactivating clostridial cytotoxins in RBL cells. J. Biol. Chem. 2000, 275, 18732–18738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebres, R.A.; Moon, C.; DeCamp, D.; Lin, K.-M.; Fraser, I.D.; Milne, S.B.; Roach, T.I.A.; Brown, H.A.; Seaman, W.E. Clostridium difficile toxin B differentially affects GPCR-stimulated Ca2+ responses in macrophages: Independent roles for Rho and PLA2. J. Leukoc. Biol. 2010, 87, 1041–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loureiro, A.V.; Moura-Neto, L.I.; Martins, C.S.; Silva, P.I.M.; Lopes, M.B.S.; Leitão, R.F.C.; Coelho-Aguiar, J.M.; Moura-Neto, V.; Warren, C.A.; Costa, D.V.S.; et al. Role of Pannexin-1-P2X7R signaling on cell death and pro-inflammatory mediator expression induced by Clostridioides difficile toxins in enteric glia. Front. Immunol. 2022, 13, 1956340. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, H. Clostridium difficile Toxin A Upregulates Bak Expression through PGE2 Pathway in Human Colonocytes. J. Microbiol. Biotechnol. 2019, 29, 1675–1681. [Google Scholar] [CrossRef]
- Loureiro, A.V.; Barbosa, M.L.L.; Morais, M.L.G.S.; Souza, I.P.; Terceiro, L.S.; Martins, C.S.; Sousa, A.P.R.; Leitão, R.F.C.; Shin, J.H.; Warren, C.A.; et al. Host and Clostridioides difficile-Response Modulated by Micronutrients and Glutamine: An Overview. Front. Nutr. 2022, 9, 849301. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Zhang, Z.; McNeil, B.D.; Luo, F.; Wu, X.S.; Sheng, J.; Shin, W.; Wu, L.G. Voltage-Dependent Calcium Channels at the Plasma Membrane, but Not Vesicular Channels, Couple Exocytosis to Endocytosis. Cell Rep. 2012, 1, 632–638. [Google Scholar] [CrossRef] [Green Version]
- El Bekay, R.; Alba, G.; Reyes, M.E.; Chacón, P.; Vega, A.; Martín-Nieto, J.; Jiménez, J.; Ramos, E.; Oliván, J.; Pintado, E.; et al. Rac2 GTPase activation by angiotensin II is modulated by Ca2+ calcineurin and mitogen-activated protein kinases in human neutrophils. J. Mol. Endocrinol. 2007, 39, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Oiki, S.; Okada, Y. C1q induces chemotaxis and K+ conductance activation coupled to increased cytosolic Ca2+ in mouse fibroblasts. J. Immunol. 1988, 141, 3177–3185. [Google Scholar] [CrossRef]
- Smani, T.; Domínguez-Rodriguez, A.; Callejo-García, P.; Rosado, J.A.; Avila-Medina, J. Phospholipase A2 as a molecular determinant of store-operated calcium entry. Adv. Exp. Med. Biol. 2016, 898, 111–131. [Google Scholar]
- Sharma, S.V. Melittin-induced hyperactivation of phospholipase A2 activity and calcium influx in ras-transformed cells. Oncogene 1993, 8, 939–947. [Google Scholar]
- Reddy, S.; Bose, R.; Rao, G.H.; Murthy, M. Phospholipase A2 activation in human neutrophils requires influx of extracellular Ca2+ and leukotriene B4. Am. J. Physiol. Cell Physiol. 1995, 268, C138–C146. [Google Scholar] [CrossRef]
- Rzigalinski, B.A.; Blackmore, P.F.; Rosenthal, M.D. Arachidonate mobilization is coupled to depletion of intracellular calcium stores and influx of extracellular calcium in differentiated U937 cells. Biochim. Biophys. Acta Lipids Lipid Metab. 1996, 1299, 342–352. [Google Scholar] [CrossRef]
- Millanvoye-Van Brussel, E.; David-Dufilho, M.; Pham, T.D.; Iouzalen, L.; Devynck, M.A. Regulation of arachidonic acid release by calcium influx in human endothelial cells. J. Vasc. Res. 1999, 36, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.A.M.; Nascimento, N.R.F.; Fang, G.D.; Yotseff, P.; Toyama, M.H.; Guerrant, R.L.; Fonteles, M.C. Role of phospholipase A2 and tyrosine kinase in Clostridium difficile toxin A-induced disruption of epithelial integrity, histologic inflammatory damage and intestinal secretion. J. Appl. Toxicol. 2008, 28, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Shoshan, M.C.; Florin, I.; Thelestam, M. Activation of cellular phospholipase A2 by Clostridium difficile toxin B. J. Cell. Biochem. 1993, 52, 116–124. [Google Scholar] [CrossRef]
- Florin, I.; Thelestam, M. Lysosomal involvement in cellular intoxication with Clostridium difficile toxin B. Microb. Pathog. 1986, 1, 373–385. [Google Scholar] [CrossRef]
- Wong, J.T.; Tran, K.; Pierce, G.N.; Chan, A.C.; Karmin, O.; Choy, P.C. Lysophosphtidylcholine stimulates the release of arachidonic acid in human endothelial cells. J. Biol. Chem. 1998, 273, 6830–6836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, X.; Thibodeau, A.; Forte, J.G. Ezrin-calpain I interactions in gastric parietal cells. Am. J. Physiol. Cell Physiol. 1993, 265, C36–C46. [Google Scholar] [CrossRef]
- Wang, F.; Xia, P.; Wu, F.; Wang, D.; Wang, W.; Ward, T.; Liu, Y.; Aikhionbare, F.; Guo, Z.; Powell, M.; et al. Helicobacter pylori VacA disrupts apical membrane-cytoskeletal interactions in gastric parietal cells. J. Biol. Chem. 2008, 283, 26714–26725. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.E.; Vervliet, T.; Bultynck, G.; Parys, J.B.; Hallett, M.B. EPIC3, a novel Ca2+ indicator located at the cell cortex and in microridges, detects high Ca2+ subdomains during Ca2+ influx and phagocytosis. Cell Calcium 2020, 92, 102291. [Google Scholar] [CrossRef]
- Marlink, K.L.; Bacon, K.D.; Sheppard, B.C.; Ashktorab, H.; Smoot, D.T.; Cover, T.L.; Deveney, C.W.; Rutten, M.J. Effects of Helicobacter pylori on intracellular Ca2+ signaling in normal human gastric mucous epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G163–G176. [Google Scholar] [CrossRef]
- Gómez, J.; Martínez-A, C.; González, A.; García, A.; Rebollo, A. The Bcl-2 gene is differentially regulated by IL-2 and IL-4: Role of the transcription factor NF-AT. Oncogene 1998, 17, 1235–1243. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.L.; Pothoulakis, C.; Thomas Lamont, J. Protein kinase C signaling regulates ZO-1 translocation and increased paracellular flux of T84 colonocytes exposed to Clostridium difficile toxin A. J. Biol. Chem. 2002, 277, 4247–4254. [Google Scholar] [CrossRef] [Green Version]
- Pothoulakis, C. Effects of Clostridium difficile toxins on epithelial cell barrier. Ann. N. Y. Acad. Sci. 2000, 915, 347–356. [Google Scholar] [CrossRef]
- Lee, J.; Song, C.H. Effect of reactive oxygen species on the endoplasmic reticulum and mitochondria during intracellular pathogen infection of mammalian cells. Antioxidants 2021, 10, 872. [Google Scholar] [CrossRef]
- Kennedy, C.L.; Smith, D.J.; Lyras, D.; Chakravorty, A.; Rood, J.I. Programmed cellular necrosis mediated by the pore-forming α-toxin from Clostridium septicum. PLoS Pathog. 2009, 5, e1000516. [Google Scholar] [CrossRef] [Green Version]
- Adamson, R.H.; Curry, F.E.; Adamson, G.; Liu, B.; Jiang, Y.; Aktories, K.; Barth, H.; Daigeler, A.; Golenhofen, N.; Ness, W.; et al. Rho and rho kinase modulation of barrier properties: Cultured endothelial cells and intact microvessels of rats and mice. J. Physiol. 2002, 539, 295–308. [Google Scholar] [CrossRef]
- Otake, K.; Sato, N.; Kitaguchi, A.; Irahara, T.; Murata, S.; Shiraga, K.; Ogawa, Y.; Fujiwara, T.K.; Koike, K.; Yokota, H. The Effect of Lactoferrin and Pepsin-Treated Lactoferrin on IEC-6 Cell Damage Induced by Clostridium difficile Toxin B. Shock 2018, 50, 119–125. [Google Scholar] [CrossRef]
- Zemljic, M.; Rupnik, M.; Scarpa, M.; Anderluh, G.; Palù, G.; Castagliuolo, I. Repetitive domain of Clostridium difficile toxin B exhibits cytotoxic effects on human intestinal epithelial cells and decreases epithelial barrier function. Anaerobe 2010, 16, 527–532. [Google Scholar] [CrossRef]
- Panther, E.; Dürk, T.; Ferrari, D.; Di Virgilio, F.; Grimm, M.; Sorichter, S.; Cicko, S.; Herouy, Y.; Norgauer, J.; Idzko, M.; et al. AMP affects intracellular Ca2+ signaling, migration, cytokine secretion and T cell priming capacity of dendritic cells. PLoS ONE 2012, 7, e37560. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.H.; Wu, T.C.; Tsai, B.Y.; Hung, Y.P.; Lin, H.J.; Tsai, Y.S.; Ko, W.C.; Tsai, P.J. Peroxisome proliferator-activated receptor-γ as the gatekeeper of tight junction in Clostridioides difficile infection. Front. Microbiol. 2022, 13, 986457. [Google Scholar] [CrossRef] [PubMed]
- Nusrat, A.; Von Eichel-Streiber, C.; Turner, J.R.; Verkade, P.; Madara, J.L.; Parkos, C.A. Clostridium difficile toxins disrupt epithelial barrier function by altering membrane microdomain localization of tight junction proteins. Infect. Immun. 2001, 69, 1329–1336. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Rhee, S.H.; Pothoulakis, C.; LaMont, J.T. Clostridium difficile toxin A binds colonocyte Src causing dephosphorylation of focal adhesion kinase and paxillin. Exp. Cell Res. 2009, 315, 3336–3344. [Google Scholar] [CrossRef] [Green Version]
- Yanda, M.K.; Guggino, W.B.; Cebotaru, L. A new role for heat shock factor 27 in the pathophysiology of Clostridium difficile toxin B. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G120–G129. [Google Scholar] [CrossRef]
- Knapp, O.; Stiles, B.; Popoff, M.R. The Aerolysin-Like Toxin Family of Cytolytic, Pore-Forming Toxins. Open Toxinol. J. 2010, 3, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Boura, E.; Ivanov, V.; Carlson, L.A.; Mizuuchi, K.; Hurley, J.H. Endosomal sorting complex required for transport (ESCRT) complexes induce phase-separated microdomains in supported lipid bilayers. J. Biol. Chem. 2012, 287, 28144–28151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Wang, M.; Zhang, Y.Y.; Zhao, S.Z.; Gu, S. The endosomal sorting complex required for transport repairs the membrane to delay cell death. Front. Oncol. 2022, 12, 1007446. [Google Scholar] [CrossRef]
- Crespin, M.; Vidal, C.; Picard, F.; Lacombe, C.; Fontenay, M. Activation of PAK1/2 during the shedding of platelet microvesicles. Blood Coagul Fibrinolysis 2009, 20, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Huber, P. Targeting of the apical junctional complex by bacterial pathogens. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183237. [Google Scholar] [CrossRef]
- Keller, M.D.; Ching, K.L.; Liang, F.X.; Dhabaria, A.; Tam, K.; Ueberheide, B.M.; Unutmaz, D.; Torres, V.J.; Cadwell, K. Decoy exosomes provide protection against bacterial toxins. Nature 2020, 579, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Markovska, R.; Dimitrov, G.; Gergova, R.; Boyanova, L. Clostridioides difficile, a New “Superbug”. Microorganisms 2023, 11, 845. [Google Scholar] [CrossRef] [PubMed]
- Kajihara, T.; Yahara, K.; Kitamura, N.; Hirabayashi, A.; Hosaka, Y.; Sugai, M. Distribution, Trends, and Antimicrobial Susceptibility of Bacteroides, Clostridium, Fusobacterium, and Prevotella Species Causing Bacteremia in Japan During 2011–2020: A Retrospective Observational Study Based on National Surveillance Data. Med. Mycol. 2022, 60, ofad334. [Google Scholar] [CrossRef]
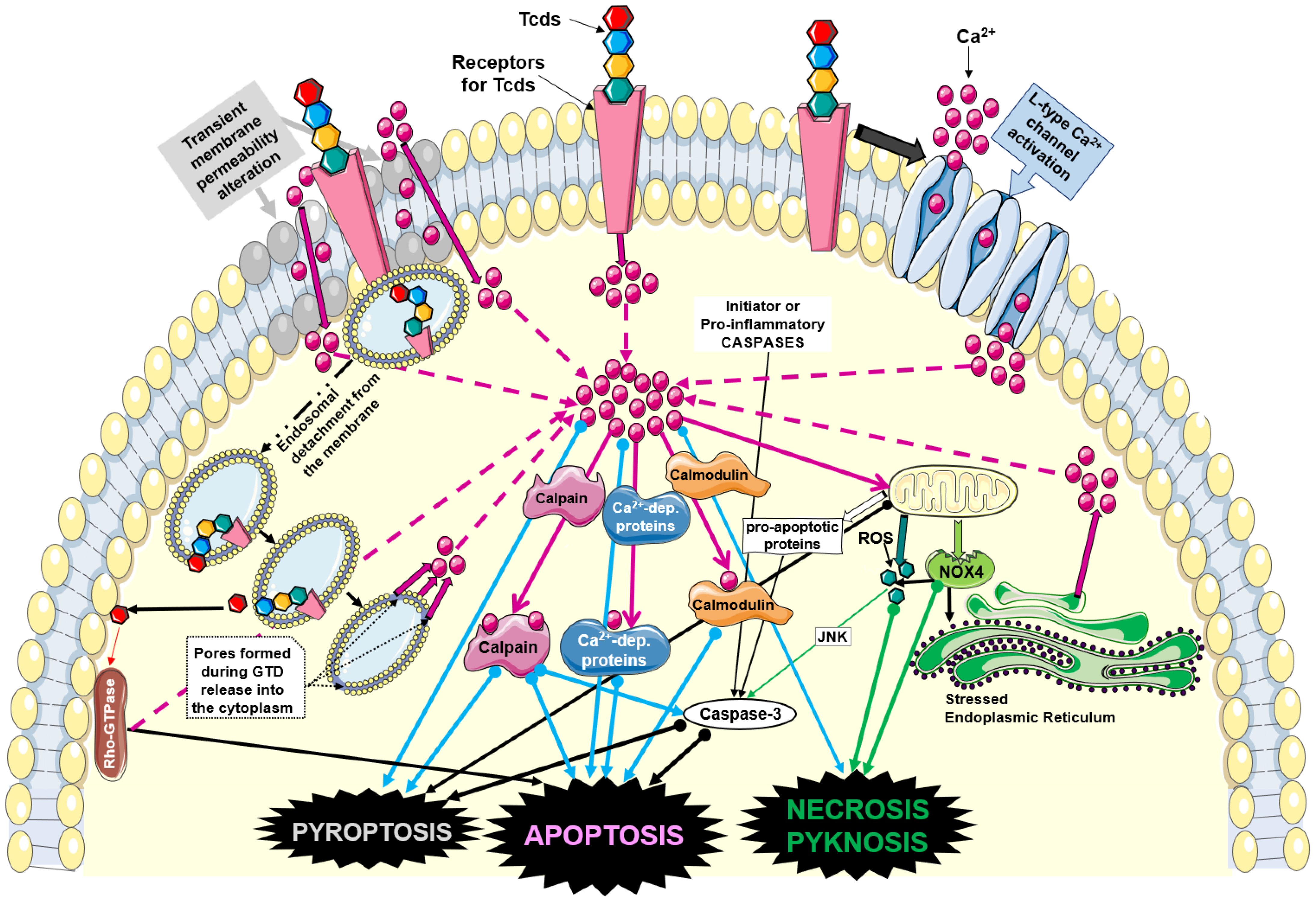

{kind=link}
| Mechanism | PFTs | Tcds |
|---|---|---|
| Formation of Ca2+ channels on the plasma membrane | PFTs monomer polymerizes within the plasma membrane, forming a channel that favors the influx of Ca2+ based on the extracellular gradient of Ca2+ which is greater than the intracellular one Refs. [48,113,114,115,116,138] | Tcds do not form pores in the plasma membrane but can activate the influx of Ca2+ by stimulating the receptors as FZDs. In addition, transient changes in the continuity of the plasma membrane during the endocytosis process of Tcds can favor Ca2+ influx Refs. [8,9,10,11,51,93,121,122,123,133,147] |
| Activation of selective Ca2+ channels in the plasma membrane | Demonstrated with inhibitors Refs. [48,113,114,115,116,138] | Demonstrated with inhibitors Refs. [82,142,148,149] |
| Activation of Ca2+ channels in the endoplasmic reticulum | PFTs induce Ca2+ release into the cytoplasm by involving IP3 and IP3R, membrane G-proteins, phospholipase A2 Refs. [48,113,114,115,116,138,150,151,152,153,154] | There are no data on the interaction between Tcds and G-proteins. However, TcdA is capable of activating phospholipase A2 and thus to generate IP3 Refs. [155,156] |
| Activation of Ca2+ channels in lysosomes | PFTs induce Ca2+ release from lysosomes by the formation of two-pore channels and the involvement of CD38 and NAADP Refs. [48,113,114,115,116,138] | Tcds alter lysosomal function as demonstrated by cathepsin release implicating alterations in lysosomal Ca2+ homeostasis Refs. [47,86,123,157] |
| Activation of Ca2+-dependent cytoplasmic proteins | PFT-induced Ca2+ influx activates calpains, calmodulin, calcineurin, PKC and phospholipase A2 Refs. [48,113,114,115,116,138,158,159,160,161,162] | Alteration in Ca2+ homeostasis activates calpains, calmodulin, PKC and phospholipase A2 Refs. [47,125,143,155,156,163,164,165] |
| Cell Death | PFTs induce: Necrosis Necroptosis Apoptosis Pyroptosis Refs. [48,101,113,114,115,116,138,166,167] | Tcds induce: Necrosis Apoptosis Pyroptosis Refs. [8,9,10,11,12,46,47,61,62,63,73,82,84,86,91,94,102,103,104,105,107,108,110,111,126,127,128,129,130,131,132,133,140,142,144,145] |
| Destruction of the intercellular junctions | Intercellular junctions are targets of PTFs Refs. [48,113,114,115,116,138] | Intercellular junctions are targets of Tcds Refs. [8,9,10,11,60,164,168,169,170,171,172,173,174,175] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fettucciari, K.; Dini, F.; Marconi, P.; Bassotti, G. Role of the Alteration in Calcium Homeostasis in Cell Death Induced by Clostridioides difficile Toxin A and Toxin B. Biology 2023, 12, 1117. https://doi.org/10.3390/biology12081117
Fettucciari K, Dini F, Marconi P, Bassotti G. Role of the Alteration in Calcium Homeostasis in Cell Death Induced by Clostridioides difficile Toxin A and Toxin B. Biology. 2023; 12(8):1117. https://doi.org/10.3390/biology12081117
Chicago/Turabian StyleFettucciari, Katia, Fabrizio Dini, Pierfrancesco Marconi, and Gabrio Bassotti. 2023. "Role of the Alteration in Calcium Homeostasis in Cell Death Induced by Clostridioides difficile Toxin A and Toxin B" Biology 12, no. 8: 1117. https://doi.org/10.3390/biology12081117