

Evaluation of Tacrolimus’ Adverse Effects on Zebrafish in Larval and Adult Stages by Using Multiple Physiological and Behavioral Endpoints

 ,
,  ,
,  , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models | Dose | Route of Administration | Adverse Effects | References |
|---|---|---|---|---|
| Human | Not described | Intravenous |
| [14] |
| 0.15 mg/kg | Intravenous, followed by oral dosing |
| [15] | |
| 0.075 mg/kg | Intravenous |
| [16] | |
| Baboon and dog | 0.1–2 mg/kg/d |
|
| [17] |
| Cynomolgus monkey | 1–10 mg/kg/d | Oral or intramuscular |
| [18] |
| Dog | 0.08–1 mg/kg/d | Oral or intramuscular |
| [19] |
| Rabbit | 1 mg/kg/d | Intramuscular injections |
| [20] |
| 10–1000 µg | Injection into the mídvitreous cavity of the eyes |
| [21] | |
| Rat | 0.5–4 mg/kg/d | Oral |
| [22] |
| 1 mg/kg/d | Intramuscular injections |
| [23] | |
| Betta splendens | 0.05 and 0.1 µg/mL | Waterborne exposure |
| [7] |
| Danio rerio | 0.1 µg/mL | Waterborne exposure |
| [13] |
2. Material and Methods
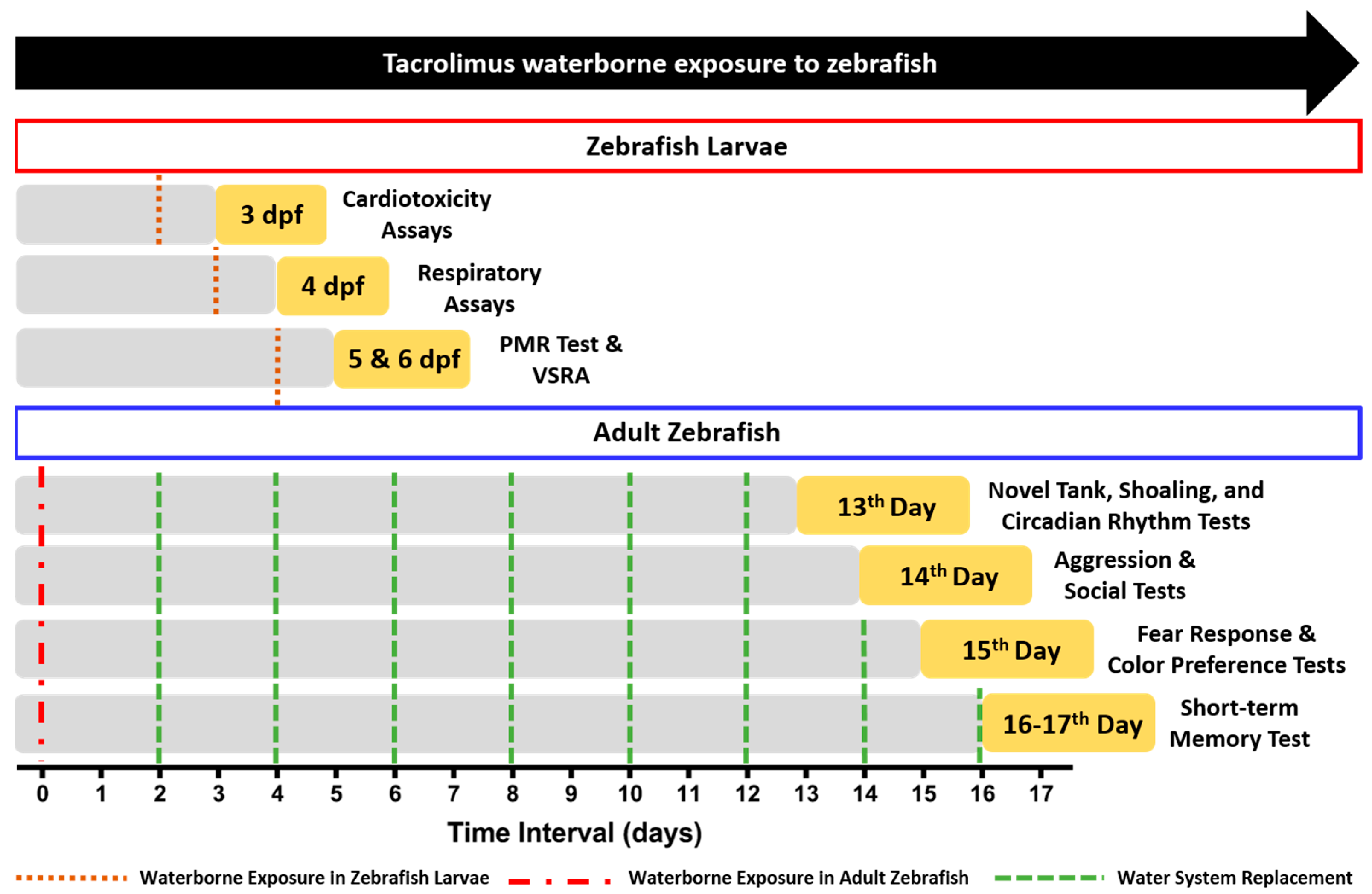
2.1. Zebrafish Husbandry
2.2. Chemical Delivery
2.3. Cardiac Performance Assays in Zebrafish Larvae
2.4. Respiratory Rate Assay in Zebrafish Larvae
2.5. Light-Dark Locomotion and Vibrational Startle Response Assays in Zebrafish Larvae
2.6. Multiple Behaviors Test in Adult Zebrafish
2.6.1. Novel Tank Test
2.6.2. Shoaling Test
2.6.3. Aggressiveness Test
2.6.4. Conspecific Social Interaction Test
2.6.5. Fear Response Test
2.7. Circadian Rhythm Locomotion Activity Test in Adult Zebrafish
2.8. Color Preference Assay in Adult Zebrafish
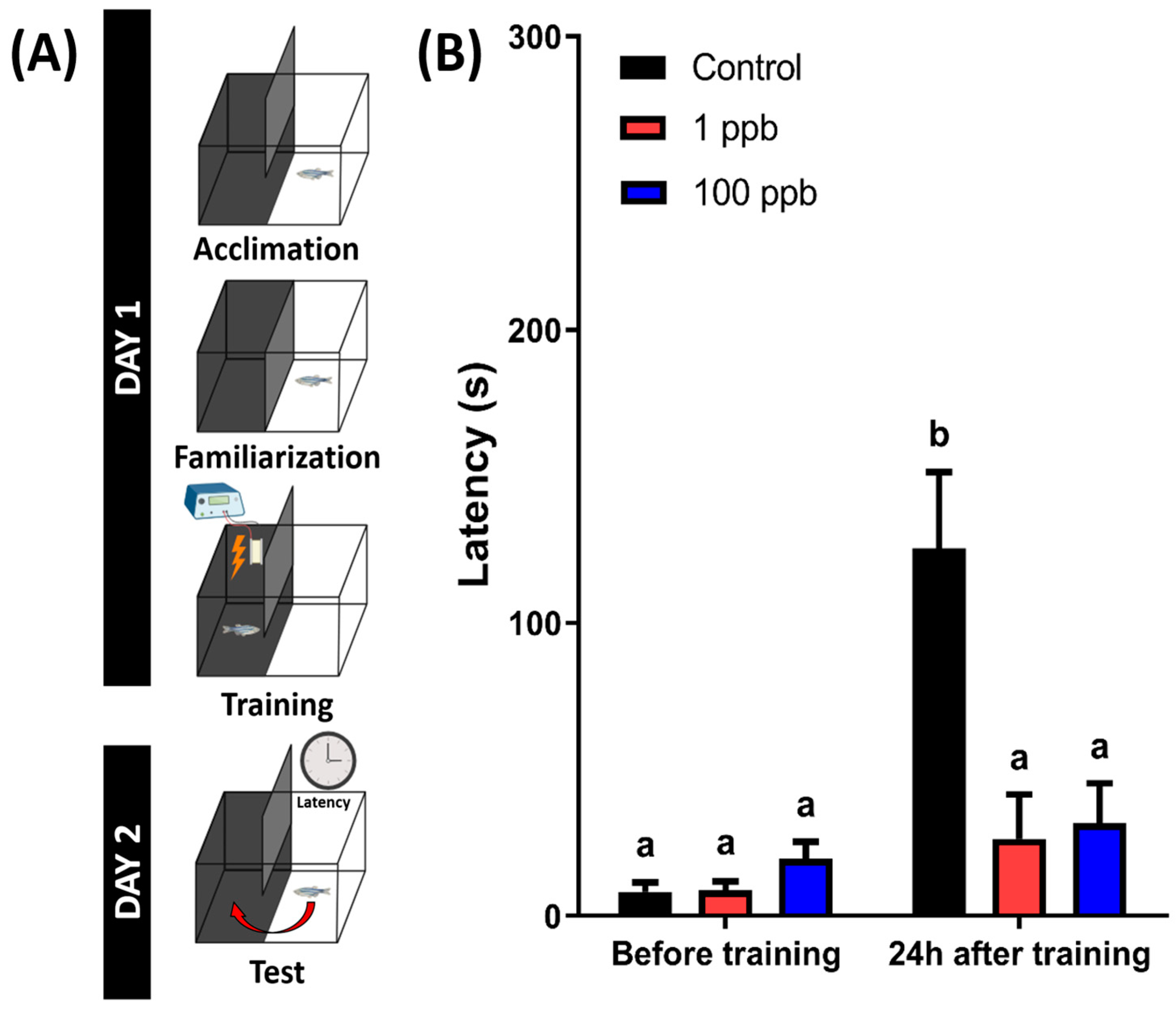
2.9. Passive Avoidance Test (Short-Term Memory Test) in Adult Zebrafish
2.10. Statistical Analyses
2.11. Molecular Docking 1FKJ Human FKBP 12 in Complex with Tacrolimus
3. Results
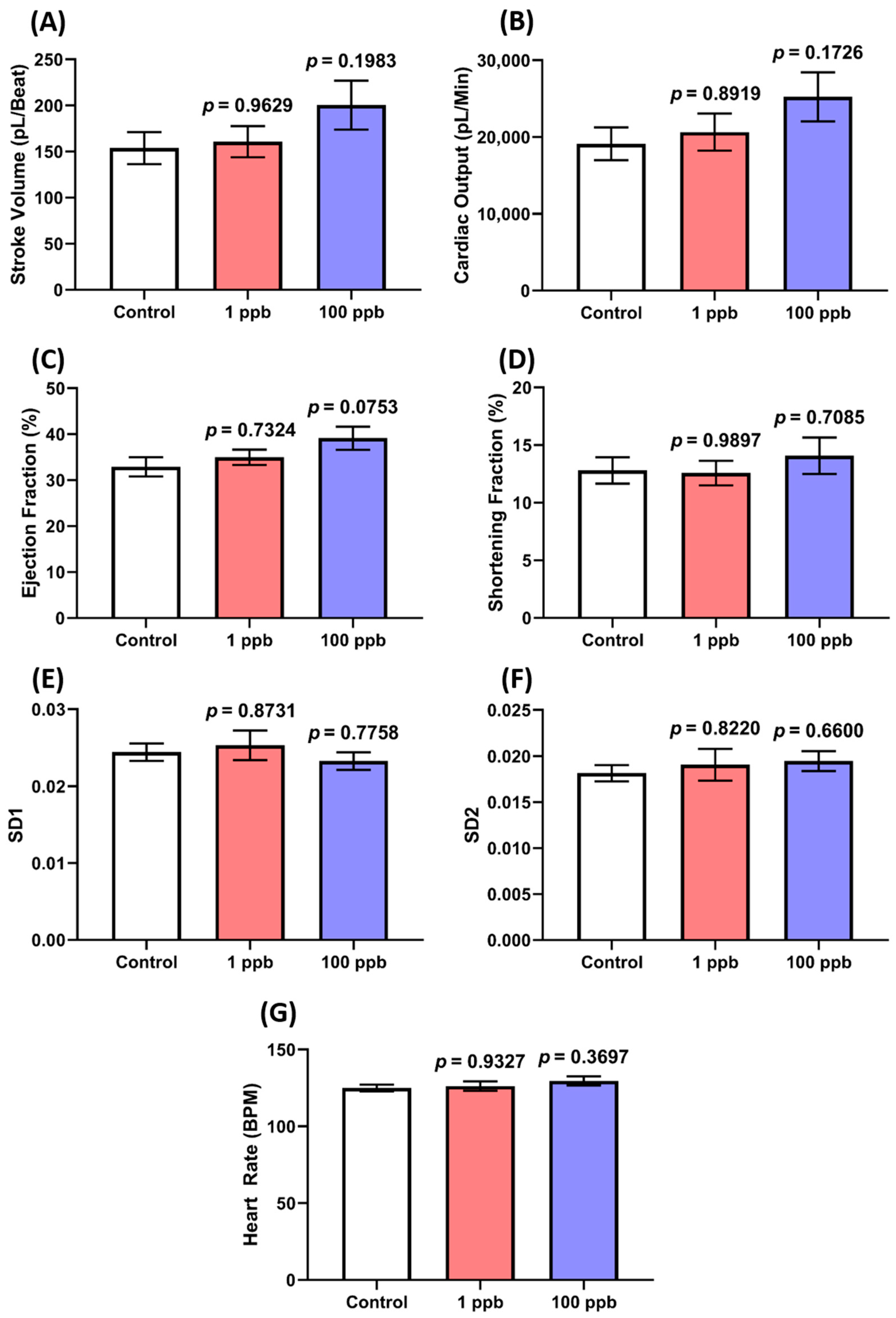
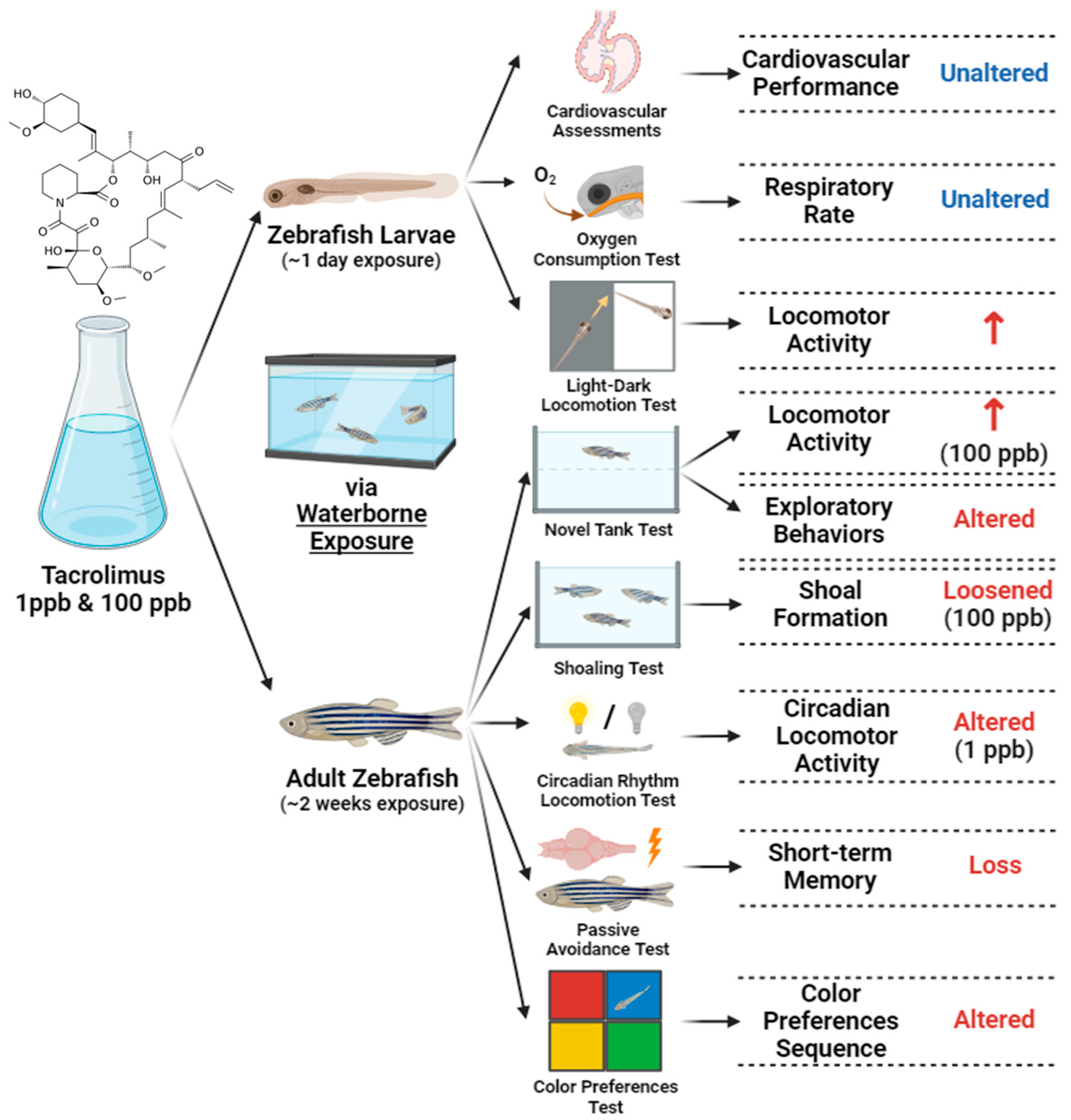
3.1. Acute Exposure to Tacrolimus Did Not Alter the Cardiac Rhythm and Physiology of Zebrafish Larvae
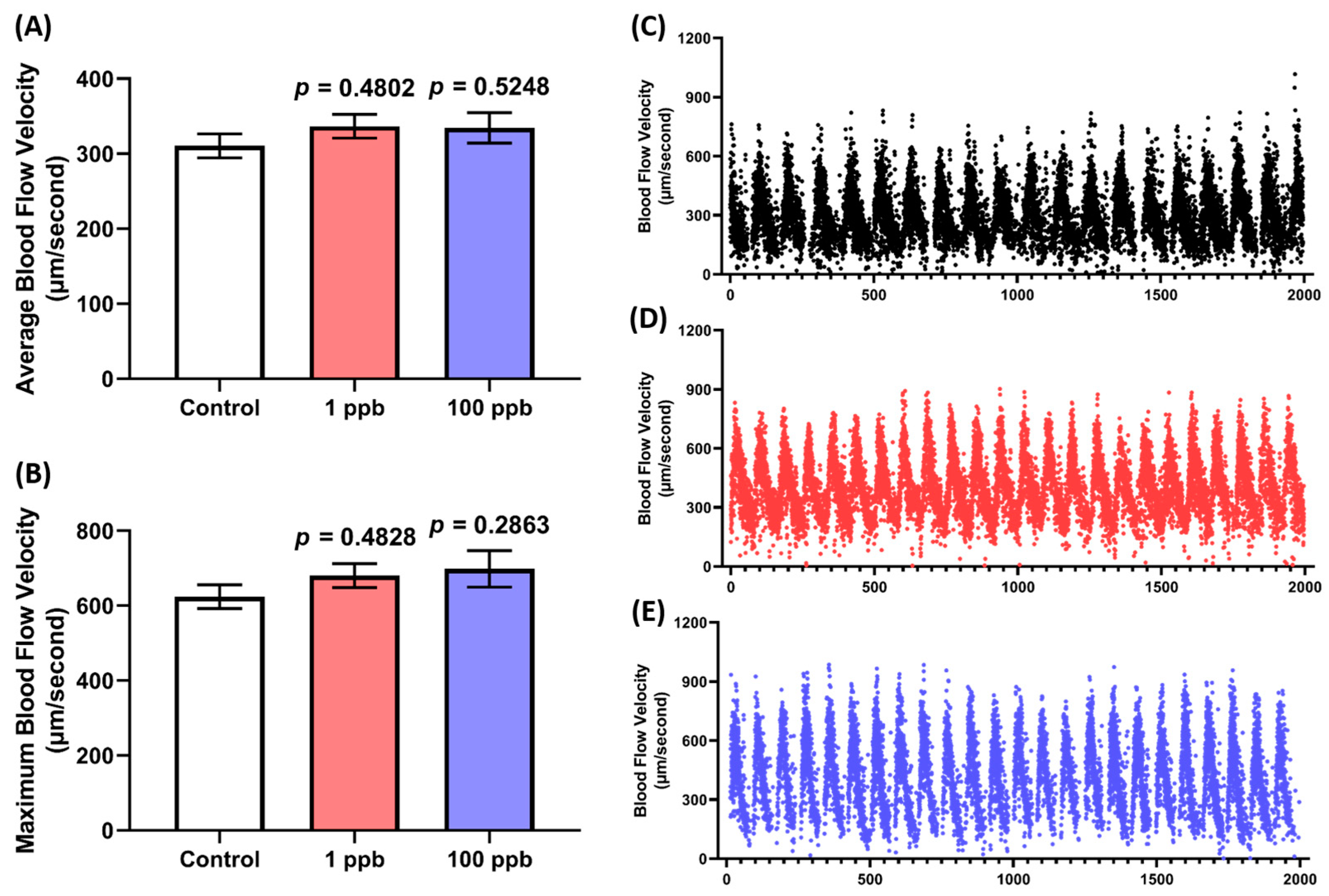
3.2. Acute Exposure to Tacrolimus Did Not Affect the Blood Flow of Zebrafish Larvae
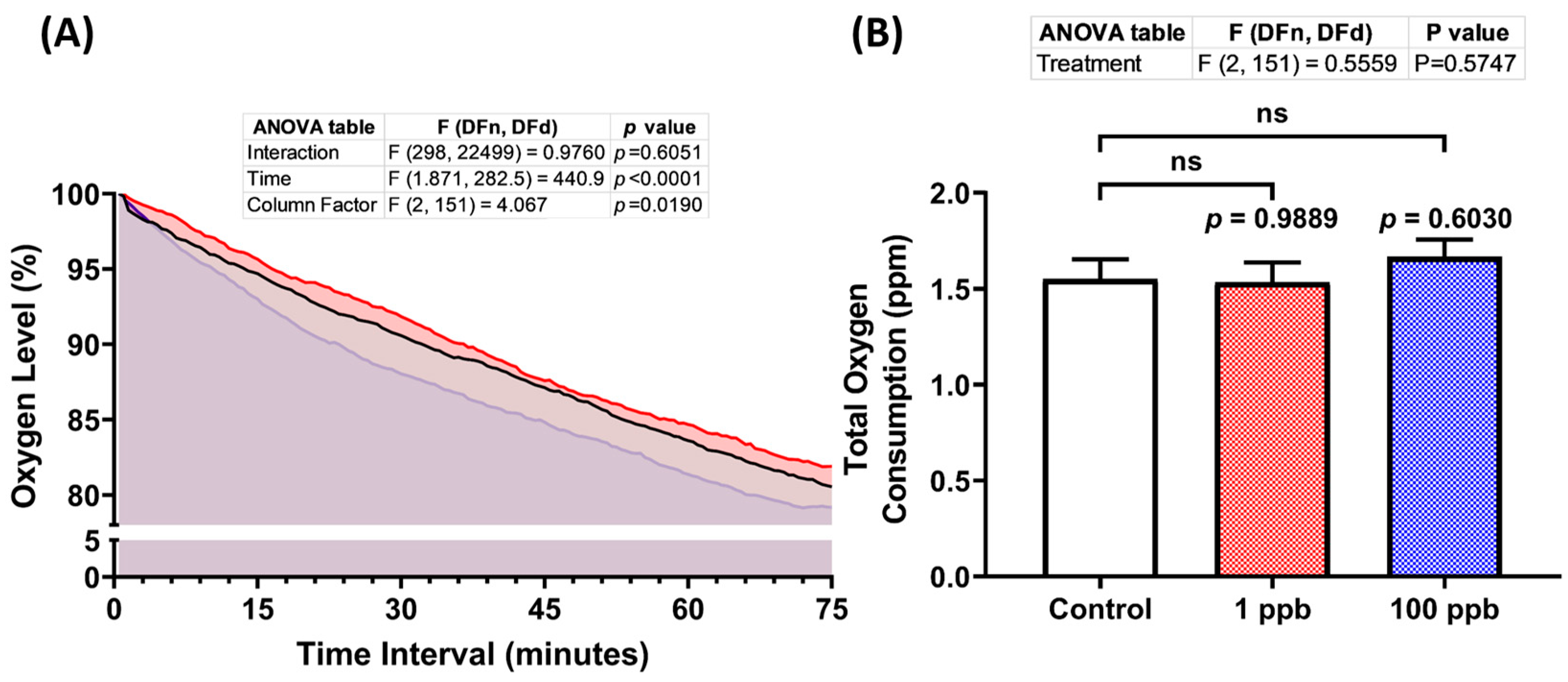
3.3. Acute Exposure to Tacrolimus Did Not Alter Zebrafish Larval Respiratory Rate
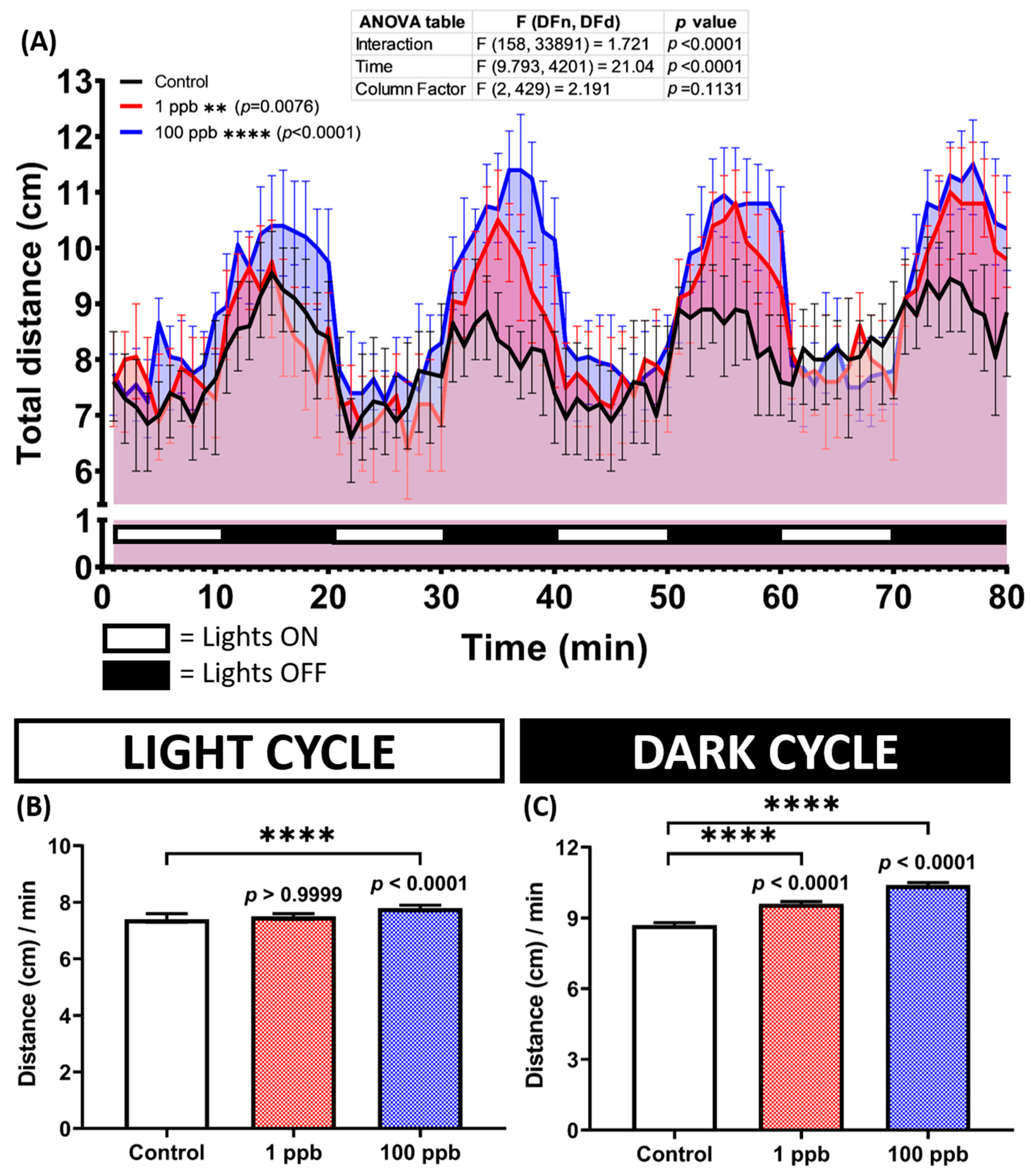
3.4. Acute Exposure to Tacrolimus Increased the Zebrafish Larval Locomotion during the Light–Dark Locomotion Test
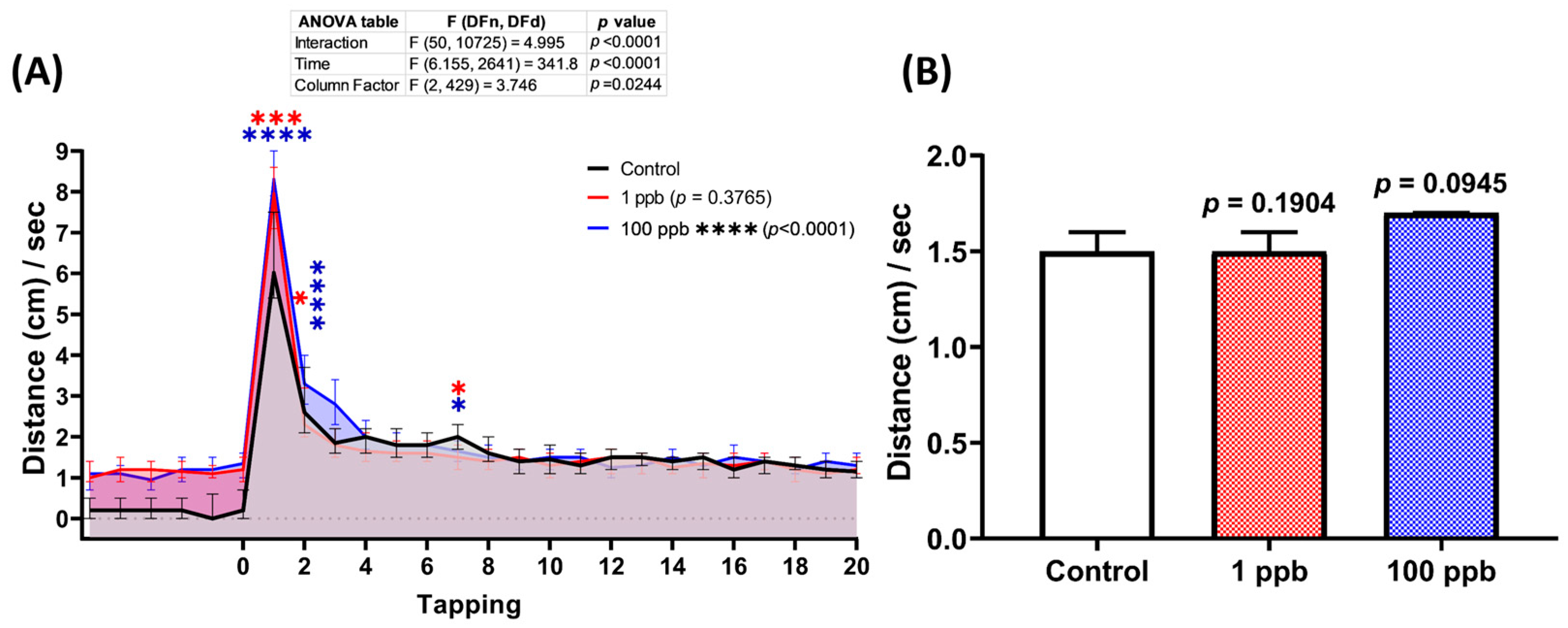
3.5. Acute Exposure to Tacrolimus Slightly Reduced the Habituation of Vibrational Startle Response (VSR) of Zebrafish Larvae
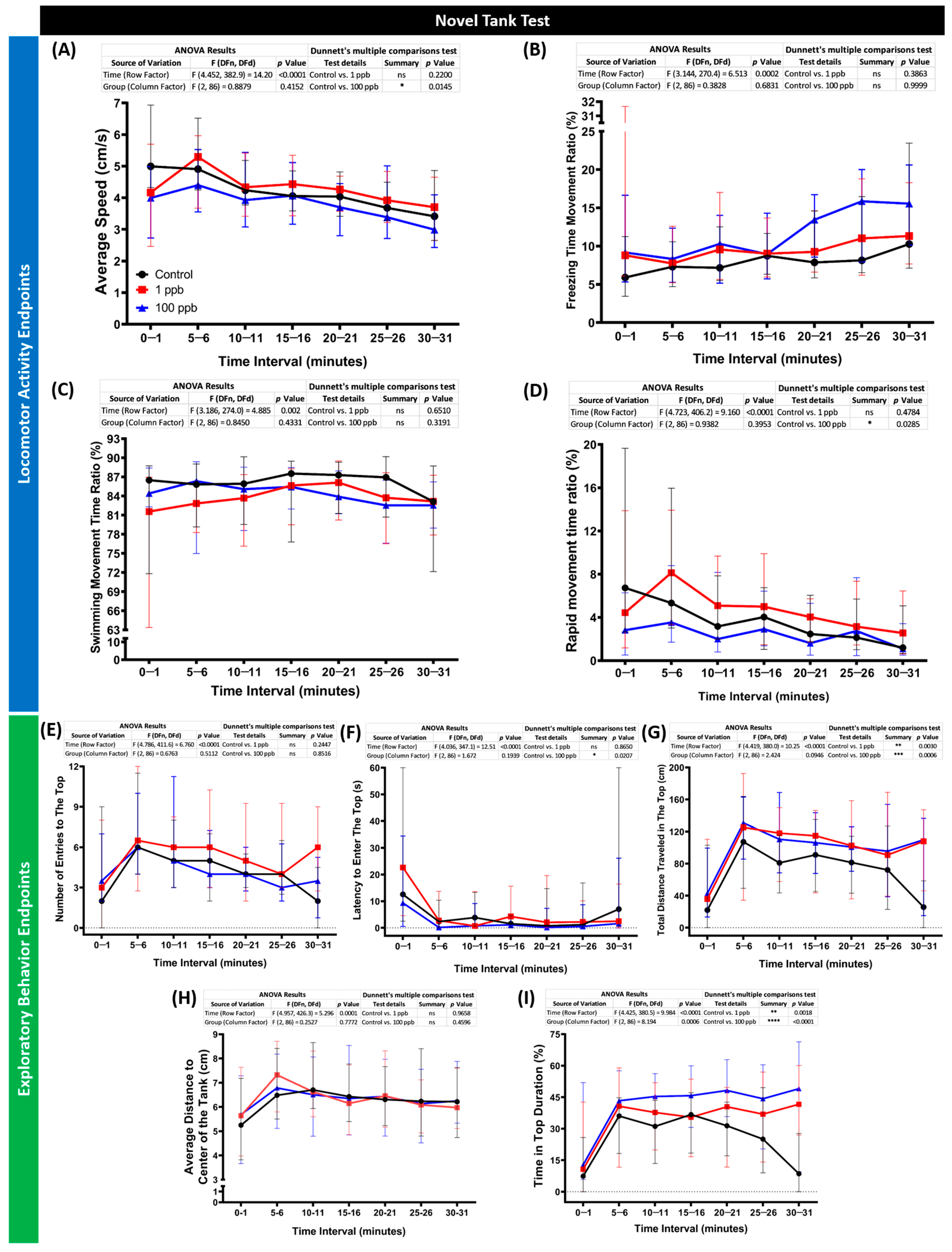
3.6. Chronic Exposure to Tacrolimus Reduced Locomotion and Altered Exploratory Behaviors of Zebrafish in the Novel Tank Test
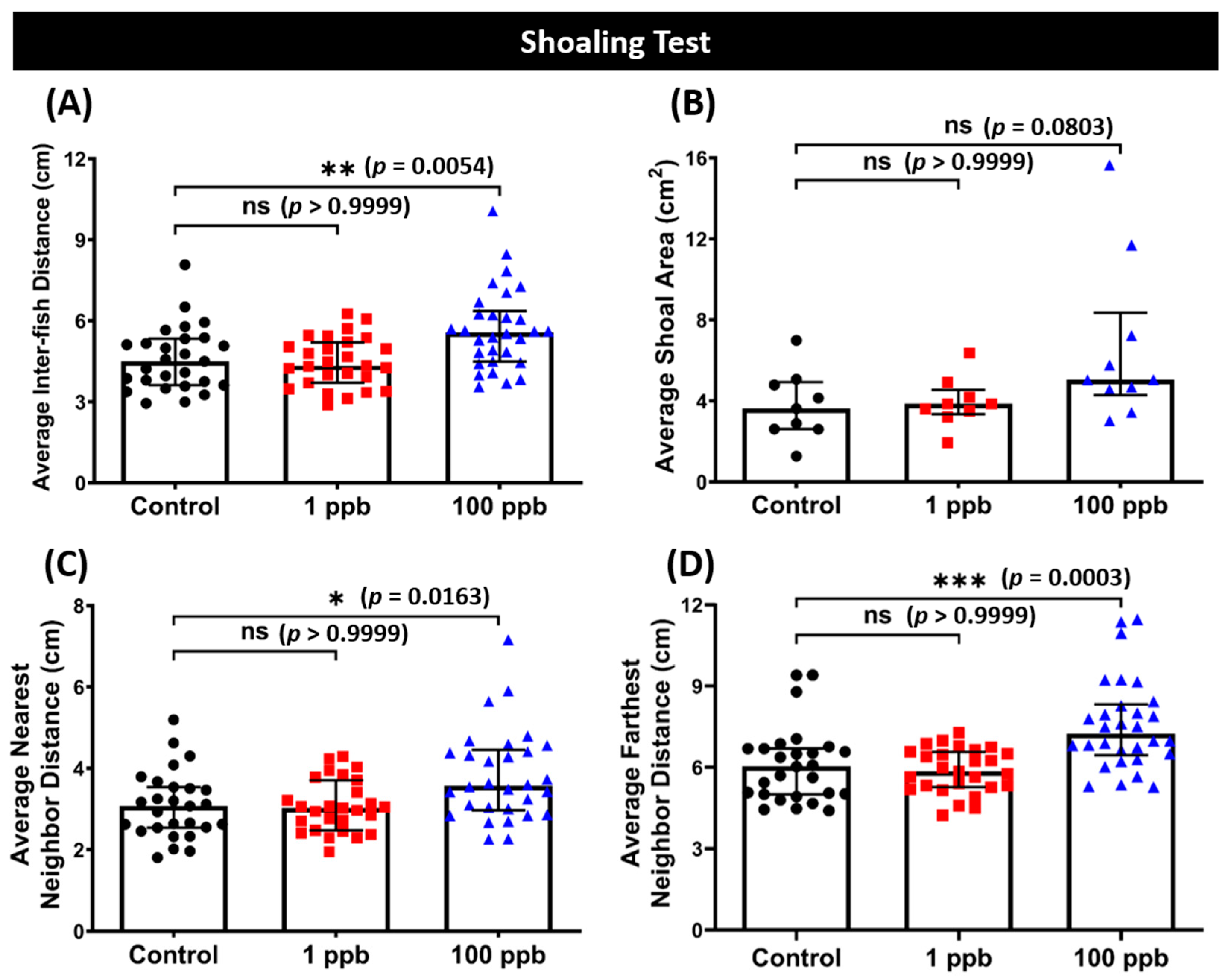
3.7. Chronic Exposure to Tacrolimus in a High Dose Resulted in Loosened Shoal Formation of Zebrafish
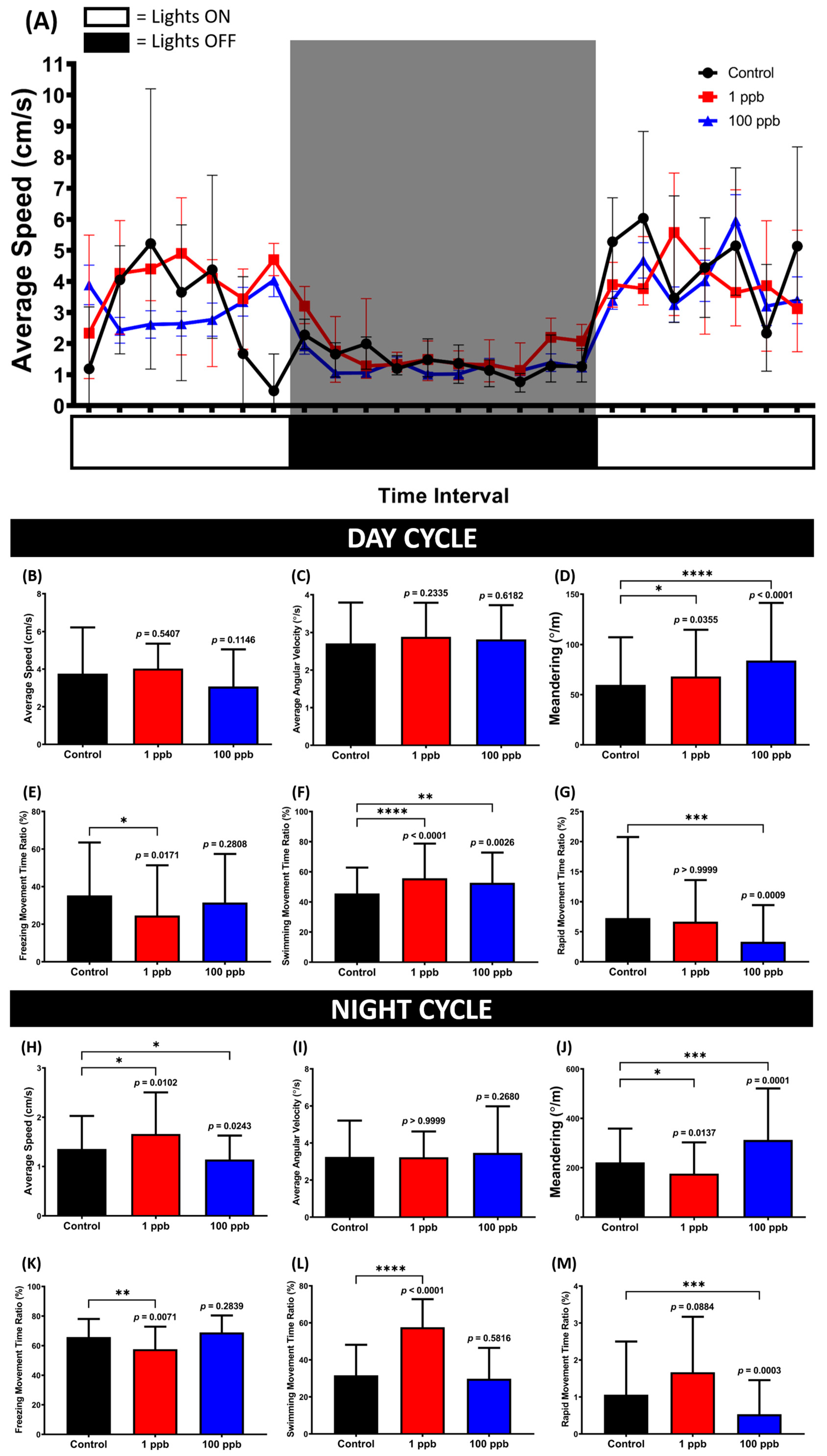
3.8. Chronic Exposure to Tacrolimus Caused Some Changes in the Circadian Rhythm Locomotor Activity of Adult Zebrafish
3.9. Chronic Exposure to Tacrolimus Induced Short-Term Memory Loss in Adult Zebrafish
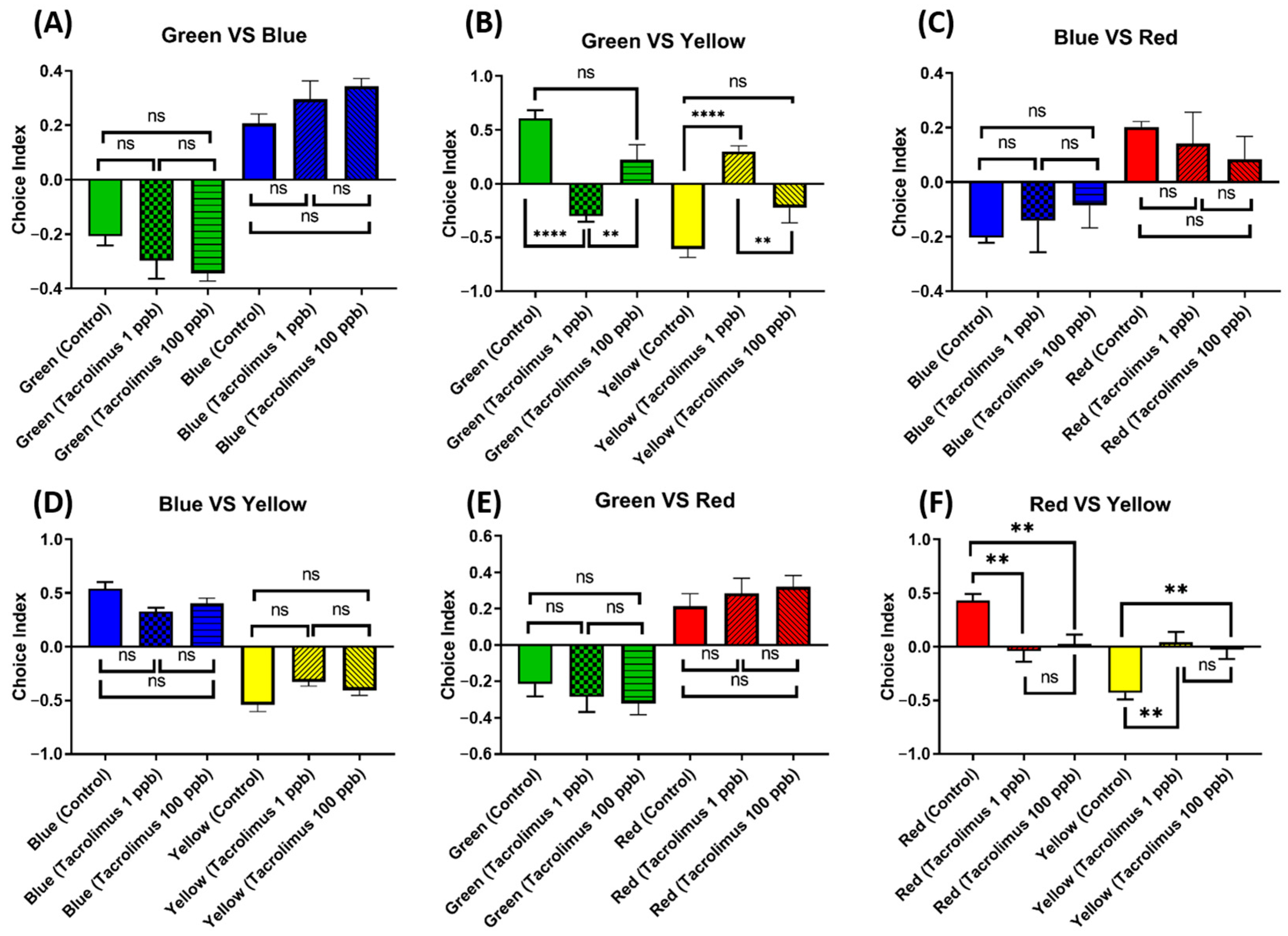
3.10. Chronic Exposure to Tacrolimus Altered the Color Preferences of Zebrafish
3.11. FKJ Human FKBP12 in Complex with Tacrolimus
4. Discussion
4.1. Tacrolimus in Affecting Locomotion in Zebrafish Larvae
4.2. High Dose of Tacrolimus in Affecting the Behavior Performance in a Novel Environment and Shoaling Formation of Adult Zebrafish
4.3. Low Dose of Tacrolimus in Affecting the Behavior Performance in a Novel Environment and Circadian Rhythm Locomotor Activity of Adult Zebrafish
4.4. Tacrolimus in Altering the Color Performance of Adult Zebrafish
4.5. Tacrolimus in Compromising the Cognitive Performance of Adult Zebrafish
4.6. The Binding Energies of Tacrolimus against Human FKB12
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jewett, A.; Tseng, H.-C. Pharmacology and Therapeutics for Dentistry, 7th ed.; Mariotti, A., Dowd, F.J., Johnson, B.S., Eds.; Elsevier Health Sciences: St. Louis, MI, USA, 2017. [Google Scholar]
- McCulloh, J.R. Infectious Diseases, 4th ed.; Cohen, J., Powderly, W.G., Opal, S.M., Eds.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Campagne, O.; Mager, D.E.; Brazeau, D.; Venuto, R.C.; Tornatore, K.M. The impact of tacrolimus exposure on extrarenal adverse effects in adult renal transplant recipients. Br. J. Clin. Pharmacol. 2019, 85, 516–529. [Google Scholar] [CrossRef] [PubMed]
- Kemper, M.J.; Spartà, G.; Laube, G.F.; Miozzari, M.; Neuhaus, T.J. Neuropsychologic side-effects of tacrolimus in pediatric renal transplantation. Clin. Transplant. 2003, 17, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Brooks, E.; Tett, S.E.; Isbel, N.M.; Staatz, C.E. Population pharmacokinetic modelling and Bayesian estimation of tacrolimus exposure: Is this clinically useful for dosage prediction yet? Clin. Pharmacokinet. 2016, 55, 1295–1335. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Oliva, A.; Hernández-Ávalos, I.; Martínez-Burnes, J.; Olmos-Hernández, A.; Verduzco-Mendoza, A.; Mota-Rojas, D. The Importance of Animal Models in Biomedical Research: Current Insights and Applications. Animals 2023, 13, 1223. [Google Scholar] [CrossRef] [PubMed]
- Javanshir Khoei, A.; Forsatkar, M.N.; Brown, C. Behavioral and immunotoxic effects of Prograf®(tacrolimus) in the male Siamese fighting fish. Ecotoxicology 2019, 28, 1032–1037. [Google Scholar] [CrossRef] [PubMed]
- Edmister, S.T.; Ibrahim, R.; Kakodkar, R.; Kreiling, J.A.; Creton, R. A zebrafish model for calcineurin-dependent brain function. Behav. Brain Res. 2022, 416, 113544. [Google Scholar] [CrossRef] [PubMed]
- Benchoula, K.; Khatib, A.; Jaffar, A.; Ahmed, Q.U.; Sulaiman, W.M.A.W.; Abd Wahab, R.; El-Seedi, H.R. The promise of zebrafish as a model of metabolic syndrome. Exp. Anim. 2019, 68, 407–416. [Google Scholar] [CrossRef]
- Mueller, T.; Wullimann, M. Chapter 2-Atlas of cellular markers in zebrafish neurogenesis: Atlas. In Atlas of Early Zebrafish Brain Development; Academic Press: Cambridge, MA, USA, 2016; pp. 27–157. [Google Scholar]
- Kalueff, A.V.; Echevarria, D.J.; Stewart, A.M. Gaining translational momentum: More zebrafish models for neuroscience research. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 55, 1–6. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an emerging model for studying complex brain disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef]
- Kujawski, S.; Lin, W.; Kitte, F.; Börmel, M.; Fuchs, S.; Arulmozhivarman, G.; Vogt, S.; Theil, D.; Zhang, Y.; Antos, C.L. Calcineurin regulates coordinated outgrowth of zebrafish regenerating fins. Dev. Cell 2014, 28, 573–587. [Google Scholar] [CrossRef]
- Shapiro, R.; Fung, J.; Jain, A.; Parks, P.; Todo, S.; Starzl, T. The side effects of FK 506 in humans. Transplant. Proc. 1990, 22, 35–36. [Google Scholar] [PubMed]
- Porayko, M.K.; Textor, S.C.; Krom, R.A.; Hay, J.E.; Gores, G.J.; Richards, T.M.; Crotty, P.H.; Beaver, S.J.; Steers, J.L.; Wiesner, R.H. Nephrotoxic effects of primary immunosuppression with FK-506 and cyclosporine regimens after liver transplantation. Mayo Clin. Proc. 1994, 69, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Mueller, A.R.; Platz, K.P.; Schattenfroh, N.; Bechstein, W.O.; Christe, W.; Neuhaus, P.; Christe, W. Neurotoxicity after orthotopic liver transplantation in cyclosporin A-and FK 506-treated patients. Transpl. Int. 1994, 7, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Thiru, S.; Collier, D.; Calne, R. Pathological studies in canine and baboon renal allograft recipients immunosuppressed with FK-506. Transplant. Proc. 1987, 19, 98–99. [Google Scholar] [PubMed]
- Tiebosch, A.; Ericzon, B.; Wijnen, R.; Arends, J.; Groth, C.; Kootstra, G. Side effects of FK 506 in cynomolgus monkeys: A histopathologic study. Transplant. Proc. 1990, 22, 1652. [Google Scholar] [PubMed]
- Ochiai, T.; Hamaguchi, K.; Isono, K. Histopathologic studies in renal transplant recipient dogs receiving treatment with FK-506. Transplant. Proc. 1987, 19, 93–97. [Google Scholar] [PubMed]
- Giessler, G.A.; Gades, N.M.; Friedrich, P.F.; Bishop, A.T. Severe tacrolimus toxicity in rabbits. Exp. Clin. Transplant. 2007, 5, 590–595. [Google Scholar]
- Passos, E.; Genaidy, M.M.; Peyman, G.A. Ocular toxicity of intravitreal tacrolimus. Ophthalmic Surg. Lasers Imaging Retin. 2002, 33, 140–144. [Google Scholar] [CrossRef]
- Nalesnik, M.; Todo, S.; Murase, N.; Gryzan, S.; Lee, P.; Makowka, L.; Starzl, T. Toxicology of FK-506 in the Lewis rat. Transplant. Proc. 1987, 19, 89–92. [Google Scholar]
- Thomson, A.; Stephen, M.; Woo, J.; Hasan, N.; Whiting, P. Immunosuppressive activity, T-cell subset analysis, and acute toxicity of FK-506 in rats. Transplant. Proc. 1989, 21, 1048–1049. [Google Scholar]
- Avdesh, A.; Chen, M.; Martin-Iverson, M.T.; Mondal, A.; Ong, D.; Rainey-Smith, S.; Taddei, K.; Lardelli, M.; Groth, D.M.; Verdile, G.; et al. Regular care and maintenance of a zebrafish (Danio rerio) laboratory: An introduction. J. Vis. Exp. 2012, 18, e4196. [Google Scholar]
- Giebułtowicz, J.; Nałęcz-Jawecki, G. Occurrence of immunosuppressive drugs and their metabolites in the sewage-impacted Vistula and Utrata Rivers and in tap water from the Warsaw region (Poland). Chemosphere 2016, 148, 137–147. [Google Scholar] [CrossRef]
- Rakotobe, L.; Berkal, M.; Huet, H.; Djediat, C.; Jeannoda, V.; Bodo, B.; Mambu, L.; Crespeau, F.; Edery, M. Effects of Madagascar yam extracts, Dioscorea antaly, on embryo-larval development of medaka fish, Oryzias latipes. Toxicon 2010, 55, 87–91. [Google Scholar] [CrossRef]
- Christou, M.; Kavaliauskis, A.; Ropstad, E.; Fraser, T.W.K. DMSO effects larval zebrafish (Danio rerio) behavior, with additive and interaction effects when combined with positive controls. Sci. Total Environ. 2020, 709, 134490. [Google Scholar] [CrossRef] [PubMed]
- Hallare, A.; Nagel, K.; Köhler, H.-R.; Triebskorn, R. Comparative embryotoxicity and proteotoxicity of three carrier solvents to zebrafish (Danio rerio) embryos. Ecotoxicol. Environ. Saf. 2006, 63, 378–388. [Google Scholar] [CrossRef]
- Hoage, T.; Ding, Y.; Xu, X. Quantifying cardiac functions in embryonic and adult zebrafish. In Cardiovascular Development: Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; pp. 11–20. [Google Scholar]
- Saputra, F.; Lai, Y.-H.; Roldan, M.J.M.; Alos, H.C.; Aventurado, C.A.; Vasquez, R.D.; Hsiao, C.-D. The Effect of the Pyrethroid Pesticide Fenpropathrin on the Cardiac Performance of Zebrafish and the Potential Mechanism of Toxicity. Biology 2023, 12, 1214. [Google Scholar] [CrossRef]
- Bagatto, B.; Pelster, B.; Burggren, W. Growth and metabolism of larval zebrafish: Effects of swim training. J. Exp. Biol. 2001, 204, 4335–4343. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, G.; Murashita, K.; Verri, T.; Gomes, A.S.; Rønnestad, I. Leptin receptor-deficient (knockout) zebrafish: Effects on nutrient acquisition. Gen. Comp. Endocrinol. 2021, 310, 113832. [Google Scholar] [CrossRef]
- Kristofco, L.A.; Cruz, L.C.; Haddad, S.P.; Behra, M.L.; Chambliss, C.K.; Brooks, B.W. Age matters: Developmental stage of Danio rerio larvae influences photomotor response thresholds to diazinion or diphenhydramine. Aquat. Toxicol. 2016, 170, 344–354. [Google Scholar] [CrossRef]
- Siregar, P.; Audira, G.; Feng, L.-Y.; Lee, J.-H.; Santoso, F.; Yu, W.-H.; Lai, Y.-H.; Li, J.-H.; Lin, Y.-T.; Chen, J.-R.; et al. Pharmaceutical Assessment Suggests Locomotion Hyperactivity in Zebrafish Triggered by Arecoline Might Be Associated with Multiple Muscarinic Acetylcholine Receptors Activation. Toxins 2021, 13, 259. [Google Scholar] [CrossRef]
- Faria, M.; Prats, E.; Novoa-Luna, K.A.; Bedrossiantz, J.; Gómez-Canela, C.; Gómez-Oliván, L.M.; Raldúa, D. Development of a vibrational startle response assay for screening environmental pollutants and drugs impairing predator avoidance. Sci. Total Environ. 2019, 650, 87–96. [Google Scholar] [CrossRef]
- Pérez-Escudero, A.; Vicente-Page, J.; Hinz, R.C.; Arganda, S.; De Polavieja, G.G. idTracker: Tracking individuals in a group by automatic identification of unmarked animals. Nat. Methods 2014, 11, 743–748. [Google Scholar] [CrossRef]
- Audira, G.; Sampurna, B.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A versatile setup for measuring multiple behavior endpoints in zebrafish. Inventions 2018, 3, 75. [Google Scholar] [CrossRef]
- Yamanaka, O.; Takeuchi, R. UMATracker: An intuitive image-based tracking platform. J. Exp. Biol. 2018, 221, jeb182469. [Google Scholar] [CrossRef] [PubMed]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Han, L.; Hsiao, C.-D. Establishing simple image-based methods and cost-effective instrument for toxicity assessment on circadian rhythm dysregulation in fish. Biol. Open 2019, 8, 041871. [Google Scholar] [CrossRef]
- Siregar, P.; Juniardi, S.; Audira, G.; Lai, Y.-H.; Huang, J.-C.; Chen, K.H.-C.; Chen, J.-R.; Hsiao, C.-D. Method Standardization for Conducting Innate Color Preference Studies in Different Zebrafish Strains. Biomedicines 2020, 8, 271. [Google Scholar] [CrossRef] [PubMed]
- Sarasamma, S.; Audira, G.; Juniardi, S.; Sampurna, B.; Liang, S.-T.; Hao, E.; Lai, Y.-H.; Hsiao, C.-D. Zinc Chloride Exposure Inhibits Brain Acetylcholine Levels, Produces Neurotoxic Signatures, and Diminishes Memory and Motor Activities in Adult Zebrafish. Int. J. Mol. Sci. 2018, 19, 3195. [Google Scholar] [CrossRef]
- Adams, D.C.; Anthony, C.D. Using randomization techniques to analyse behavioural data. Anim. Behav. 1996, 51, 733–738. [Google Scholar] [CrossRef]
- Shelton, G.; Randall, D.J. The relationship between heart beat and respiration in teleost fish. Comp. Biochem. Physiol. 1962, 7, 237–250. [Google Scholar] [CrossRef]
- Stevens, E.D.; Randall, D. Changes in blood pressure, heart rate and breathing rate during moderate swimming activity in rainbow trout. J. Exp. Biol. 1967, 46, 307–315. [Google Scholar] [CrossRef]
- Shin, Y.J.; Lim, S.W.; Cui, S.; Ko, E.J.; Chung, B.H.; Kim, H.L.; Riew, T.R.; Lee, M.Y.; Yang, C.W. Tacrolimus decreases cognitive function by impairing hippocampal synaptic balance: A possible role of Klotho. Mol. Neurobiol. 2021, 58, 5954–5970. [Google Scholar] [CrossRef]
- Grillner, S.; Markram, H.; De Schutter, E.; Silberberg, G.; LeBeau, F.E. Microcircuits in action–from CPGs to neocortex. Trends Neurosci. 2005, 28, 525–533. [Google Scholar] [CrossRef]
- Horzmann, K.A.; Freeman, J.L. Zebrafish get connected: Investigating neurotransmission targets and alterations in chemical toxicity. Toxics 2016, 4, 19. [Google Scholar] [CrossRef]
- Ahmad, F.; Noldus, L.P.; Tegelenbosch, R.A.; Richardson, M.K. Zebrafish embryos and larvae in behavioural assays. Behaviour 2012, 149, 1241. [Google Scholar]
- Basnet, R.M.; Zizioli, D.; Taweedet, S.; Finazzi, D.; Memo, M. Zebrafish larvae as a behavioral model in neuropharmacology. Biomedicines 2019, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, A.; Sun, Y.; Cao, X.; Zhang, N. Treatment with immunosuppressants FTY720 and tacrolimus promotes functional recovery after spinal cord injury in rats. Tohoku J. Exp. Med. 2009, 219, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Sasakawa, T.; Sasakawa, Y.; Ohkubo, Y.; Mutoh, S. FK506 ameliorates spontaneous locomotor activity in collagen-induced arthritis: Implication of distinct effect from suppression of inflammation. Int. Immunopharmacol. 2005, 5, 503–510. [Google Scholar] [CrossRef]
- Sakanoue, M.; Mori, N.; Takei, N.; Kawai, M.; Tani, K.; Suzuki, K.; Iwata, Y.; Sekine, Y.; Ashby, C.R., Jr.; Minabe, Y. Tacrolimus, a specific inhibitor of calcineurin, modifies the locomotor activity of quinpirole, but not that of SKF82958, in male rats. Eur. J. Pharmacol. 2002, 438, 93–97. [Google Scholar] [CrossRef]
- Shin, Y.J.; Chun, Y.T.; Lim, S.W.; Luo, K.; Quan, Y.; Cui, S.; Ko, E.J.; Chung, B.H.; Lee, J.; Hong, S.; et al. Influence of tacrolimus on depressive-like behavior in diabetic rats through brain-derived neurotrophic factor regulation in the hippocampus. Neurotox. Res. 2019, 36, 396–410. [Google Scholar] [CrossRef]
- Maximino, C.; de Oliveira, D.L.; Rosemberg, D.B.; Batista, E.d.J.O.; Herculano, A.M.; Oliveira, K.R.M.; Benzecry, R.; Blaser, R. A comparison of the light/dark and novel tank tests in zebrafish. Behaviour 2012, 149, 1099–1123. [Google Scholar] [CrossRef]
- Blaser, R.E.; Rosemberg, D.B. Measures of anxiety in zebrafish (Danio rerio): Dissociation of black/white preference and novel tank test. PLoS ONE 2012, 7, e36931. [Google Scholar] [CrossRef]
- Cachat, J.; Stewart, A.; Grossman, L.; Gaikwad, S.; Kadri, F.; Chung, K.M.; Wu, N.; Wong, K.; Roy, S.; Suciu, C.; et al. Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Nat. Protoc. 2010, 5, 1786–1799. [Google Scholar] [CrossRef]
- Meijide, F.J.; Da Cuña, R.H.; Prieto, J.P.; Dorelle, L.S.; Babay, P.A.; Nostro, F.L.L. Effects of waterborne exposure to the antidepressant fluoxetine on swimming, shoaling and anxiety behaviours of the mosquitofish Gambusia holbrooki. Ecotoxicol. Environ. Saf. 2018, 163, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Grossman, L.; Utterback, E.; Stewart, A.; Gaikwad, S.; Chung, K.M.; Suciu, C.; Wong, K.; Elegante, M.; Elkhayat, S.; Tan, J.; et al. Characterization of behavioral and endocrine effects of LSD on zebrafish. Behav. Brain Res. 2010, 214, 277–284. [Google Scholar] [CrossRef]
- Hamilton, T.J.; Krook, J.; Szaszkiewicz, J.; Burggren, W. Shoaling, boldness, anxiety-like behavior and locomotion in zebrafish (Danio rerio) are altered by acute benzo[a]pyrene exposure. Sci. Total Environ. 2021, 774, 145702. [Google Scholar] [CrossRef]
- Johnson, A.; Loh, E.; Verbitsky, R.; Slessor, J.; Franczak, B.C.; Schalomon, M.; Hamilton, T.J. Examining behavioural test sensitivity and locomotor proxies of anxiety-like behaviour in zebrafish. Sci. Rep. 2023, 13, 3768. [Google Scholar] [CrossRef] [PubMed]
- Sackerman, J.; Donegan, J.J.; Cunningham, C.S.; Nguyen, N.N.; Lawless, K.; Long, A.; Benno, R.H.; Gould, G.G. Zebrafish behavior in novel environments: Effects of acute exposure to anxiolytic compounds and choice of Danio rerio line. Int. J. Comp. Psychol./ISCP Spons. Int. Soc. Comp. Psychol. Univ. Calabr. 2010, 23, 43. [Google Scholar]
- Maximino, C.; Marques de Brito, T.; Dias, C.A.G.d.M.; Gouveia, A., Jr.; Morato, S. Scototaxis as anxiety-like behavior in fish. Nat. Protoc. 2010, 5, 209–216. [Google Scholar] [CrossRef]
- Foldvik, A.; Einum, S.; Finstad, A.G. Spatial diffusion modelling of juvenile Atlantic salmon (Salmo salar) shows ontogenetic increase in movement rates. Can. J. Fish. Aquat. Sci. 2017, 74, 202–207. [Google Scholar] [CrossRef]
- Werner, E.E.; Gilliam, J.F. The ontogenetic niche and species interactions in size-structured populations. Annu. Rev. Ecol. Syst. 1984, 15, 393–425. [Google Scholar] [CrossRef]
- Landgraf, D.; Long, J.E.; Proulx, C.D.; Barandas, R.; Malinow, R.; Welsh, D.K. Genetic disruption of circadian rhythms in the suprachiasmatic nucleus causes helplessness, behavioral despair, and anxiety-like behavior in mice. Biol. Psychiatry 2016, 80, 827–835. [Google Scholar] [CrossRef]
- Kalmbach, D.A.; Pillai, V.; Cheng, P.; Arnedt, J.T.; Drake, C.L. Shift work disorder, depression, and anxiety in the transition to rotating shifts: The role of sleep reactivity. Sleep Med. 2015, 16, 1532–1538. [Google Scholar] [CrossRef]
- Roybal, K.; Theobold, D.; Graham, A.; DiNieri, J.A.; Russo, S.J.; Krishnan, V.; Chakravarty, S.; Peevey, J.; Oehrlein, N.; Birnbaum, S.; et al. Mania-like behavior induced by disruption of CLOCK. Proc. Natl. Acad. Sci. USA 2007, 104, 6406–6411. [Google Scholar] [CrossRef] [PubMed]
- Germain, A.; Kupfer, D.J. Circadian rhythm disturbances in depression. Hum. Psychopharmacol. Clin. Exp. 2008, 23, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Hühne, A.; Volkmann, P.; Stephan, M.; Rossner, M.; Landgraf, D. An in-depth neurobehavioral characterization shows anxiety-like traits, impaired habituation behavior, and restlessness in male Cryptochrome-deficient mice. Genes Brain Behav. 2020, 19, e12661. [Google Scholar] [CrossRef] [PubMed]
- Reebs, S.G. Plasticity of diel and circadian activity rhythms in fishes. Rev. Fish Biol. Fish. 2002, 12, 349–371. [Google Scholar] [CrossRef]
- Pinheiro-da-Silva, J.; Tran, S.; Luchiari, A.C. Sleep deprivation impairs cognitive performance in zebrafish: A matter of fact? Behav. Process. 2018, 157, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Miner, K.A.; Huertas, M.; Aspbury, A.S.; Gabor, C.R. Artificial light at night alters the physiology and behavior of western mosquitofish (Gambusia affinis). Front. Ecol. Evol. 2021, 9, 617063. [Google Scholar] [CrossRef]
- Merola, C.; Lucon-Xiccato, T.; Bertolucci, C.; Perugini, M. Behavioural effects of early-life exposure to parabens in zebrafish larvae. J. Appl. Toxicol. 2021, 41, 1852–1862. [Google Scholar] [CrossRef]
- Kim, J.L.; Kim, S.S.; Hwang, K.-S.; Park, H.-C.; Cho, S.-H.; Bae, M.A.; Kim, K.-T. Chronic exposure to butyl-paraben causes photosensitivity disruption and memory impairment in adult zebrafish. Aquat. Toxicol. 2022, 251, 106279. [Google Scholar] [CrossRef]
- Ruchin, A.B. Environmental colour impact on the life of lower aquatic vertebrates: Development, growth, physiological and biochemical processes. Rev. Aquac. 2020, 12, 310–327. [Google Scholar] [CrossRef]
- Maia, C.M.; Volpato, G.L. Environmental light color affects the stress response of Nile tilapia. Zoology 2013, 116, 64–66. [Google Scholar] [CrossRef]
- Peeters, B.W.; Moeskops, M.; Veenvliet, A.R. Color preference in Danio rerio: Effects of age and anxiolytic treatments. Zebrafish 2016, 13, 330–334. [Google Scholar] [CrossRef]
- Neuhauss, S.C. Behavioral genetic approaches to visual system development and function in zebrafish. J. Neurobiol. 2003, 54, 148–160. [Google Scholar] [CrossRef]
- Poisson, A.; Valotaire, C.; Borel, F.; Bertin, A.; Darmaillacq, A.-S.; Dickel, L.; Colson, V. Embryonic exposure to a conspecific alarm cue triggers behavioural plasticity in juvenile rainbow trout. Anim. Behav. 2017, 133, 35–45. [Google Scholar] [CrossRef]
- Colson, V.; Ferreira, V.H.B.; Luchiari, A.C.; Valotaire, C.; Borel, F.; Bugeon, J.; Prigent, S.; Dickel, L.; Calandreau, L.; Guesdon, V. Loss of light colour preference after chronic embryonic stress in rainbow trout fry: A novel and potential indicator of fish welfare? Appl. Anim. Behav. Sci. 2021, 239, 105335. [Google Scholar] [CrossRef]
- Jia, L.; Raghupathy, R.K.; Albalawi, A.; Zhao, Z.; Reilly, J.; Xiao, Q.; Shu, X. A colour preference technique to evaluate acrylamide-induced toxicity in zebrafish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 199, 11–19. [Google Scholar] [CrossRef]
- Wong, K.; Elegante, M.; Bartels, B.; Elkhayat, S.; Tien, D.; Roy, S.; Goodspeed, J.; Suciu, C.; Tan, J.; Grimes, C.; et al. Analyzing habituation responses to novelty in zebrafish (Danio rerio). Behav. Brain Res. 2010, 208, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Sison, M.; Gerlai, R. Associative learning in zebrafish (Danio rerio) in the plus maze. Behav. Brain Res. 2010, 207, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Boeve, B.F.; Kimmel, D.W.; Aronson, A.E.; De Groen, P.C. Dysarthria and apraxia of speech associated with FK-506 (tacrolimus). Mayo Clin. Proc. 1996, 71, 969–972. [Google Scholar] [CrossRef] [PubMed]
- Benetoli, A.; Paganelli, R.A.; Giordani, F.o.; Lima, K.C.M.; Fávero Filho, L.A.; Milani, H. Effect of tacrolimus (FK506) on ischemia-induced brain damage and memory dysfunction in rats. Pharmacol. Biochem. Behav. 2004, 77, 607–615. [Google Scholar] [CrossRef]
- Ganesh, L.; Deepa, B.; Holla, R.; Manikkoth, S. Role of tacrolimus in alcohol-induced cognitive impairment in Wistar albino rats. Drug Invent. Today 2020, 14, 1201. [Google Scholar]
- Lee, S.-H.; Kim, B.-C.; Yang, D.-H.; Park, M.-S.; Choi, S.-M.; Kim, M.-K.; Cho, K.-H. Calcineurin inhibitormediated bilateral hippocampal injury after bone marrow transplantation. J. Neurol. 2008, 255, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Wijdicks, E.F.; Wiesner, R.H.; Dahlke, L.J.; Krom, R.A. FK506-induced neurotoxicity in liver transplantation. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 1994, 35, 498–501. [Google Scholar] [CrossRef] [PubMed]
- Hadjiasgary, A.; Banafshe, H.R.; Ardjmand, A. Intra-CA1 administration of FK-506 (tacrolimus) in rat impairs learning and memory in an inhibitory avoidance paradigm. Iran. J. Basic Med. Sci. 2015, 18, 130. [Google Scholar] [PubMed]
- Gaikwad, S.; Stewart, A.; Hart, P.; Wong, K.; Piet, V.; Cachat, J.; Kalueff, A.V. Acute stress disrupts performance of zebrafish in the cued and spatial memory tests: The utility of fish models to study stress–memory interplay. Behav. Process. 2011, 87, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Harding, M.W.; Galat, A.; Uehling, D.E.; Schreiber, S.L. A receptor for the immuno-suppressant FK506 is a cis–trans peptidyl-prolyl isomerase. Nature 1989, 341, 758–760. [Google Scholar] [CrossRef] [PubMed]
- Siekierka, J.J.; Hung, S.H.; Poe, M.; Lin, C.S.; Sigal, N.H. A cytosolic binding protein for the immunosuppressant FK506 has peptidyl-prolyl isomerase activity but is distinct from cyclophilin. Nature 1989, 341, 755–757. [Google Scholar] [CrossRef]
- Galat, A. Peptidylprolyl cis/trans isomerases (immunophilins): Biological diversity-targets-functions. Curr. Top. Med. Chem. 2003, 3, 1315–1347. [Google Scholar] [CrossRef]
- Huai, Q.; Kim, H.-Y.; Liu, Y.; Zhao, Y.; Mondragon, A.; Liu, J.O.; Ke, H. Crystal structure of calcineurin–cyclophilin–cyclosporin shows common but distinct recognition of immunophilin–drug complexes. Proc. Natl. Acad. Sci. USA 2002, 99, 12037–12042. [Google Scholar] [CrossRef]
- Ke, H.; Huai, Q. Structures of calcineurin and its complexes with immunophilins–immunosuppressants. Biochem. Biophys. Res. Commun. 2003, 311, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Kumer, A.; Zaben, F.; Khan, M.W. Investigating the binding affinity, molecular dynamics, and ADMET properties of 2, 3-dihydrobenzofuran derivatives as an inhibitor of fungi, bacteria, and virus protein. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 36. [Google Scholar] [CrossRef]
- Dawson, T.; Steiner, J.P.; Lyons, W.; Fotuhi, M.; Blue, M.; Snyder, S. The immunophilins, FK506 binding protein and cyclophilin, are discretely localized in the brain: Relationship to calcineurin. Neuroscience 1994, 62, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.B.; Hong, Y.; Dhe-Paganon, S.; Yoon, H.S. FKBP family proteins: Immunophilins with versatile biological functions. Neurosignals 2008, 16, 318–325. [Google Scholar] [CrossRef] [PubMed]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, W.-W.; Chen, H.-C.; Audira, G.; Suryanto, M.E.; Saputra, F.; Kurnia, K.A.; Vasquez, R.D.; Casuga, F.P.; Lai, Y.-H.; Hsiao, C.-D.; et al. Evaluation of Tacrolimus’ Adverse Effects on Zebrafish in Larval and Adult Stages by Using Multiple Physiological and Behavioral Endpoints. Biology 2024, 13, 112. https://doi.org/10.3390/biology13020112
Feng W-W, Chen H-C, Audira G, Suryanto ME, Saputra F, Kurnia KA, Vasquez RD, Casuga FP, Lai Y-H, Hsiao C-D, et al. Evaluation of Tacrolimus’ Adverse Effects on Zebrafish in Larval and Adult Stages by Using Multiple Physiological and Behavioral Endpoints. Biology. 2024; 13(2):112. https://doi.org/10.3390/biology13020112
Chicago/Turabian StyleFeng, Wen-Wei, Hsiu-Chao Chen, Gilbert Audira, Michael Edbert Suryanto, Ferry Saputra, Kevin Adi Kurnia, Ross D. Vasquez, Franelyne P. Casuga, Yu-Heng Lai, Chung-Der Hsiao, and et al. 2024. "Evaluation of Tacrolimus’ Adverse Effects on Zebrafish in Larval and Adult Stages by Using Multiple Physiological and Behavioral Endpoints" Biology 13, no. 2: 112. https://doi.org/10.3390/biology13020112
APA StyleFeng, W.-W., Chen, H.-C., Audira, G., Suryanto, M. E., Saputra, F., Kurnia, K. A., Vasquez, R. D., Casuga, F. P., Lai, Y.-H., Hsiao, C.-D., & Hung, C.-H. (2024). Evaluation of Tacrolimus’ Adverse Effects on Zebrafish in Larval and Adult Stages by Using Multiple Physiological and Behavioral Endpoints. Biology, 13(2), 112. https://doi.org/10.3390/biology13020112