
Can Extensive Training Transform a Mouse into a Guinea Pig? An Evaluation Based on the Discriminative Abilities of Inferior Colliculus Neurons


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
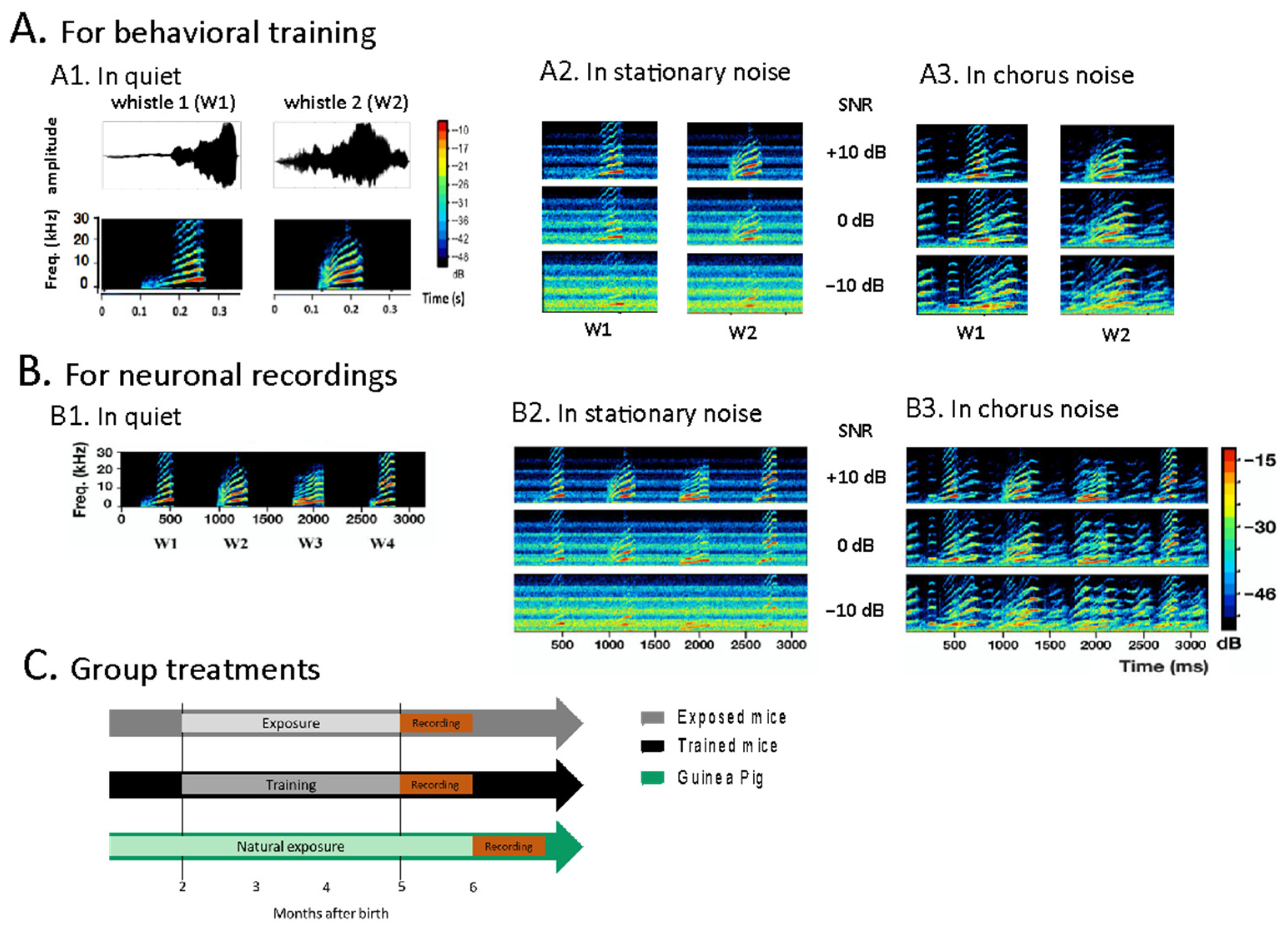
2.2. Behavioral Training
2.3. Recording Procedures
2.4. Experimental Protocol during Neuronal Recordings
2.5. Data Analysis
2.5.1. The Quantification of Responses to Pure Tones
2.5.2. The Quantification of Evoked Responses to the Four Vocalizations
2.5.3. The Quantification of Mutual Information from the Responses to Vocalizations
2.6. Statistical Analysis
3. Results
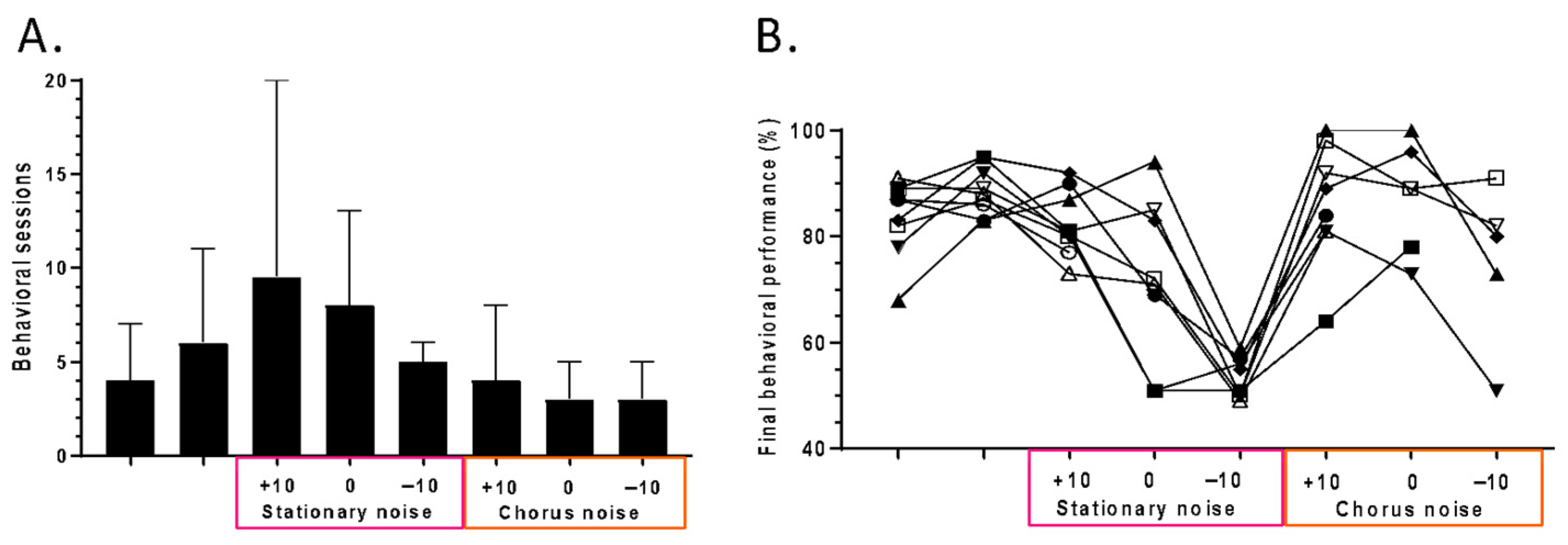
3.1. The Behavioral Performance of the Trained Mice in Quiet and in Noisy Conditions
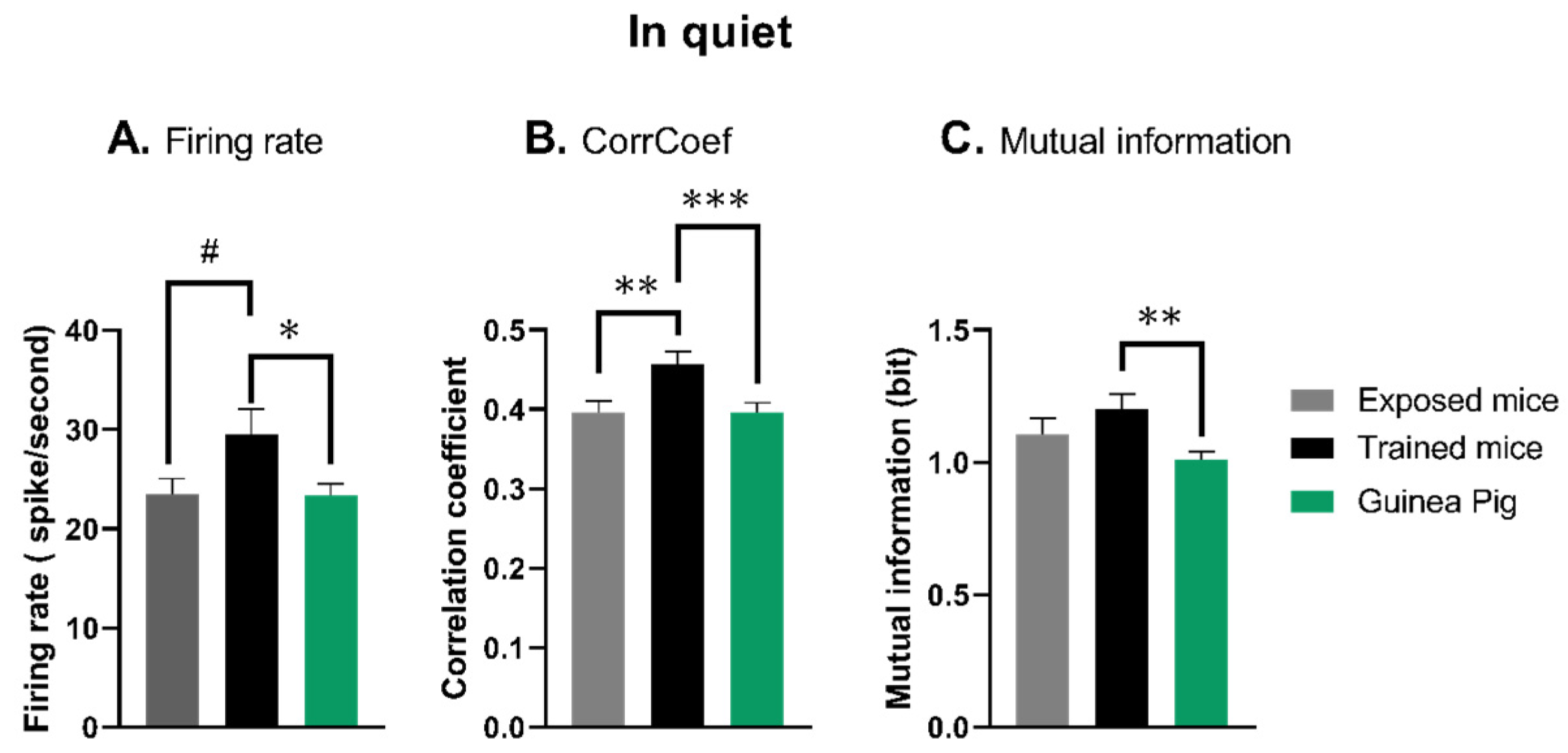
3.2. The Quantification of Neuronal Responses in Anesthetized Mice and Guinea Pigs in Quiet Conditions
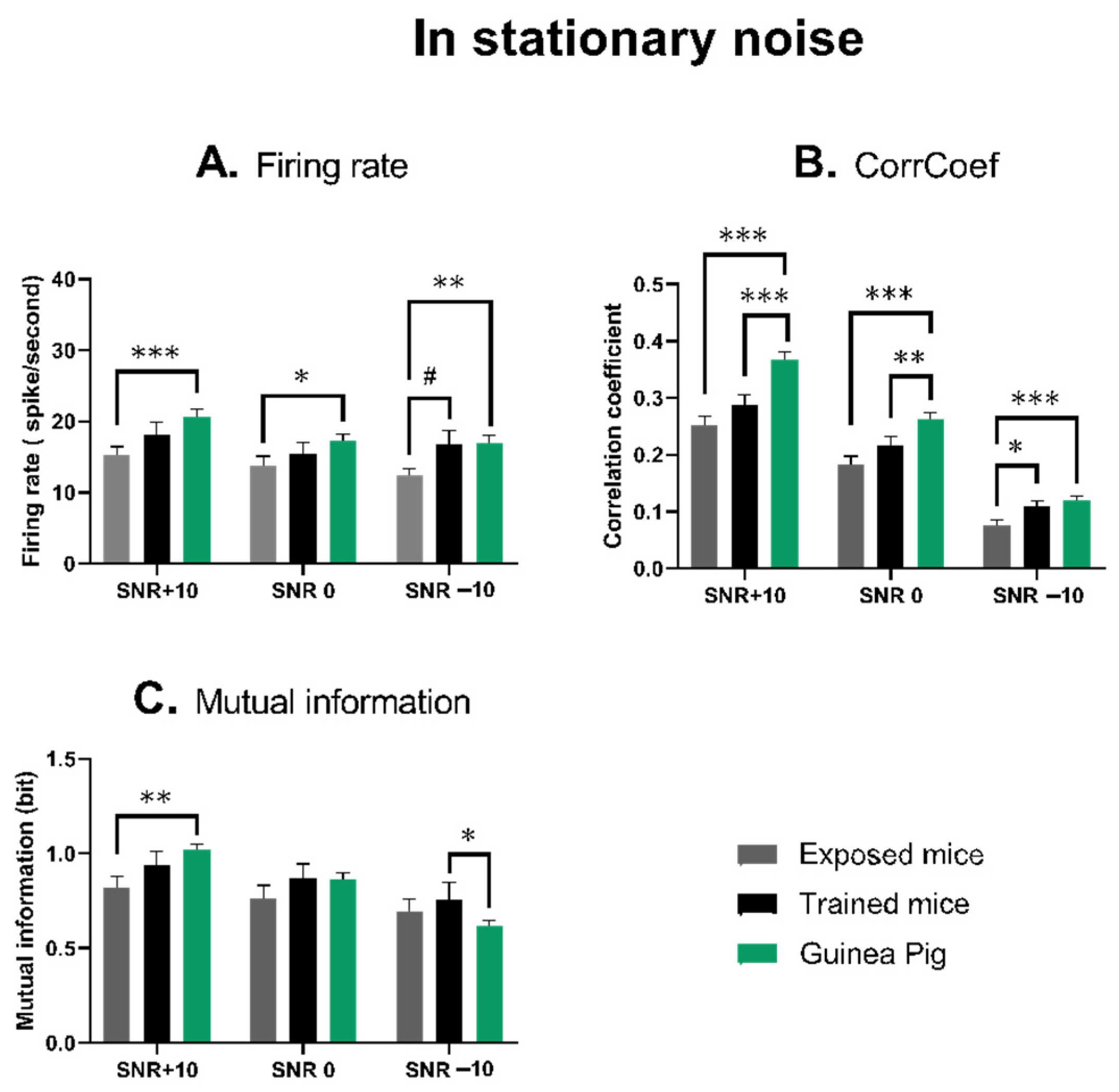
3.3. The Quantification of the Neuronal Responses in Stationary Noise
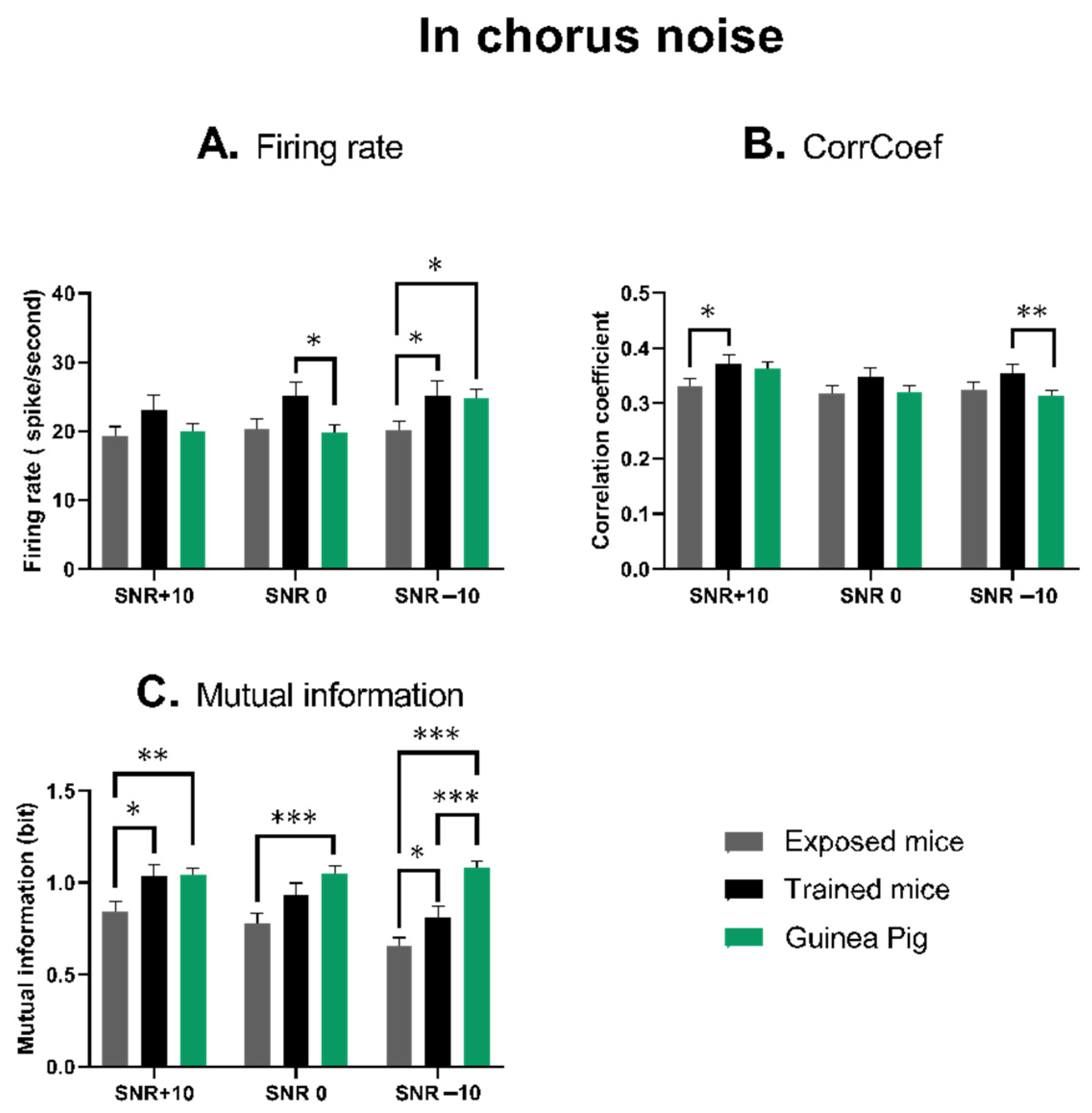
3.4. The Quantification of the Neuronal Responses in Chorus Noise
4. Discussion
4.1. Behavioral Training with Conspecific or Heterospecific Communication Sounds
4.2. The Neuronal Consequences of Training with Heterospecific Vocalizations
4.3. The Potential Mechanisms of This Neuronal Plasticity Occurring at the IC Level
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagarajan, S.S.; Cheung, S.W.; Bedenbaugh, P.; Beitel, R.E.; Schreiner, C.E.; Merzenich, M.M. Representation of Spectral and Temporal Envelope of Twitter Vocalizations in Common Marmoset Primary Auditory Cortex. J. Neurophysiol. 2002, 87, 1723–1737. [Google Scholar] [CrossRef] [PubMed]
- Narayan, R.; Best, V.; Ozmeral, E.; McClaine, E.; Dent, M.; Shinn-Cunningham, B.; Sen, K. Cortical interference effects in the cocktail party problem. Nat. Neurosci. 2007, 10, 1601–1607. [Google Scholar] [CrossRef]
- Schneider, D.M.; Woolley, S.M.N. Sparse and Background-Invariant Coding of Vocalizations in Auditory Scenes. Neuron 2013, 79, 141–152. [Google Scholar] [CrossRef]
- Carruthers, I.M.; Laplagne, D.A.; Jaegle, A.; Briguglio, J.J.; Mwilambwe-Tshilobo, L.; Natan, R.G.; Geffen, M.N. Emergence of invariant representation of vocalizations in the auditory cortex. J. Neurophysiol. 2015, 114, 2726–2740. [Google Scholar] [CrossRef] [PubMed]
- Ni, R.; Bender, D.A.; Shanechi, A.M.; Gamble, J.R.; Barbour, D.L. Contextual effects of noise on vocalization encoding in primary auditory cortex. J. Neurophysiol. 2017, 117, 713–727. [Google Scholar] [CrossRef]
- Town, S.M.; Wood, K.C.; Bizley, J.K. Sound identity is represented robustly in auditory cortex during perceptual constancy. Nat. Commun. 2018, 9, 4786. [Google Scholar] [CrossRef]
- Shetake, J.A.; Wolf, J.T.; Cheung, R.J.; Engineer, C.T.; Ram, S.K.; Kilgard, M.P. Cortical activity patterns predict robust speech discrimination ability in noise. Eur. J. Neurosci. 2011, 34, 1823–1838. [Google Scholar] [CrossRef] [PubMed]
- Souffi, S.; Varnet, L.; Lorenzi, C.; Huetz, C.; Edeline, J.M. Noise-sensitive but more precise subcortical representations co-exist with robust cortical encoding of natural vocalizations. J. Neurosci. 2020, 40, 5228–5246. [Google Scholar] [CrossRef]
- Souffi, S.; Varnet, L.; Zaidi, M.; Bathellier, B.; Huetz, C.; Edeline, J.M. Reduction in sound discrimination in noise is related to envelope similarity and not to a decrease in envelope tracking abilities. J. Physiol. 2023, 601, 123–149. [Google Scholar] [CrossRef] [PubMed]
- Schnupp, J.W.; Hall, T.M.; Kokelaar, R.F.; Ahmed, B. Plasticity of temporal pattern codes for vocalization stimuli in primary auditory cortex. J. Neurosci. 2006, 26, 4785–4795. [Google Scholar] [CrossRef]
- Deneux, T.; Kempf, A.; Daret, A.; Ponsot, E.; Bathellier, B. Temporal asymmetries in auditory coding and perception reflect multi-layered nonlinearities. Nat. Commun. 2016, 7, 12682. [Google Scholar] [CrossRef] [PubMed]
- Ceballo, S.; Piwkowska, Z.; Bourg, J.; Daret, A.; Bathellier, B. Targeted Cortical Manipulation of Auditory Perception. Neuron 2019, 104, 1168–1179.e5. [Google Scholar] [CrossRef] [PubMed]
- Bathellier, B.; Ushakova, L.; Rumpel, S. Discrete neocortical dynamics predict behavioral categorization of sounds. Neuron 2012, 76, 435–449. [Google Scholar] [CrossRef]
- Souffi, S.; Lorenzi, C.; Huetz, C.; Edeline, J.M. Robustness to noise in the auditory system: A distributed and predictable property. eNeuro 2021, 8, ENEURO.0043-21.2021. [Google Scholar] [CrossRef] [PubMed]
- Franke, F.; Quian Quiroga, R.; Hierlemann, A.; Obermayer, K. Bayes optimal template matching for spike sorting—Combining fisher discriminant analysis with optimal filtering. J. Comput. Neurosci. 2015, 38, 439–459. [Google Scholar] [CrossRef]
- Pouzat, C.; Delescluse, M.; Viot, P.; Diebolt, J. Improved spike-sorting by modeling firing statistics and burst-dependent spike amplitude attenuation: A Markov chain Monte Carlo approach. J. Neurophysiol. 2004, 91, 2910–2928. [Google Scholar] [CrossRef]
- Quiroga, R.Q.; Nadasdy, Z.; Ben-Shaul, Y. Unsupervised spike detection and sorting with wavelets and superparamagnetic clustering. Neural Comput. 2004, 16, 1661–1687. [Google Scholar] [CrossRef]
- Gaucher, Q.; Huetz, C.; Gourévitch, B.; Edeline, J.M. Cortical inhibition reduces information redundancy at presentation of communication sounds in the primary auditory cortex. J. Neurosci. 2013, 33, 10713–10728. [Google Scholar] [CrossRef]
- Neilans, E.G.; Holfoth, D.P.; Radziwon, K.E.; Portfors, C.V.; Dent, M.L. Discrimination of ultrasonic vocalizations by CBA/CaJ mice (Mus musculus) is related to spectrotemporal dissimilarity of vocalizations. PLoS ONE 2014, 9, e85405. [Google Scholar] [CrossRef]
- Perrodin, C.; Verzat, C.; Bendor, D. Courtship behaviour reveals temporal regularity is a critical social cue in mouse communication. eLife 2023, 12, RP86464. [Google Scholar] [CrossRef]
- Engineer, C.T.; Perez, C.A.; Chen, Y.H.; Carraway, R.S.; Reed, A.C.; Shetake, J.A.; Jakkamsetti, V.; Chang, K.Q.; Kilgard, M.P. Cortical activity patterns predict speech discrimination ability. Nat. Neurosci. 2008, 11, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Engineer, N.D.; Engineer, C.T.; Reed, A.C.; Pandya, P.K.; Jakkamsetti, V.; Moucha, R.; Kilgard, M.P. Inverted-U function relating cortical plasticity and task difficulty. Neuroscience 2012, 205, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, K.G.; Carraway, R.S.; Borland, M.S.; Moreno, N.A.; Hanacik, E.A.; Miller, R.S.; Kilgard, M.P. Speech discrimination after early exposure to pulsed-noise or speech. Hear. Res. 2012, 289, 1–12. [Google Scholar] [CrossRef]
- Ranasinghe, K.G.; Vrana, W.A.; Matney, C.J.; Kilgard, M.P. Neural mechanisms supporting robust discrimination of spectrally and temporally degraded speech. J. Assoc. Res. Otolaryngol. 2012, 13, 527–542. [Google Scholar] [CrossRef]
- von Trapp, G.; Buran, B.N.; Sen, K.; Semple, M.N.; Sanes, D.H. A Decline in Response Variability Improves Neural Signal Detection during Auditory Task Performance. J. Neurosci. 2016, 36, 11097–11106. [Google Scholar] [CrossRef] [PubMed]
- Olds, J.; Nienhuis, R.; Olds, M.E. Patterns of conditioned unit responses in the auditory system of the rat. Exp. Neurol. 1978, 59, 209–228. [Google Scholar] [CrossRef] [PubMed]
- Nienhuis, R.; Olds, J. Changes in unit responses to tones after food reinforcement in the auditory pathway of the rat: Intertrial arousal. Exp. Neurol. 1978, 59, 229–242. [Google Scholar] [CrossRef]
- Birt, D.; Nienhuis, R.; Olds, J. Effects of bilateral auditory cortex ablation on behavior and unit activity in rat inferior colliculus during differential conditioning. J. Neurophysiol. 1978, 41, 705–715. [Google Scholar] [CrossRef]
- Oleson, T.D.; Ashe, J.H.; Weinberger, N.M. Modification of auditory and somatosensory system activity during pupillary conditioning in the paralyzed cat. J. Neurophysiol. 1975, 38, 1114–1139. [Google Scholar] [CrossRef]
- Edeline, J.-M.; Neuenschwander-El Massioui, N.; Dutrieux, G. Frequency-specific cellular changes in the auditory system during acquisition and reversal of discriminative conditioning. Psychobiology 1990, 18, 382–393. [Google Scholar] [CrossRef]
- Bajo, V.M.; Nodal, F.R.; Moore, D.R.; King, A.J. The descending corticocollicular pathway mediates learning-induced auditory plasticity. Nat. Neurosci. 2010, 13, 253–260. [Google Scholar] [CrossRef]
- Bajo, V.M.; Nodal, F.R.; Korn, C.; Constantinescu, A.O.; Mann, E.O.; Boyden, E.S., 3rd; King, A.J. Silencing cortical activity during sound-localization training impairs auditory perceptual learning. Nat. Commun. 2019, 10, 3075. [Google Scholar] [CrossRef] [PubMed]
- Souffi, S.; Nodal, F.R.; Bajo, V.M.; Edeline, J.M. When and How Does the Auditory Cortex Influence Subcortical Auditory Structures? New Insights About the Roles of Descending Cortical Projections. Front. Neurosci. 2021, 15, 690223. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, J.M.; Lesicko, A.M.; Rao, W.; De Biasi, M.; Geffen, M.N. Auditory cortex shapes sound responses in the inferior colliculus. eLife 2020, 9, e51890. [Google Scholar] [CrossRef] [PubMed]
- Gittelman, J.X.; Perkel, D.J.; Portfors, C.V. Dopamine modulates auditory responses in the inferior colliculus in a heterogeneous manner. J. Assoc. Res. Otolaryngol. 2013, 14, 719–729. [Google Scholar] [CrossRef]
- Hoyt, J.M.; Perkel, D.J.; Portfors, C.V. Dopamine Acts via D2-Like Receptors to Modulate Auditory Responses in the Inferior Colliculus. eNeuro. 2019, 6, ENEURO.0350-19.2019. [Google Scholar] [CrossRef]
- Valdés-Baizabal, C.; Carbajal, G.V.; Pérez-González, D.; Malmierca, M.S. Dopamine modulates subcortical responses to surprising sounds. PLoS Biol. 2020, 18, e3000744. [Google Scholar] [CrossRef]
- Bao, S.; Chan, V.T.; Merzenich, M.M. Cortical remodelling induced by activity of ventral tegmental dopamine neurons. Nature 2001, 412, 79–83. [Google Scholar] [CrossRef]
- Edeline, J.-M. Beyond traditional approaches to understanding the functional role of neuromodulators in sensory cortices. Front. Behav. Neurosci. 2012, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- Gurden, H.; Takita, M.; Jay, T.M. Essential role of D1 but not D2 receptors in the NMDA receptor-dependent long-term potentiation at hippocampal-prefrontal cortex synapses in vivo. J. Neurosci. 2000, 20, RC106. [Google Scholar] [CrossRef] [PubMed]
- Otmakhova, N.A.; Lisman, J.E. D1/D5 dopamine receptors inhibit depotentiation at CA1 synapses via cAMP-dependent mechanism. J. Neurosci. 1998, 18, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, S.H. Long-term potentiation in the inferior colliculus studied in rat brain slice. Hear. Res. 2000, 147, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Ma, C.L.; Sivaramakrishnan, S.; Oliver, D.L. Synaptic modification in neurons of the central nucleus of the inferior colliculus. Hear. Res. 2002, 168, 43–54. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin, A.; Souffi, S.; Huetz, C.; Edeline, J.-M. Can Extensive Training Transform a Mouse into a Guinea Pig? An Evaluation Based on the Discriminative Abilities of Inferior Colliculus Neurons. Biology 2024, 13, 92. https://doi.org/10.3390/biology13020092
Martin A, Souffi S, Huetz C, Edeline J-M. Can Extensive Training Transform a Mouse into a Guinea Pig? An Evaluation Based on the Discriminative Abilities of Inferior Colliculus Neurons. Biology. 2024; 13(2):92. https://doi.org/10.3390/biology13020092
Chicago/Turabian StyleMartin, Alexandra, Samira Souffi, Chloé Huetz, and Jean-Marc Edeline. 2024. "Can Extensive Training Transform a Mouse into a Guinea Pig? An Evaluation Based on the Discriminative Abilities of Inferior Colliculus Neurons" Biology 13, no. 2: 92. https://doi.org/10.3390/biology13020092
APA StyleMartin, A., Souffi, S., Huetz, C., & Edeline, J. -M. (2024). Can Extensive Training Transform a Mouse into a Guinea Pig? An Evaluation Based on the Discriminative Abilities of Inferior Colliculus Neurons. Biology, 13(2), 92. https://doi.org/10.3390/biology13020092