
CCL28: A Promising Biomarker for Assessing Salivary Gland Functionality and Maintaining Healthy Oral Environments
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Histology and Immunofluorescence Staining
2.3. RT-PCR
2.4. Antibodies
2.5. Statistical Analysis
3. Results
3.1. Deletion of CCL28 Reduces Acidic Mucin Production in Both SMG and SLG and Promotes Bacterial Invasion
3.2. Deletion of CCL28 Diminishes the Expression of Salivary Gland Markers
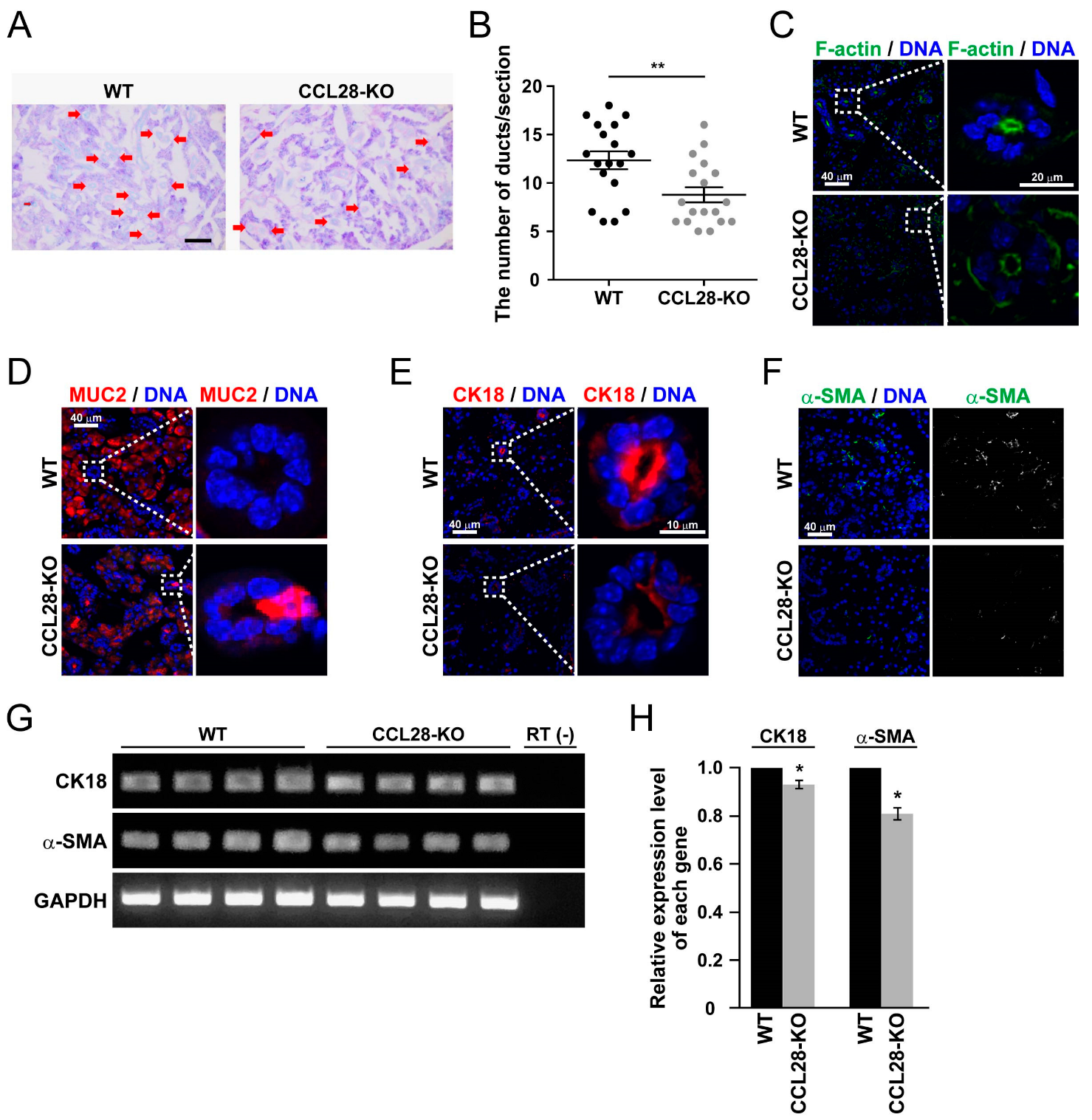
3.3. Deletion of CCL28 Causes Alterations in the Ductal Structure of SMG
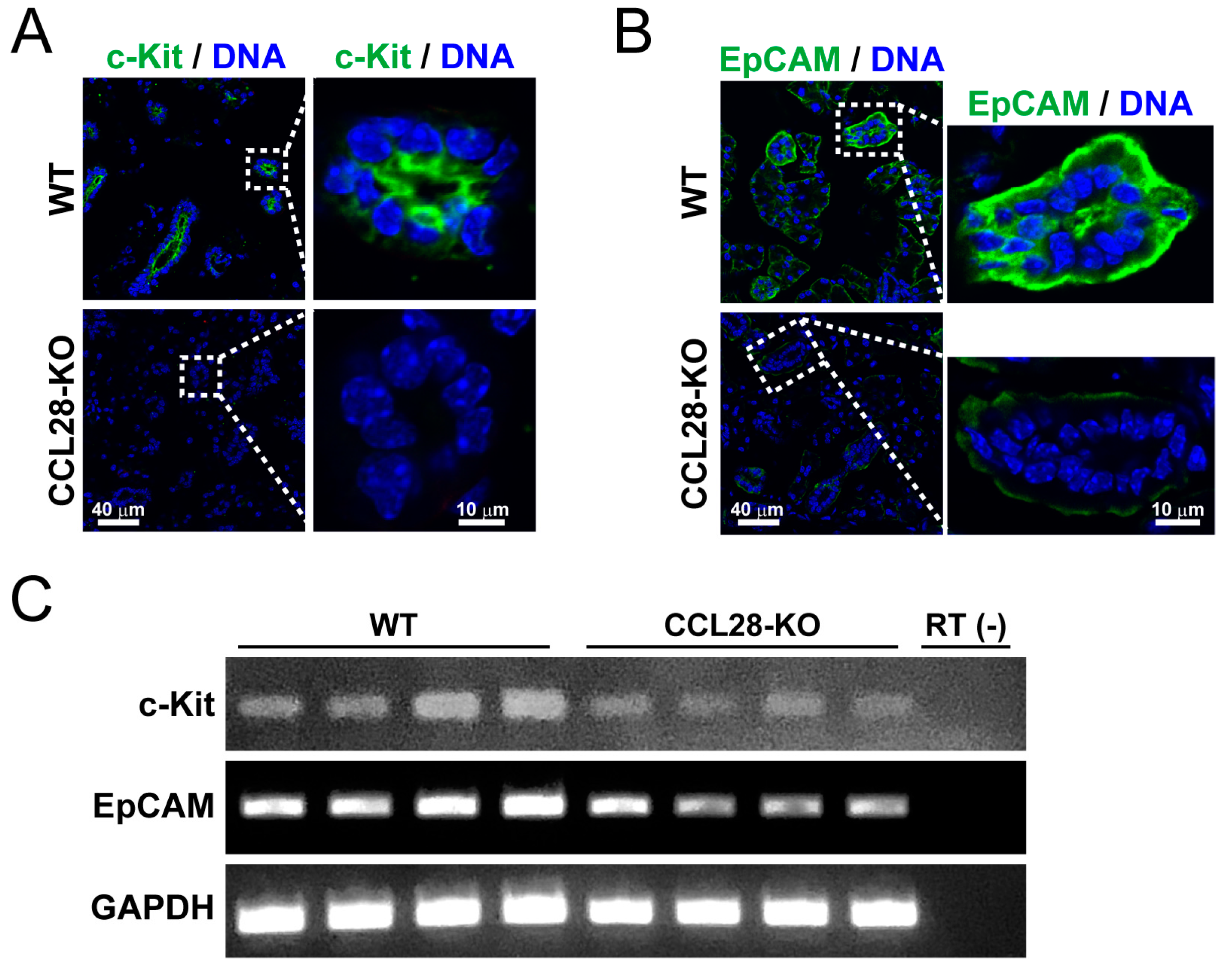
3.4. CCL28 Regulates the Differentiation of the Stem Cells in the Salivary Gland
3.5. Angiogenesis for the Organization of the Salivary Gland Will Require CCL28
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Şenel, S. An Overview of Physical, Microbiological and Immune Barriers of Oral Mucosa. Int. J. Mol. Sci. 2021, 22, 7821. [Google Scholar] [CrossRef]
- Marusiak, M.J.; Paulden, M.; Ohinmaa, A. Professional Oral Health Care Prevents Mouth-Lung Infection in Long-Term Care Homes: A Systematic Review. Can. J. Dent. Hyg. CJDH = J. Can. l’hygiene Dent. JCHD 2023, 57, 180–190. [Google Scholar]
- Yilmaz, Ö.; Lee, K.L. The Inflammasome and Danger Molecule Signaling: At the Crossroads of Inflammation and Pathogen Persistence in the Oral Cavity. Periodontology 2000 2015, 69, 83–95. [Google Scholar] [CrossRef]
- Peng, X.; Cheng, L.; You, Y.; Tang, C.; Ren, B.; Li, Y.; Xu, X.; Zhou, X. Oral Microbiota in Human Systematic Diseases. Int. J. Oral Sci. 2022, 14, 14. [Google Scholar] [CrossRef]
- Rusthen, S.; Kristoffersen, A.K.; Young, A.; Galtung, H.K.; Petrovski, B.É.; Palm, Ø.; Enersen, M.; Jensen, J.L. Dysbiotic Salivary Microbiota in Dry Mouth and Primary Sjögren’s Syndrome Patients. PLoS ONE 2019, 14, e0218319. [Google Scholar] [CrossRef] [PubMed]
- El Kholy, K.; Genco, R.J.; Van Dyke, T.E. Oral Infections and Cardiovascular Disease. Trends Endocrinol. Metab. 2015, 26, 315–321. [Google Scholar] [CrossRef]
- Casarin, R.C.V.; Barbagallo, A.; Meulman, T.; Santos, V.R.; Sallum, E.A.; Nociti, F.H.; Duarte, P.M.; Casati, M.Z.; Gonçalves, R.B. Subgingival Biodiversity in Subjects with Uncontrolled Type-2 Diabetes and Chronic Periodontitis. J. Periodontal Res. 2013, 48, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Nagakubo, D.; Kaibori, Y. Oral Microbiota: The Influences and Interactions of Saliva, IgA, and Dietary Factors in Health and Disease. Microorganisms 2023, 11, 2307. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, A.; Sørensen, C.E.; Proctor, G.B.; Carpenter, G.H. Salivary Functions in Mastication, Taste and Textural Perception, Swallowing and Initial Digestion. Oral Dis. 2018, 24, 1399–1416. [Google Scholar] [CrossRef] [PubMed]
- van der Bilt, A.; Engelen, L.; Pereira, L.J.; van der Glas, H.W.; Abbink, J.H. Oral Physiology and Mastication. Physiol. Behav. 2006, 89, 22–27. [Google Scholar] [CrossRef] [PubMed]
- van’t Hof, W.; Veerman, E.C.I.; Nieuw Amerongen, A.V.; Ligtenberg, A.J.M. Antimicrobial Defense Systems in Saliva. In Monographs in Oral Science; S. Karger AG: Basel, Switzerland, 2014; Volume 24, pp. 40–51. [Google Scholar]
- Fábián, T.K.; Hermann, P.; Beck, A.; Fejérdy, P.; Fábián, G. Salivary Defense Proteins: Their Network and Role in Innate and Acquired Oral Immunity. Int. J. Mol. Sci. 2012, 13, 4295–4320. [Google Scholar] [CrossRef]
- Walker, D.M. Oral Mucosal Immunology: An Overview. Ann. Acad. Med. Singap. 2004, 33, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.F.; Shen, Y.Y.; Zhang, M.C.; Lv, M.C.; Wang, T.Y.; Chen, X.Q.; Lin, J. Progress in Salivary Glands: Endocrine Glands with Immune Functions. Front. Endocrinol. 2023, 14, 1061235. [Google Scholar] [CrossRef]
- Proctor, G.B.; Carpenter, G.H. Regulation of Salivary Gland Function by Autonomic Nerves. Auton. Neurosci. 2007, 133, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Selye, H.; Veilleux, R.; Cantin, M. Excessive Stimulation of Salivary Gland Growth by Isoproterenol. Science 1961, 133, 44–45. [Google Scholar] [CrossRef] [PubMed]
- Hand, A.R.; Ho, B. Mitosis and Hypertrophy of Intercalated Duct Cells and Endothelial Cells in the Isoproterenol-Treated Rat Parotid Gland. J. Dent. Res. 1985, 64, 1031–1038. [Google Scholar] [CrossRef]
- Boshell, J.; Pennington, C. Histological Observations on the Effects of Isoproterenol on Regenerating Submandibular Glands of the Rat. Cell Tissue Res. 1980, 213, 411–416. [Google Scholar] [CrossRef]
- Emmerson, E.; Knox, S.M. Salivary Gland Stem Cells: A Review of Development, Regeneration and Cancer. Genesis 2018, 56, e23211. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Susa, T.; Shimizu, K.; Sawai, N.; Suzuki, T.; Aoki, T.; Yokoo, S.; Takata, K. Function of the Membrane Water Channel Aquaporin-5 in the Salivary Gland. ACTA Histochem. Cytochem. 2012, 45, 251–259. [Google Scholar] [CrossRef]
- Ma, T.; Song, Y.; Gillespie, A.; Carlson, E.J.; Epstein, C.J.; Verkman, A.S. Defective Secretion of Saliva in Transgenic Mice Lacking Aquaporin-5 Water Channels. J. Biol. Chem. 1999, 274, 20071–20074. [Google Scholar] [CrossRef]
- Hosoi, K.; Yao, C.; Hasegawa, T.; Yoshimura, H.; Akamatsu, T. Dynamics of Salivary Gland AQP5 under Normal and Pathologic Conditions. Int. J. Mol. Sci. 2020, 21, 1182. [Google Scholar] [CrossRef] [PubMed]
- Hieshima, K.; Kawasaki, Y.; Hanamoto, H.; Nakayama, T.; Nagakubo, D.; Kanamaru, A.; Yoshie, O. CC Chemokine Ligands 25 and 28 Play Essential Roles in Intestinal Extravasation of IgA Antibody-Secreting Cells. J. Immunol. 2004, 173, 3668–3675. [Google Scholar] [CrossRef] [PubMed]
- Hieshima, K.; Ohtani, H.; Shibano, M.; Izawa, D.; Nakayama, T.; Kawasaki, Y.; Shiba, F.; Shiota, M.; Katou, F.; Saito, T.; et al. CCL28 Has Dual Roles in Mucosal Immunity as a Chemokine with Broad-Spectrum Antimicrobial Activity. J. Immunol. 2003, 170, 1452–1461. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Nagakubo, D.; Yamamoto, S.; Shigeta, A.; Tomida, S.; Fujita, M.; Hirata, T.; Tsunoda, I.; Nakayama, T.; Yoshie, O. CCL28-Deficient Mice Have Reduced IgA Antibody–Secreting Cells and an Altered Microbiota in the Colon. J. Immunol. 2018, 200, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Nagakubo, D.; Yoshie, O.; Hirata, T. Upregulated CCL28 Expression in the Nasal Mucosa in Experimental Allergic Rhinitis: Implication for CD4+ Memory T Cell Recruitment. Cell. Immunol. 2016, 302, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Kaibori, Y.; Yamashita, K.; Nagakubo, D. The Altered Production and Property of Saliva Induced by Ingesting Fermented Food Ingredients Affect the Oral Microbiome Composition in Mice. Biosci. Biotechnol. Biochem. 2023, 87, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Nagakubo, D.; Swann, J.B.; Birmelin, S.; Boehm, T. Autoimmunity Associated with Chemically Induced Thymic Dysplasia. Int. Immunol. 2017, 29, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Kaibori, Y.; Nagakubo, D. CCR3 Blockage Elicits Polyploidization Associated with the Signatures of Epithelial-Mesenchymal Transition in Carcinoma Cell Lines. Cancer Gene Ther. 2023, 30, 137–148. [Google Scholar] [CrossRef]
- Das, B.; Cash, M.N.; Hand, A.R.; Shivazad, A.; Grieshaber, S.S.; Robinson, B.; Culp, D.J. Tissue Distribution of Murine Muc19/Smgc Gene Products. J. Histochem. Cytochem. 2010, 58, 141–156. [Google Scholar] [CrossRef]
- Bu, Y.; Liu, Y.; Zhang, T.; Liu, Y.; Zhang, Z.; Yi, H. Bacteriocin-Producing Lactiplantibacillus Plantarum YRL45 Enhances Intestinal Immunity and Regulates Gut Microbiota in Mice. Nutrients 2023, 15, 3437. [Google Scholar] [CrossRef]
- Culp, D.J.; Robinson, B.; Cash, M.N.; Bhattacharyya, I.; Stewart, C.; Cuadra-Saenz, G. Salivary Mucin 19 Glycoproteins. J. Biol. Chem. 2015, 290, 2993–3008. [Google Scholar] [CrossRef]
- Peyrot des Gachons, C.; Breslin, P.A.S. Salivary Amylase: Digestion and Metabolic Syndrome. Curr. Diab. Rep. 2016, 16, 102. [Google Scholar] [CrossRef]
- Martens, S.; Coolens, K.; Olsen, C.; Lefesvre, P.; Rooman, I. Heterotopia of Salivary Gland Tissue in the Pancreas. Diagn. Pathol. 2023, 18, 98. [Google Scholar] [CrossRef]
- Khan, S.; Fitch, S.; Knox, S.; Arora, R. Exocrine Gland Structure-Function Relationships. Development 2022, 149, dev197657. [Google Scholar] [CrossRef]
- Odenwald, M.A.; Choi, W.; Buckley, A.; Shashikanth, N.; Joseph, N.E.; Wang, Y.; Warren, M.H.; Buschmann, M.M.; Pavlyuk, R.; Hildebrand, J.; et al. ZO-1 Interactions with F-Actin and Occludin Direct Epithelial Polarization and Single Lumen Specification in 3D Culture. J. Cell Sci. 2016, 130, 243–259. [Google Scholar] [CrossRef]
- Adhikari, N.; Neupane, S.; Roh, J.; Jun, J.H.; Jung, J.-K.; Sohn, W.-J.; Kim, J.-Y.; Kim, J.-Y. Immunolocalization Patterns of Cytokeratins during Salivary Acinar Cell Development in Mice. J. Mol. Histol. 2018, 49, 1–15. [Google Scholar] [CrossRef]
- Furuse, C.; de Sousa, S.O.M.; Nunes, F.D.; de Magalhaes, M.H.C.G.; de Arauijo, V.C. Myoepithelial Cell Markers in Salivary Gland Neoplasms. Int. J. Surg. Pathol. 2005, 13, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Lombaert, I.M.A.; Brunsting, J.F.; Wierenga, P.K.; Faber, H.; Stokman, M.A.; Kok, T.; Visser, W.H.; Kampinga, H.H.; de Haan, G.; Coppes, R.P. Rescue of Salivary Gland Function after Stem Cell Transplantation in Irradiated Glands. PLoS ONE 2008, 3, e2063. [Google Scholar] [CrossRef] [PubMed]
- Nanduri, L.S.Y.; Maimets, M.; Pringle, S.A.; Van Der Zwaag, M.; Van Os, R.P.; Coppes, R.P. Regeneration of Irradiated Salivary Glands with Stem Cell Marker Expressing Cells. Radiother. Oncol. 2011, 99, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Banh, A.; Xiao, N.; Cao, H.; Chen, C.-H.; Kuo, P.; Krakow, T.; Bavan, B.; Khong, B.; Yao, M.; Ha, C.; et al. A Novel Aldehyde Dehydrogenase-3 Activator Leads to Adult Salivary Stem Cell Enrichment In Vivo. Clin. Cancer Res. 2011, 17, 7265–7272. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; van der Zwaag, M.; Stokman, M.A.; van Os, R.; Coppes, R.P. Isolation and Characterization of Human Salivary Gland Cells for Stem Cell Transplantation to Reduce Radiation-Induced Hyposalivation. Radiother. Oncol. 2009, 92, 466–471. [Google Scholar] [CrossRef]
- Maimets, M.; Rocchi, C.; Bron, R.; Pringle, S.; Kuipers, J.; Giepmans, B.N.G.; Vries, R.G.J.; Clevers, H.; de Haan, G.; van Os, R.; et al. Long-Term In Vitro Expansion of Salivary Gland Stem Cells Driven by Wnt Signals. Stem Cell Rep. 2016, 6, 150–162. [Google Scholar] [CrossRef]
- Huang, G.; Tao, L.; Shen, S.; Chen, L. Hypoxia Induced CCL28 Promotes Angiogenesis in Lung Adenocarcinoma by Targeting CCR3 on Endothelial Cells. Sci. Rep. 2016, 6, 27152. [Google Scholar] [CrossRef]
- Facciabene, A.; Peng, X.; Hagemann, I.S.; Balint, K.; Barchetti, A.; Wang, L.-P.; Gimotty, P.A.; Gilks, C.B.; Lal, P.; Zhang, L.; et al. Tumour Hypoxia Promotes Tolerance and Angiogenesis via CCL28 and Treg Cells. Nature 2011, 475, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Lertkiatmongkol, P.; Liao, D.; Mei, H.; Hu, Y.; Newman, P.J. Endothelial Functions of Platelet/Endothelial Cell Adhesion Molecule-1 (CD31). Curr. Opin. Hematol. 2016, 23, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Blokland, S.L.M.; Flessa, C.-M.; van Roon, J.A.G.; Mavragani, C.P. Emerging Roles for Chemokines and Cytokines as Orchestrators of Immunopathology in Sjögren’s Syndrome. Rheumatology 2021, 60, 3072–3087. [Google Scholar] [CrossRef]
- Hernandez-Molina, G.; Burkhardt, A.M.; Lima, G.; Zlotnik, A.; Betanzos, J.L.; Bahena, S.; Llorente, L. Absence of Salivary CCL28 in Primary Sjögren’s Syndrome. Rheumatol. Int. 2015, 35, 1431–1434. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhu, F.; Yu, X.; Wang, J.; Wu, B.; Li, C. Serum CCL28 as a Biomarker for Diagnosis and Evaluation of Sjögren’s Syndrome. Scand. J. Rheumatol. 2023, 52, 200–207. [Google Scholar] [CrossRef]
- Slomiany, B.L.; Murty, V.L.N.; Piotrowski, J.; Slomiany, A. Salivary Mucins in Oral Mucosal Defense. Gen. Pharmacol. Vasc. Syst. 1996, 27, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Urkasemsin, G.; Ferreira, J.N. Unveiling Stem Cell Heterogeneity Toward the Development of Salivary Gland Regenerative Strategies. In Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2019; Volume 1123, pp. 151–164. [Google Scholar]
- Pringle, S.; Van Os, R.; Coppes, R.P. Concise Review: Adult Salivary Gland Stem Cells and a Potential Therapy for Xerostomia. Stem Cells 2013, 31, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, C.; Emmerson, E. Mouth-Watering Results: Clinical Need, Current Approaches, and Future Directions for Salivary Gland Regeneration. Trends Mol. Med. 2020, 26, 649–669. [Google Scholar] [CrossRef] [PubMed]
- Hai, B.; Yang, Z.; Shangguan, L.; Zhao, Y.; Boyer, A.; Liu, F. Concurrent Transient Activation of Wnt/β-Catenin Pathway Prevents Radiation Damage to Salivary Glands. Int. J. Radiat. Oncol. 2012, 83, e109–e116. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.; Lee, S.; Choi, N.; Shin, H.-S.; Kim, J.; Lim, J.-Y. Single Cell Clones Purified from Human Parotid Glands Display Features of Multipotent Epitheliomesenchymal Stem Cells. Sci. Rep. 2016, 6, 36303. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, P.; Carrillo, E.; Vélez, C.; Hita-Contreras, F.; Martínez-Amat, A.; Rodríguez-Serrano, F.; Boulaiz, H.; Ortiz, R.; Melguizo, C.; Prados, J.; et al. Regulatory Systems in Bone Marrow for Hematopoietic Stem/Progenitor Cells Mobilization and Homing. Biomed Res. Int. 2013, 2013, 312656. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, L.; Pock, T.; Tröndle, I.; Neuhaus, N. The C-X-C Signalling System in the Rodent vs Primate Testis: Impact on Germ Cell Niche Interaction. Reproduction 2018, 155, R211–R219. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hale, J.S.; Rich, J.N.; Ransohoff, R.M.; Lathia, J.D. Chemokine CXCL12 in Neurodegenerative Diseases: An SOS Signal for Stem Cell-Based Repair. Trends Neurosci. 2012, 35, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Nakamura, S.; Suzuki, R.; Islam, N.; Domon, T.; Yamamoto, T.; Wakita, M. Apoptosis and Mitosis of Parenchymal Cells in the Duct-Ligated Rat Submandibular Gland. Tissue Cell 2000, 32, 457–463. [Google Scholar] [CrossRef]
- Purwanti, N.; Karabasil, M.R.; Matsuo, S.; Chen, G.; Javkhlan, P.; Azlina, A.; Hasegawa, T.; Yao, C.; Akamatsu, T.; Hosoi, K. Induction of Sca-1 via Activation of STAT3 System in the Duct Cells of the Mouse Submandibular Gland by Ligation of the Main Excretory Duct. Am. J. Physiol. Liver Physiol. 2011, 301, G814–G824. [Google Scholar] [CrossRef]
- Hellhammer, D.H.; Wüst, S.; Kudielka, B.M. Salivary Cortisol as a Biomarker in Stress Research. Psychoneuroendocrinology 2009, 34, 163–171. [Google Scholar] [CrossRef]
- Nater, U.M.; La Marca, R.; Florin, L.; Moses, A.; Langhans, W.; Koller, M.M.; Ehlert, U. Stress-Induced Changes in Human Salivary Alpha-Amylase Activity—Associations with Adrenergic Activity. Psychoneuroendocrinology 2006, 31, 49–58. [Google Scholar] [CrossRef]
- Song, M.; Bai, H.; Zhang, P.; Zhou, X.; Ying, B. Promising Applications of Human-Derived Saliva Biomarker Testing in Clinical Diagnostics. Int. J. Oral Sci. 2023, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Vedam, V.K.V.; Boaz, K.; Natarajan, S.; Ganapathy, S. Salivary Amylase as a Marker of Salivary Gland Function in Patients Undergoing Radiotherapy for Oral Cancer. J. Clin. Lab. Anal. 2017, 31, e22048. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaibori, Y.; Tamoto, S.; Okuda, S.; Matsuo, K.; Nakayama, T.; Nagakubo, D. CCL28: A Promising Biomarker for Assessing Salivary Gland Functionality and Maintaining Healthy Oral Environments. Biology 2024, 13, 147. https://doi.org/10.3390/biology13030147
Kaibori Y, Tamoto S, Okuda S, Matsuo K, Nakayama T, Nagakubo D. CCL28: A Promising Biomarker for Assessing Salivary Gland Functionality and Maintaining Healthy Oral Environments. Biology. 2024; 13(3):147. https://doi.org/10.3390/biology13030147
Chicago/Turabian StyleKaibori, Yuichiro, Saho Tamoto, Sayoko Okuda, Kazuhiko Matsuo, Takashi Nakayama, and Daisuke Nagakubo. 2024. "CCL28: A Promising Biomarker for Assessing Salivary Gland Functionality and Maintaining Healthy Oral Environments" Biology 13, no. 3: 147. https://doi.org/10.3390/biology13030147
APA StyleKaibori, Y., Tamoto, S., Okuda, S., Matsuo, K., Nakayama, T., & Nagakubo, D. (2024). CCL28: A Promising Biomarker for Assessing Salivary Gland Functionality and Maintaining Healthy Oral Environments. Biology, 13(3), 147. https://doi.org/10.3390/biology13030147