

Identification and Characterisation of the CircRNAs Involved in the Regulation of Leaf Colour in Quercus mongolica
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Measurement of Leaf Colour Parameters
2.3. Determination of the Physiological Indicators
2.4. RNA Extraction, Library Construction and Sequencing
2.5. Identification of CircRNA
2.6. Analysis of Differentially Expressed CircRNAs
2.7. Real-Time PCR Analysis
2.8. Enrichment Analysis of Parental DEC Genes
2.9. Statistical Analysis
3. Results
3.1. Changes in the Leaf Colour Parameters
3.2. Changes in Leaf Pigment Content
3.3. Hormonal Changes
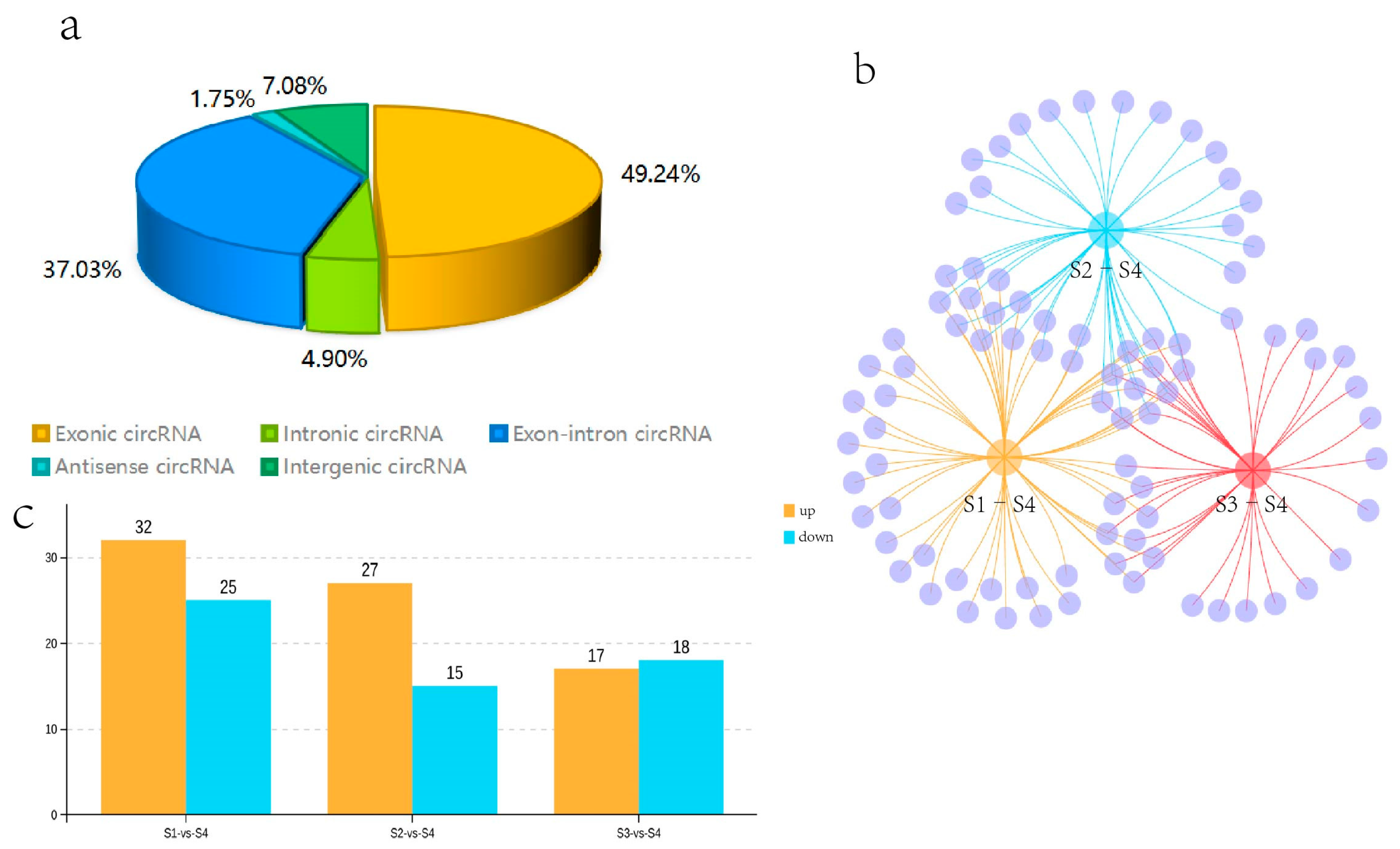
3.4. CircRNA Identification
3.5. Identification of Differentially Expressed CircRNAs
3.6. Enrichment Analysis of the DEC Parental Genes
3.7. Correlation Analysis
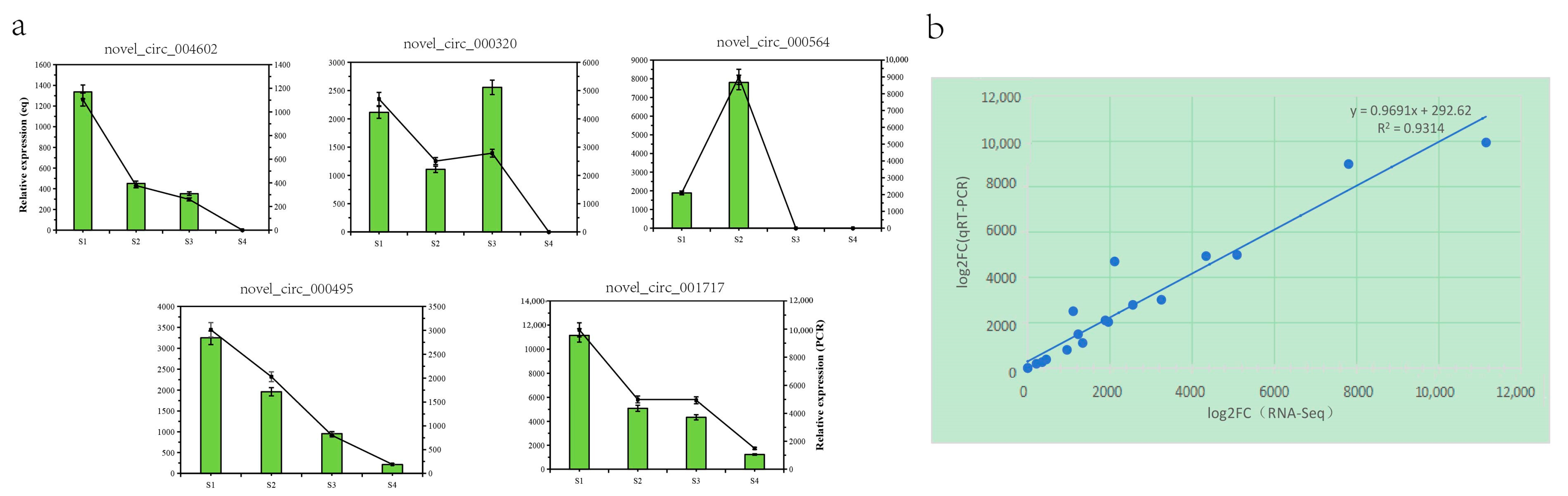
3.8. Real-Time PCR Verification of the DECs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zeng, Y.; Wang, W.; Liao, W.; Wang, H.; Zhang, D. Multiple glacial refugia for cool-temperate deciduous trees in northern East Asia: The Mongolian oak as a case study. Mol. Ecol. 2015, 24, 5676–5691. [Google Scholar] [CrossRef]
- Li, Z. Study on Drought Resistance and Ornamental Value of Betula Platyphylla and Quercus mongolica at Hohhot. Ph.D. Thesis, Inner MongoliaAgricultural University, Hohhot, China, 2015. [Google Scholar]
- Szabo, L.; Salzman, J. Detecting circular RNAs: Bioinformatic and experimental challenges. Nat. Rev. Genet. 2016, 17, 679–692. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, H.; Zhang, Y.; Lu, X.; Chen, L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef]
- Wang, D.; Gao, Y.; Sun, S.; Li, L.; Wang, K. Expression characteristics in roots, phloem, leaves, flowers and fruits of apple circRNA. Genes 2022, 13, 712. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Yang, X.; Li, X.; Wang, J.; Gao, Y.; Shang, R.; Sun, W.; Dou, K.; Li, H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015, 365, 141–148. [Google Scholar] [CrossRef]
- Pan, T.; Sun, X.; Liu, Y.; Li, H.; Deng, G.; Lin, H.; Wang, S. Heat stress alters genome-wide profiles of circular RNAs in Arabidopsis. Plant Mol. Biol. 2018, 96, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Chen, G.; Shi, T. Identifying and characterizing the circular RNAs during the lifespan of Arabidopsis leaves. Front. Plant Sci. 2017, 8, 1278. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Chen, L.; Liu, C.; Zhu, Q.; Fan, L. Widespread noncoding circular RNAs in plants. New Phytol. 2015, 208, 88–95. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, P.; Chen, Q.; Wang, J.; Chen, M. Identification and characteri-zation of ncRNA-associated ceRNA networks in Arabidopsis leaf develop-ment. BMC Genom. 2018, 19, 607. [Google Scholar] [CrossRef]
- Zhao, W.; Cheng, Y.; Zhang, C.; You, Q.; Shen, X.; Guo, W.; Jiao, Y. Genome-wide identification and characterization of circular RNAs by high throughput sequencing in soybean. Sci. Rep. 2017, 7, 5636. [Google Scholar] [CrossRef]
- Chen, L.; Ding, X.; Zhang, H.; He, T.; Li, Y.; Wang, T.; Li, X.; Jin, L.; Song, Q.; Yang, S.; et al. Comparative analysis of circular RNAs between soybean cytoplas-mic male-sterile line NJCMS1A and its maintainer NJCMS1B by high-throughput sequencing. BMC Genom. 2018, 19, 663. [Google Scholar] [CrossRef]
- Lu, T.; Cui, L.; Zhou, Y.; Zhu, C.; Fan, D.; Gong, H.; Zhao, Q.; Zhou, C.; Zhao, Z.; Lu, D.; et al. Transcriptome-wide investigation of circular RNAs in rice. RNA 2015, 21, 2076–2087. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, P.; Fan, Y.; Lu, Q.; Li, Q.; Yan, J.; Muehlbauer, G.; Schnable, P.; Dai, M.; Li, L. Circular RNAs mediated by transposons are associated with transcriptomic and phenotypic variation in maize. New Phytol. 2017, 217, 1292–1306. [Google Scholar] [CrossRef]
- Ghorbani, A.; Izadpanah, K.; Peters, J.; Dietzgen, R.; Mitter, N. Detection and profiling of circular RNAs in uninfected and maize Iranian mosaic virus-infected maize. Plant Sci. 2018, 274, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Hao, Z.; Zhu, Y.; Zhang, H.; Li, G. Genome-wide identification and functional analysis of circRNAs in Zea mays. PLoS ONE 2018, 13, e0202375. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Wang, Q.; Zhu, B.; Luo, Y.; Gao, L. Deciphering the roles of circRNAs on chilling injury in tomato. Biochem. Biophys. Res. Commun. 2016, 479, 132–138. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Y.; Jin, L.; Ling, X.; Liu, T.; Chen, T.; Ji, Y.; Yu, W.; Zhang, B. Re-analysis of long non-coding RNAs and prediction of circRNAs reveal their novel roles in susceptible tomato following TYLCV infection. BMC Plant Biol. 2018, 18, 104. [Google Scholar] [CrossRef] [PubMed]
- Darbani, B.; Noeparvar, S.; Borg, S. Identification of circular RNAs from the parental genes involved in multiple aspects of cellular metabolism in barley. Front. Plant Sci. 2016, 7, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.; Yu, J.; Hou, Y.; Li, F.; Zhou, Q.; Wei, C.; Bennetzen, J. Circular RNA archi-tecture and differentiation during leaf bud to young leaf development in tea (Camellia sinensis). Planta 2018, 248, 1417–1429. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Wang, L.; Li, S.; Xu, M.; Guan, X.; Zhou, B. Characterization of conserved circular RNA in polyploid Gossypium species and their ancestors. FEBS Lett. 2017, 591, 3660–3669. [Google Scholar] [CrossRef]
- Xiang, L.; Cai, C.; Cheng, J.; Wang, L.; Wu, C.; Shi, Y.; Luo, J.; He, L.; Deng, Y.; Zhang, X.; et al. Identification of circularRNAs and their targets in Gossypium under Verticillium wilt stress based on RNA-seq. PeerJ 2018, 6, e4500. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, M.; Wei, S.; Qin, F.; Zhao, H.; Suo, B. Identification of circular RNAs and their targets in leaves of Triticum aestivum L. under dehydration stress. Front. Plant Sci. 2017, 7, 2024. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Yue, H.; Li, L.; Xu, Y.; Wang, Z.; Xin, Z.; Lin, T. Identification and characteri-zation of circRNAs involved in the regulation of low nitrogen-promoted-root growth in hexaploid wheat. Biol. Res. 2018, 51, 43. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ren, Y.; Lin, T.; Cui, D. Identifcation and characterization of CircRNAs involved in the regulation of wheat root length. Biol. Res. 2019, 52, 19. [Google Scholar] [CrossRef] [PubMed]
- Chappell, J.; Hahlbrock, K. Transcription of plant defence genes in response to UV light or fungal elicitor. Nature 1984, 311, 76–78. [Google Scholar] [CrossRef]
- Shi, M.; Xie, D. Biosynthesis and metabolic engineering of anthocyanins in Arabidopsis thaliana. Recent Pat. Biotechnol. 2014, 8, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ding, W.; Li, Y.; Liu, J.; Wang, L.; Yue, Y. Cloning and expression analysis of anthocyanidin synthase (LlANS) in Lycoris longituba. J. Henan Agric. Univ. 2018, 52, 611–617. [Google Scholar]
- Wang, Y.; Wang, C.C.; Zhou, X.; Bi, W.; Yang, Y.M.; Wang, J. Cloning and functional analysis of a flavonoid-3-O-glucosyltransferas gene from Indosasa hispida. Guihaia 2015, 35, 244–249. [Google Scholar]
- Zhang, S.L. Preliminary Study on Leaf Structure and Physiological and Biochemical Characteristics of Photinia frasery during Leaf Color Change. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2017. [Google Scholar]
- Yang, X.; Yang, N.; Zhang, Q.; Pei, Z.; Chang, M.; Zhou, H.; Ge, Y.; Yang, Q.; Li, G. Anthocyanin biosynthesis associated with natural variation in autumn leaf coloration in Quercus aliena accessions. Int. J. Mol. Sci. 2022, 23, 12179. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, W.; Pang, J.; Zhou, M.; Liu, J.; Zhao, J.; Sui, J.; Huang, D.; Yang, M. Integrated physiological and metabolomic analyses reveal changes during the natural senescence of Quercus mongolica leaves. PLoS ONE 2023, 18, e0289272. [Google Scholar] [CrossRef]
- Wang, Y. Research on Mechanism of Leaf Coloration in Autumn of Fraxinus angustifolia. Ph.D. Thesis, Hebei Agricultural University, Baoding, China, 2021. [Google Scholar]
- Wang, X.; Huang, J. Principles and Techniques of Plant Physiological Biochemical Experiment, 2nd ed.; Beijing Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Li, H. Principles and Techniques of Plant Physiological Biochemical Experiment, 3rd ed.; Beijing Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Zhang, J.; Chen, S.; Yang, J.; Zhao, F. Accurate quantification of circular RNAs identifies extensive circular isoform switching events. Nat. Commun. 2020, 11, 90. [Google Scholar] [CrossRef]
- Bao, G.L.; Zhao, F.F.; Wang, J.Q.; Liu, X.; Hu, J.; Shi, B.G.; Wen, Y.L.; Zhao, L.; Luo, Y.Z.; Li, S.B. Characterization of the circRNA-miRNA-mRNA network to reveal the potential functional ceRNAs associated with dynamic changes in the meat quality of the longissimus thoracis muscle in Tibetan Sheep at different growth stages. Front. Vet. Sci. 2022, 9, 803758. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.; McCarthy, D.; Smyth, G. edgeR: A Bioconductor package for differential expression analysis of digital gene expressio data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Jeyaraj, A.; Zhang, X.; Hou, Y.; Shangguan, M.; Gajjeraman, P.; Li, Y.; Wei, C. Genome-wide identifcation of conserved and novel microRNAs in one bud and two tender leaves of tea plant (Camellia sinensis) by small RNA sequencing, microarray-based hybridization and genome survey scafold sequences. BMC Plant Biol. 2017, 17, 212. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Tian, C.; Jiang, Q.; Li, X.; Zhuang, J. Selection of suitable reference genes for qRT-PCR normalization during leaf development and hormonal stimuli in tea plant (Camellia sinensis). Sci. Rep. 2016, 6, 19748. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Q.M.; Ding, Y.F.; Yu, H.; Zhu, G.Z. Physiological characters of Euonymus europaea leaves during the color-changing period in autumn and winter. Acta Bot. Boreali-Occident. Sin. 2018, 38, 1072–1079. [Google Scholar]
- Guo, H.H.; Liu, Y.; Yao, F.; Li, S.A.; Song, X.H. Relationship between pigment contents and leaf color parameters of Pistacia chinensis bunge of different provenances in autumn. Acta Bot. Boreali-Occident. Sin. 2017, 37, 2003–2009. [Google Scholar]
- Meyer, H.J.; Staden, J. The in vitro production of an anthocyanin from callus cultures of Oxalis linearis. Plant Cell Tissue Organ Cult. 1995, 40, 55–58. [Google Scholar] [CrossRef]
- Zhou, L.L.; Zeng, H.N.; Shi, M.Z.; Xie, D.Y. Development of tobacco callus cultures over expressing Arabidopsis PAP1/MYB75 transcription factor and characterization of anthocyanin biosynthesis. Planta 2008, 229, 37–51. [Google Scholar] [CrossRef]
- Ji, X.H.; Wang, Y.T.; Zhang, R.; Wu, S.J.; An, M.M.; Li, M.; Wang, C.Z.; Chen, X.L.; Zhang, Y.M.; Chen, X.S. Effect of auxin, cytokinin and nitrogen on anthocyanin biosynthesis in callus cultures of redfleshed apple (Malus sieversii f. niedzwetzkyana). Plant Cell Tissue Organ Cult. 2015, 120, 325–337. [Google Scholar] [CrossRef]
- Das, P.K.; Shin, D.H.; Choi, S.B.; Yoo, S.D.; Choi, G.; Park, Y.I. Cytokinins enhance sugarinduced anthocyanin biosynthesis in Arabidopsis. Mol. Cells 2012, 34, 93–101. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, B.H.; Kim, S.H.; Oh, K.H.; Yun Cho, K. Response to environmental and chemical signals for anthocyanin biosynthesis in nonchlorophyllous corn (Zea mays L.) leaf. J. Plant Biol. 2006, 49, 16–25. [Google Scholar] [CrossRef]
- Wang, H.C.; Huang, X.M.; Hu, G.B.; Huang, H.B. Studies on the relationship between anthocyanin biosynthesis andrelated enzymes in litchi pericarp. Sci. Agric. Sin. 2004, 37, 2028–2032. [Google Scholar]
- Wei, Y.Z.; Hu, F.C.; Hu, G.B.; Li, X.J.; Huang, X.M.; Wang, H.C. Differential expression of anthocyanin biosynthetic genes in relation to anthocyanin accumulation in the pericarp of Litchi chinensis Sonn. PLoS ONE 2011, 6, e19455. [Google Scholar] [CrossRef] [PubMed]
- Lai, B.; Li, X.J.; Hu, B.; Qin, Y.H.; Huang, X.M.; Wang, H.C.; Hu, G.B. LcMYB1 is a key determinant of differential anthocyanin accumulation among genotypes, tissues, developmental phases and ABA and light stimuli in Litchi chinensis. PLoS ONE 2014, 9, e86293. [Google Scholar] [CrossRef]
- Hosokawa, K. Cell layer-specific accumulation of anthocyanins in response to gibberellic acid in tepals of Hyacinthus orientalis. Biosci. Biotechnol. Biochem. 1999, 63, 930–931. [Google Scholar] [CrossRef]
- Weiss, D.; Van der Luit, A.; Knegt, E.; Vermeer, E.; Mol, J.N.M.; Kooter, J.M. Identification of endogenous gibberellins in petunia flower, induction of anthocyanin biosynthetic gene expression and the antagonistic effect of abscisic acid. Plant Physiol. 1995, 107, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Ilan, A.; Dougall, D.K. The effect of growth retardantson anthocyanin production in carrot cell suspension cultures. Plant Cell Rep. 1992, 11, 304–309. [Google Scholar] [PubMed]
- Jeong, S.W.; Das, P.K.; Jeoung, S.C.; Song, J.Y.; Lee, H.K.; Kim, Y.K.; Kim, W.J.; Park, Y.I.; Yoo, S.D.; Choi, S.B.; et al. Ethylene suppression of sugar-induced anthocyanin pigmentation in Arabidopsis. Plant Physiol. 2010, 154, 1515–1531. [Google Scholar] [CrossRef] [PubMed]
- El-Kereamy, A.; Chervin, C.; Roustan, J.P.; Cheyhnier, V.; Souquet, J.M.; Moutounet, M.; Raynal, J.; Ford, C.; Latché, A.; Pech, J.C.; et al. Exogenous ethylene stimulates the long term expression of genes related to anthocyanin biosynthesis in grape berries. Physiol. Plant. 2003, 119, 175–282. [Google Scholar] [CrossRef]
- Gagné, S.; Cluzet, S.; Mérillon, J.M.; Gény, L. ABA initiates anthocyanin production in grape cell cultures. J. Plant Growth Regul. 2011, 30, 1–10. [Google Scholar] [CrossRef]
- Qu, S.S.; Li, M.M.; Wang, G.; Zhu, S.J. Application of ABA and GA3 alleviated browning of litchi (Litchi chinensis Sonn.) via different strategies. Postharvest Biol. Technol. 2021, 181, 111672. [Google Scholar] [CrossRef]
- Ni, J.B.; Zhao, Y.; Tao, R.Y.; Yin, L.; Gao, L.; Strid, A.; Qian, M.J.; Li, J.C.; Li, Y.J.; Shen, J.Q.; et al. Ethylene mediates the branching of the jasmonate-induced flavonoid biosynthesis pathway by suppressing anthocyanin biosynthesis in red Chinese pear fruits. Plant Biotechnol. J. 2020, 18, 1223–1240. [Google Scholar] [CrossRef] [PubMed]
- Lasda, E.; Parker, R. Circular RNAs: Diversity of form and function. RNA 2014, 20, 1829–1842. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef]
- Litholdo, C.G.; Da, F.G.C. Circular RNAs and plant stress responses. Adv. Exp. Med. Biol. 2018, 1087, 345–353. [Google Scholar]
- Huang, X.; Zhang, H.; Guo, R.; Wang, Q.; Liu, X.; Kuang, W.; Song, H.; Liao, J.; Huang, Y.; Wang, Z. Systematic identification and characterization of circular RNAs involved in flag leaf senescence of rice. Planta 2021, 253, 26. [Google Scholar] [CrossRef]
- Liu, G.; Gu, H.; Cai, H.; Guo, C.; Chen, Y.; Wang, L.; Chen, G. Integrated transcriptome and biochemical analysis provides new insights into the leaf color change in Acer fabri. Forests 2023, 14, 1638. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Zheng, Q.; Bao, C.; Guo, W.; Li, S.; Chen, J.; Chen, B.; Luo, Y.; Lyu, D.; Li, Y.; Shi, G.; et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat. Commun. 2016, 7, 11215. [Google Scholar] [CrossRef]
- Kulcheski, F.R.; Christoff, A.P.; Margis, R. Circular RNAs are miRNA sponges and can be used as a new class of biomarker. J. Biotechnol. 2016, 238, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Zhou, J.; Hu, C.; Zhang, J. Transcriptome-wide identification and functional prediction of novel and flowering-related circular RNAs from trifoliate orange (Poncirus trifoliata L. Raf.). Planta 2018, 247, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Capelari, R.F.; Fonseca, G.C.D.; Guzman, F.; Margis, R. Circular and micro RNAs from Arabidopsis thaliana flowers are simultaneously isolated from AGO-IP libraries. Plants 2019, 8, 302. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Y.; Pang, X.; Pang, J.; Wang, Q.; Zhou, M.; Lu, Y.; Xu, C.; Huang, D. Identification and Characterisation of the CircRNAs Involved in the Regulation of Leaf Colour in Quercus mongolica. Biology 2024, 13, 183. https://doi.org/10.3390/biology13030183
Yuan Y, Pang X, Pang J, Wang Q, Zhou M, Lu Y, Xu C, Huang D. Identification and Characterisation of the CircRNAs Involved in the Regulation of Leaf Colour in Quercus mongolica. Biology. 2024; 13(3):183. https://doi.org/10.3390/biology13030183
Chicago/Turabian StyleYuan, Yangchen, Xinbo Pang, Jiushuai Pang, Qian Wang, Miaomiao Zhou, Yan Lu, Chenyang Xu, and Dazhuang Huang. 2024. "Identification and Characterisation of the CircRNAs Involved in the Regulation of Leaf Colour in Quercus mongolica" Biology 13, no. 3: 183. https://doi.org/10.3390/biology13030183
APA StyleYuan, Y., Pang, X., Pang, J., Wang, Q., Zhou, M., Lu, Y., Xu, C., & Huang, D. (2024). Identification and Characterisation of the CircRNAs Involved in the Regulation of Leaf Colour in Quercus mongolica. Biology, 13(3), 183. https://doi.org/10.3390/biology13030183