Estrogen and Progesterone Receptors Are Dysregulated at the BPH/5 Mouse Preeclamptic-Like Maternal–Fetal Interface

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Husbandry
2.2. Quantitative (q) RT-PCR
2.3. Immunohistochemistry
2.4. Statistical Analysis
3. Results
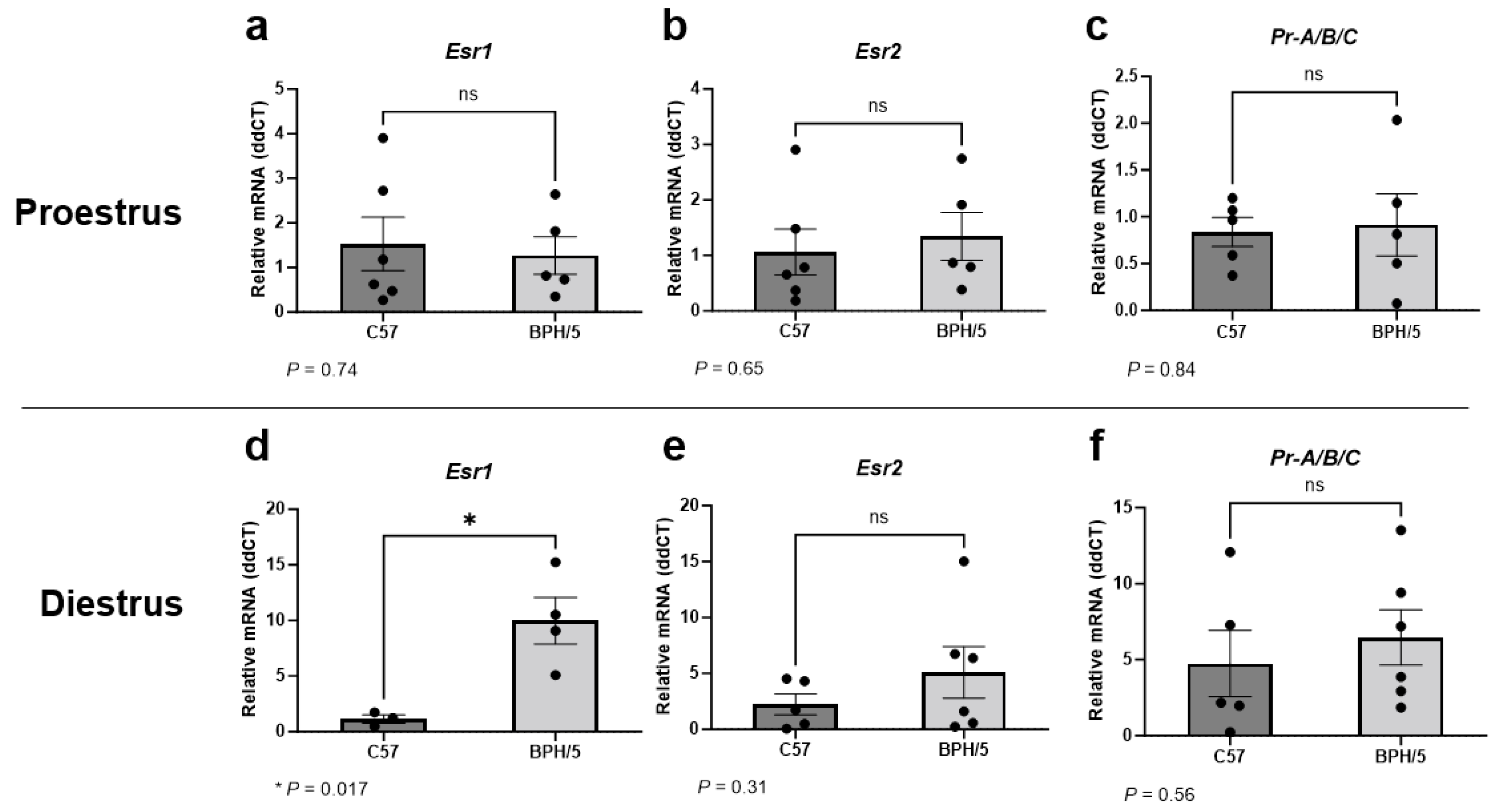
3.1. Uterine Esr1 Is Upregulated in Virgin BPH/5 Females during Diestrus
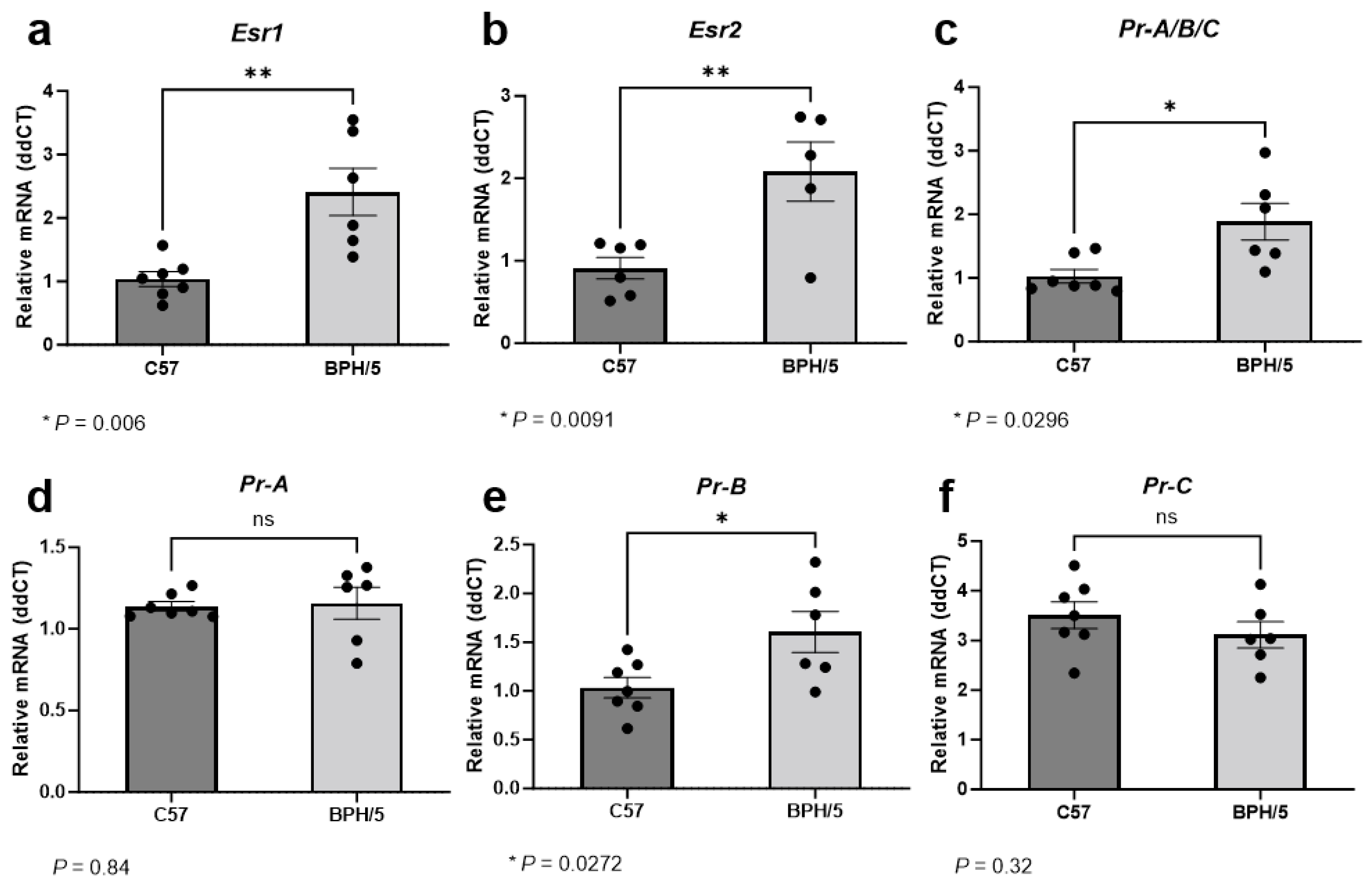
3.2. At the Peak of Decidualization, Esr1, Esr2, and Pr Isoform B Are Upregulated in the Preeclamptic-Like BPH/5 Maternal–Fetal Interface
3.3. A Higher Population of Decidual Cells Display Nuclear Erα Expression in Early BPH/5 Pregnancies
3.4. At the Peak of Decidualization, Pr Is Downregulated in the BPH/5 Decidua, Luminal, and Glandular Uterine Epithelium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Say, L.; Chou, D.; Gemmill, A.; Tunçalp, Ö.; Moller, A.-B.; Daniels, J.; Gülmezoglu, A.M.; Temmerman, M.; Alkema, L. Global causes of maternal death: A WHO systematic analysis. Lancet Glob. Health 2014, 2, e323–e333. [Google Scholar] [CrossRef]
- Wallis, A.B.; Saftlas, A.F.; Hsia, J.; Atrash, H.K. Secular trends in the rates of preeclampsia, eclampsia, and gestational hypertension, United States, 1987–2004. Am. J. Hypertens. 2008, 21, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Annath, C.V.; Keyes, K.M.; Wapner, R.J. Pre-eclampsia rates in the United States, 1980–2010: Age-period-cohort analysis. BMJ 2013, 347, f6564. [Google Scholar] [CrossRef] [PubMed]
- Wojtowicz, A.; Zembala-Szczerba, M.; Babczyk, D.; Kolodziejczyk-Pietruszka, M.; Lewaczynska, O.; Huras, H. Early- and Late-Onset Preeclampsia: A Comprehensive Cohort Study of Laboratory and Clinical Findings according to the New ISHHP Criteria. Int. J. Hypertens. 2019, 2019, 4108271. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Redman, C.W.; Roberts, J.M.; Moffett, A. Pre-eclampsia: Pathophysiology and clinical implications. BMJ 2019, 366, l2381. [Google Scholar] [CrossRef] [PubMed]
- ACOG. ACOG Practice Bulletin No. 222: Clinical, management guidelines, for obstetrician-gynecologists. Obstet. Gynecol. 2020, 135, e237–e260. [Google Scholar]
- Roberts, J.M.; Rich-Edwards, J.W.; McElrath, T.F.; Garmire, L.; Myatt, L.; Global Pregnancy, C. Subtypes of Preeclampsia: Recognition and Determining Clinical Usefulness. Hypertension 2021, 77, 1430–1441. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.G.; Staff, A.C.; Roberts, J.M. Syncytiotrophoblast stress in preeclampsia: The convergence point for multiple pathways. Am. J. Obstet. Gynecol. 2022, 226, S907–S927. [Google Scholar] [CrossRef]
- Than, N.G.; Romero, R.; Tarca, A.L.; Kekesi, K.A.; Xu, Y.; Xu, Z.; Juhasz, K.; Bhatti, G.; Leavitt, R.J.; Gelencser, Z.; et al. Integrated Systems Biology Approach Identifies Novel Maternal and Placental Pathways of Preeclampsia. Front. Immunol. 2018, 9, 1661. [Google Scholar] [CrossRef]
- Raymond, D.; Peterson, E. A Critical Review of Early-Onset and Late-Onset Preeclampsia. Obstet. Gynecol. Surv. 2011, 66, 497–506. [Google Scholar] [CrossRef]
- Lisonkova, S.; Joseph, K.S. Incidence of preeclampsia: Risk factors and outcomes associated with early- versus late-onset disease. Am. J. Obstet. Gynecol. 2013, 209, 544.e1–544.e12. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.J.; Seow, K.M.; Chen, K.H. Preeclampsia: Recent Advances in Predicting, Preventing, and Managing the Maternal and Fetal Life-Threatening Condition. Int. J. Environ. Res. Public Health 2023, 20, 2994. [Google Scholar] [CrossRef]
- Rybak-Krzyszkowska, M.; Staniczek, J.; Kondracka, A.; Boguslawska, J.; Kwiatkowski, S.; Gora, T.; Strus, M.; Gorczewski, W. From Biomarkers to the Molecular Mechanism of Preeclampsia-A Comprehensive Literature Review. Int. J. Mol. Sci. 2023, 24, 13252. [Google Scholar] [CrossRef] [PubMed]
- Stairs, J.; Hsieh, T.Y.J.; Rolnik, D.L. In Vitro Fertilization and Adverse Pregnancy Outcomes in the Elective Single Embryo Transfer Era. Am. J. Perinatol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Ganer Herman, H.; Volodarsky-Perel, A.; Ton Nu, T.N.; Machado-Gedeon, A.; Cui, Y.; Shaul, J.; Dahan, M.H. Diminished ovarian reserve is a risk factor for preeclampsia and placental malperfusion lesions. Fertil. Steril. 2023, 119, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Reschke, L.; Segars, J.; Baker, V.L. Frozen-thawed embryo transfer: The potential importance of the corpus luteum in preventing obstetrical complications. Fertil. Steril. 2020, 113, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Naicker, T.; Khedun, S.M.; Moddley, J.; Pijnenborg, R. Quantitative analysis of trophoblast invasion in preeclampsia. Acta Obstet. Gynecol. Scand. 2003, 82, 722–729. [Google Scholar] [CrossRef]
- Gomes, V.C.L.; Woods, A.K.; Crissman, K.R.; Landry, C.A.; Beckers, K.F.; Gilbert, B.M.; Ferro, L.F.; Liu, C.-C.; Oberhaus, E.L.; Sones, J.L. Kisspeptin is upregulated at the maternal-fetal interface of the preeclamptic-like BPH/5 mouse and normalized after synchronization of sex steroid hormones. Reprod. Med. 2022, 3, 263–279. [Google Scholar] [CrossRef]
- O’Brien, J.E.; Peterson, T.J.; Tong, M.H.; Lee, E.J.; Pfaff, L.E.; Hewitt, S.C.; Korach, K.S.; Weiss, J.; Jameson, J.L. Estrogen-induced proliferation of uterine epithelial cells is independent of estrogen receptor alpha binding to classical estrogen response elements. J. Biol. Chem. 2006, 281, 26683–26692. [Google Scholar] [CrossRef]
- Noyola-Martinez, N.; Halhali, A.; Barrera, D. Steroid hormones and pregnancy. Gynecol. Endocrinol. 2019, 35, 376–384. [Google Scholar] [CrossRef]
- Kim, H.R.; Kim, Y.S.; Yoon, J.A.; Yang, S.C.; Park, M.; Seol, D.W.; Lyu, S.W.; Jun, J.H.; Lim, H.J.; Lee, D.R.; et al. Estrogen induces EGR1 to fine-tune its actions on uterine epithelium by controlling PR signaling for successful embryo implantation. FASEB J. 2018, 32, 1184–1195. [Google Scholar] [CrossRef]
- Tan, J.; Paria, B.C.; Dey, S.K.; Das, S.K. Differential uterine expression of estrogen and progesterone receptors correlates with uterine preparation for implantation and decidualization in the mouse. Endocrinology 1999, 140, 5310–5321. [Google Scholar] [CrossRef]
- Massri, N.; Loia, R.; Sones, J.L.; Arora, R.; Douglas, N.C. Vascular changes in the cycling and early pregnant uterus. JCI Insight 2023, 8, e163422. [Google Scholar] [CrossRef]
- Cheng, J.; Sha, Z.; Li, J.; Li, B.; Luo, X.; Zhang, Z.; Zhou, Y.; Chen, S.; Wang, Y. Progress on the Role of Estrogen and Progesterone Signaling in Mouse Embryo Implantation and Decidualization. Reprod. Sci. 2023, 30, 1746–1757. [Google Scholar] [CrossRef]
- Condon, J.C.; Hardy, D.B.; Kovaric, K.; Mendelson, C.R. Up-regulation of the progesterone receptor (PR)-C isoform in laboring myometrium by activation of nuclear factor-κB may contribute to the onset of labor through inhibition of PR function. Mol. Endocrinol. 2006, 20, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Troisi, R.; Potischman, N.; Roberts, J.M.; Ness, R.; Cromblehome, W.; Lykins, D.; Siiteri, P.; Hoover, R.N. Maternal serum oestrogen and androgen concentrations in preeclamptic and uncomplicated pregnancies. Int. J. Epidemiol. 2003, 32, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Jobe, S.O.; Tyler, C.T.; Magness, R.R. Aberrant Synthesis, Metabolism, and Plasma Accumulation of Circulating Estrogens and Estrogen Metabolites in Preeclampsia: Implications for Vascular Dysfunction. Hypertension 2013, 61, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Zhu, X.; Guo, C.; Yang, Y.; Han, T.; Chen, L.; Yin, W.; Gao, P.; Zhang, H.; Geng, J.; et al. Differential expression of estradiol and estrogen receptor alpha in severe preeclamptic pregnancies compared with normal pregnancies. Mol. Med. Rep. 2013, 7, 981–985. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Hu, Z.; Zeng, K.; Yin, Y.; Zhao, M.; Chen, M.; Chen, Q. The reduction in circulating levels of estrogen and progesterone in women with preeclampsia. Pregnancy Hypertens. 2018, 11, 18–25. [Google Scholar] [CrossRef]
- Lan, K.C.; Lai, Y.J.; Cheng, H.H.; Tsai, N.C.; Su, Y.T.; Tsai, C.C.; Hsu, T.Y. Levels of sex steroid hormones and their receptors in women with preeclampsia. Reprod. Biol. Endocrinol. 2020, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.Y.; An, S.M.; Jeong, J.S.; Yang, S.Y.; Lee, G.S.; Hong, E.J.; Jeung, E.B.; Kim, S.C.; An, B.S. Comparison of steroid hormones in three different preeclamptic models. Mol. Med. Rep. 2021, 23, 252. [Google Scholar] [CrossRef]
- Shin, Y.Y.; Jeong, J.S.; Park, M.N.; Lee, J.E.; An, S.M.; Cho, W.S.; Kim, S.C.; An, B.S.; Lee, K.S. Regulation of steroid hormones in the placenta and serum of women with preeclampsia. Mol. Med. Rep. 2018, 17, 2681–2688. [Google Scholar] [CrossRef]
- Shu, C.; Han, S.; Xu, P.; Wang, Y.; Cheng, T.; Hu, C. Estrogen and Preeclampsia: Potential of estrogens as therapeutic agents in preeclampsia. Drug Des. Devel Ther. 2021, 15, 2543–2550. [Google Scholar]
- Iou, S.G.; Eskandari, M.; Dabiri, A. Evaluation of androgen and progesterone levels of women with preeclampsia in third trimester. Med. J. Islam. World Acad. Sci. 2005, 15, 19–22. [Google Scholar]
- Hertig, A.; Liere, P.; Chabbert-Buffet, N.; Fort, J.; Pianos, A.; Eychenne, B.; Cambourg, A.; Schumacher, M.; Berkane, N.; Lefevre, G.; et al. Steroid profiling in preeclamptic women: Evidence for aromatase deficiency. Am. J. Obstet. Gynecol. 2010, 203, 477.e1–477.e9. [Google Scholar] [CrossRef]
- Eissa, S.; Mostafa, M.M.; El-Gendy, A.A.E.A.; Senna, I.A. Quantitative immunological detection of total estrogen receptor (cytosolic and nuclear) in term decidua of preeclampsia: A preliminary study. Clin. Chem. 1997, 43, 405–406. [Google Scholar] [CrossRef]
- Park, M.-N.; Park, K.-H.; Lee, J.-E.; Shin, Y.Y.; An, S.-M.; Kang, S.S.; Cho, W.-S.; An, B.-S.; Kim, S.C. The Expression and Activation of Sex Steroid Receptors in the Preeclamptic Placenta. Int. J. Mol. Med. 2018, 41, 2943–2951. [Google Scholar] [CrossRef]
- Hamilton, K.J.; Hewitt, S.C.; Arao, Y.; Korach, K.S. Estrogen Hormone Biology. Curr. Top. Dev. Biol. 2017, 125, 109–146. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Nakshatri, H.; Nephew, K.P. Inhibitig proteasomal proteolysis sustains estrogen receptor-alpha activation. Mol. Endocrinol. 2004, 18, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef] [PubMed]
- Davisson, R.L.; Hoffman, D.S.; Butz, G.M.; Aldape, G.; Schlager, G.; Merrill, D.C.; Sethi, S.; Weiss, R.M.; Bates, J.N. Discovery of a Spontaneous Genetic Mouse Model of Preeclampsia. Hypertension 2002, 39, 337–342. [Google Scholar]
- Dokras, A.; Hoffmann, D.S.; Eastvold, J.S.; Kienzle, M.F.; Gruman, L.M.; Kirby, P.A.; Weiss, R.M.; Davisson, R.L. Severe feto-placental abnormalities precede the onset of hypertension and proteinuria in a mouse model of preeclampsia. Biol. Reprod. 2006, 75, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Heyward, C.Y.; Sones, J.L.; Lob, H.E.; Yuen, L.C.; Abbott, K.E.; Huang, W.; Begun, Z.R.; Butler, S.D.; August, A.; Leifer, C.A.; et al. The decidua of preeclamptic-like BPH/5 mice exhibits an exaggerated inflammatory response during early pregnancy. J. Reprod. Immunol. 2017, 120, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Sones, J.L.; Cha, J.; Woods, A.K.; Bartos, A.; Heyward, C.Y.; Lob, H.E.; Isroff, C.E.; Butler, S.D.; Shapiro, S.E.; Dey, S.K.; et al. Decidual Cox2 inhibition improves fetal and maternal outcomes in a preeclampsia-like mouse model. JCI Insight 2016, 1, e75351. [Google Scholar] [CrossRef] [PubMed]
- Sutton, E.F.; Lob, H.E.; Song, J.; Xia, Y.; Butler, S.; Liu, C.C.; Redman, L.M.; Sones, J.L. Adverse metabolic phenotype of female offspring exposed to preeclampsia in utero: A characterization of the BPH/5 mouse in postnatal life. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R485–R491. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.N.; Reijnders, D.; Gomes, V.C.L.; Hebert, R.C.; Liu, C.C.; Stephens, J.M.; Redman, L.M.; Douglas, N.C.; Sones, J.L. Complement in Reproductive White Adipose Tissue Characterizes the Obese Preeclamptic-Like BPH/5 Mouse Prior to and During Pregnancy. Biology 2020, 9, 304. [Google Scholar] [CrossRef]
- Woods, A.K.; Hoffmann, D.S.; Weydert, C.J.; Butler, S.D.; Zhou, Y.; Sharma, R.V.; Davisson, R.L. Adenoviral delivery of VEGF121 early in pregnancy prevents spontaneous development of preeclampsia in BPH/5 mice. Hypertension 2011, 57, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Caligioni, C.S. Assessing reproductive status/stages in mice. Curr. Protoc. Neurosci. 2009, 48, A.4I.1–A.4I.8. [Google Scholar] [CrossRef]
- Cora, M.C.; Kooistra, L.; Travlos, G. Vaginal Cytology of the Laboratory Rat and Mouse: Review and Criteria for the Staging of the Estrous Cycle Using Stained Vaginal Smears. Toxicol. Pathol. 2015, 43, 776–793. [Google Scholar] [CrossRef]
- Sones, J.L.; Merriam, A.A.; Seffens, A.; Brown-Grant, D.A.; Butler, S.D.; Zhao, A.M.; Xu, X.; Shawber, C.J.; Grenier, J.K.; Douglas, N.C. Angiogenic factor imbalance precedes complement deposition in placentae of the BPH/5 model of preeclampsia. FASEB J. 2018, 32, 2574–2586. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Goldman, S.; Shalev, E. Difference in progesterone-receptor isoforms ratio between early and late first-trimester human trophoblast is associated with differential cell invasion and matrix metalloproteinase 2 expression. Biol. Reprod. 2006, 74, 13–22. [Google Scholar] [CrossRef]
- Mori, M.; Bogdan, A.; Balassa, T.; Csabai, T.; Szekeres-Bartho, J. The Decidua—The Maternal Bed Embracing the Embryo—Maintains the Pregnancy. Semin. Immunopathol. 2016, 38, 635–649. [Google Scholar] [CrossRef]
- Parisi, F.; Fenizia, C.; Introini, A.; Zavatta, A.; Scaccabarozzi, C.; Biasin, M.; Savasi, V. The pathophysiological role of estrogens in the initial stages of pregnancy: Molecular mechanisms and clinical implications for pregnancy outcome from the periconceptional period to end of the first trimester. Hum. Reprod. Update 2023, 29, 699–720. [Google Scholar] [CrossRef]
- Garrido-Gomez, T.; Dominguez, F.; Quinonero, A.; Diaz-Gimeno, P.; Kapidzic, M.; Gormley, M.; Ona, K.; Padilla-Iserte, P.; McMaster, M.; Genbacev, O.; et al. Defective decidualization during and after severe preeclampsia reveals a possible maternal contribution to the etiology. Proc. Natl. Acad. Sci. USA 2017, 114, E8468–E8477. [Google Scholar] [CrossRef]
- Staff, A.C.; Redman, C.W.G. IFPA Award in Placentology Lecture: Preeclampsia, the decidual battleground and future maternal cardiovascular disease. Placenta 2014, 28, S26–S31. [Google Scholar] [CrossRef] [PubMed]
- Ruane, P.T.; Berneau, S.C.; Koeck, R.; Watts, J.; Kimber, S.J.; Brison, D.R.; Westwood, M.; Aplin, J.D. Apposition to endometrial epithelial cells activates mouse blastocysts for implantation. Mol. Hum. Reprod. 2017, 23, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Sones, J.L.; Yarborough, C.C.; O’Besso, V.; Lemenze, A.; Douglas, N.C. Genotypic analysis of the female BPH/5 mouse, a model of superimposed preeclampsia. PLoS ONE 2021, 16, e0253453. [Google Scholar] [CrossRef] [PubMed]
- Gomes, V.C.L.; Beckers, K.F.; Crissman, K.J.R.; Landry, C.A.; Awad, R.M.; Del Piero, F.; Liu, C.; Sones, J.L. Sexually Dimorphic Pubertal Development and Adipose Tissue Kisspeptin Dysregulation in the Obese and Preeclamptic-like BPH/5 Mouse Model Offspring. Front. Physiol. 2023, 14, 1070426. [Google Scholar] [CrossRef] [PubMed]
- Suba, Z. Compensatory Estrogen Signal Is Capable of DNA Repair in Antiestrogen-Responsive Cancer Cells via Activating Mutations. J. Oncol. 2020, 2020, 5418365. [Google Scholar] [CrossRef] [PubMed]
- Tibbetts, T.A.; Mendoza-Meneses, M.; O’Malley, B.W.; Conneely, O.M. Mutual and Intercompartmental Regulation of Estrogen Receptor and Progesterone Receptor Expression in the Mouse Uterus. Biol. Reprod. 1998, 59, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Bukovsky, A.; Caudle, M.R.; Cekanova, M.; Fernando, R.I.; Wimalasena, J.; Foster, J.S.; Henley, D.C.; Elder, R.F. Placental expression of estrogen receptor beta and its hormonal binding variant—Comparison with estrogen receptor alpha and a role for estrogen receptors in asymetric division and differentiation of estrogen-dependent cells. Reprod. Biol. Endocrinol. 2003, 1, 36. [Google Scholar]
- Feng, X.; Zhou, L.; Mao, X.; Tong, C.; Chen, X.; Zhao, D.; Baker, P.N.; Xia, Y.; Zhang, H. Association of a reduction of Gprotein coupled receptor 30 expression and the pathogenesis of preeclampsia. Mol. Med. Rep. 2017, 16, 5997–6003. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.C.; Fang, L.; Li, Y.; Thakur, A.; Hoodless, P.A.; Guo, Y.; Wang, Z.; Wu, Z.; Yan, Y.; Jia, Q.; et al. G protein-coupled estrogen receptor stimulates human trophoblast cell invasion via YAP-mediated ANGPTL4 expression. Commun. Biol. 2021, 4, 1285. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Feng, X.; Chen, J.; Qi, X.; Zhou, L.; Shi, S.; Kc, K.; Stanley, J.L.; Baker, P.N.; Zhang, H. G protein-coupled receptor 30 regulates trophoblast invasion and its deficiency is associated with preeclampsia. J. Hypertens. 2016, 34, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, N.; Salas-Perez, F.; Ortiz, M.; Alvarez, D.; Echiburu, B.; Maliqueo, M. Rodent models in placental research. Implications for fetal origins of adult disease. Anim. Reprod. 2022, 19, e20210134. [Google Scholar] [CrossRef]
- Arany, Z.; Hilfiker-Kleiner, D.; Karumanchi, S.A. Animal Models of Cardiovascular Complications of Pregnancy. Circ. Res. 2022, 130, 1763–1779. [Google Scholar] [CrossRef]
- Sones, J.L.; Davisson, R.L. Preeclampsia, of mice and women. Physiol. Genom. 2016, 48, 565–572. [Google Scholar] [CrossRef]
- Ain, R.; Canham, L.N.; Soares, M.J. Gestation stage-dependent intrauterine trophoblast cell invasion in the rat and mouse: Novel endocrine phenotype and regulation. Dev. Biol. 2003, 260, 176–190. [Google Scholar] [CrossRef]
- Ramathal, C.Y.; Bagchi, I.C.; Taylor, R.N.; Bagchi, M.K. Endometrial decidualization: Of mice and men. Semin. Reprod. Med. 2010, 28, 17–26. [Google Scholar] [CrossRef]
- Egashira, M.; Hirota, Y. Uterine receptivity and embryo-uterine interactions in embryo implantation: Lessons from mice. Reprod. Med. Biol. 2013, 12, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Geisert, R.D.; Spencer, T.E. (Eds.) Placentation in Mammals: Advances in Anatomy, Embryology and Cell Biology; Springer: Berlin/Heidelberg, Germany, 2021; Volume 234. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, V.C.L.; Gilbert, B.M.; Bernal, C.; Crissman, K.R.; Sones, J.L. Estrogen and Progesterone Receptors Are Dysregulated at the BPH/5 Mouse Preeclamptic-Like Maternal–Fetal Interface. Biology 2024, 13, 192. https://doi.org/10.3390/biology13030192
Gomes VCL, Gilbert BM, Bernal C, Crissman KR, Sones JL. Estrogen and Progesterone Receptors Are Dysregulated at the BPH/5 Mouse Preeclamptic-Like Maternal–Fetal Interface. Biology. 2024; 13(3):192. https://doi.org/10.3390/biology13030192
Chicago/Turabian StyleGomes, Viviane C. L., Bryce M. Gilbert, Carolina Bernal, Kassandra R. Crissman, and Jenny L. Sones. 2024. "Estrogen and Progesterone Receptors Are Dysregulated at the BPH/5 Mouse Preeclamptic-Like Maternal–Fetal Interface" Biology 13, no. 3: 192. https://doi.org/10.3390/biology13030192