
Modulation of Apoptosis by Bovine Gammaherpesvirus 4 Infection in Bovine Endometrial Cells and the Possible Role of LPS in This Process
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of BEC
2.2. BoGHV4 Strain
2.3. Infection of Cells Cultures with BoGHV4 and LPS
2.4. Morphological Analysis by Staining with Rhodamine 123/Propidium Iodide
2.5. Detection of Nuclear Morphological Changes by DAPI Staining
2.6. Detection of DNA Fragmentation by TUNEL Assay
2.7. Total RNA Isolation, cDNA Preparation, and mRNA Expression Analysis by RT-qPCR
2.8. Data Analysis
3. Results
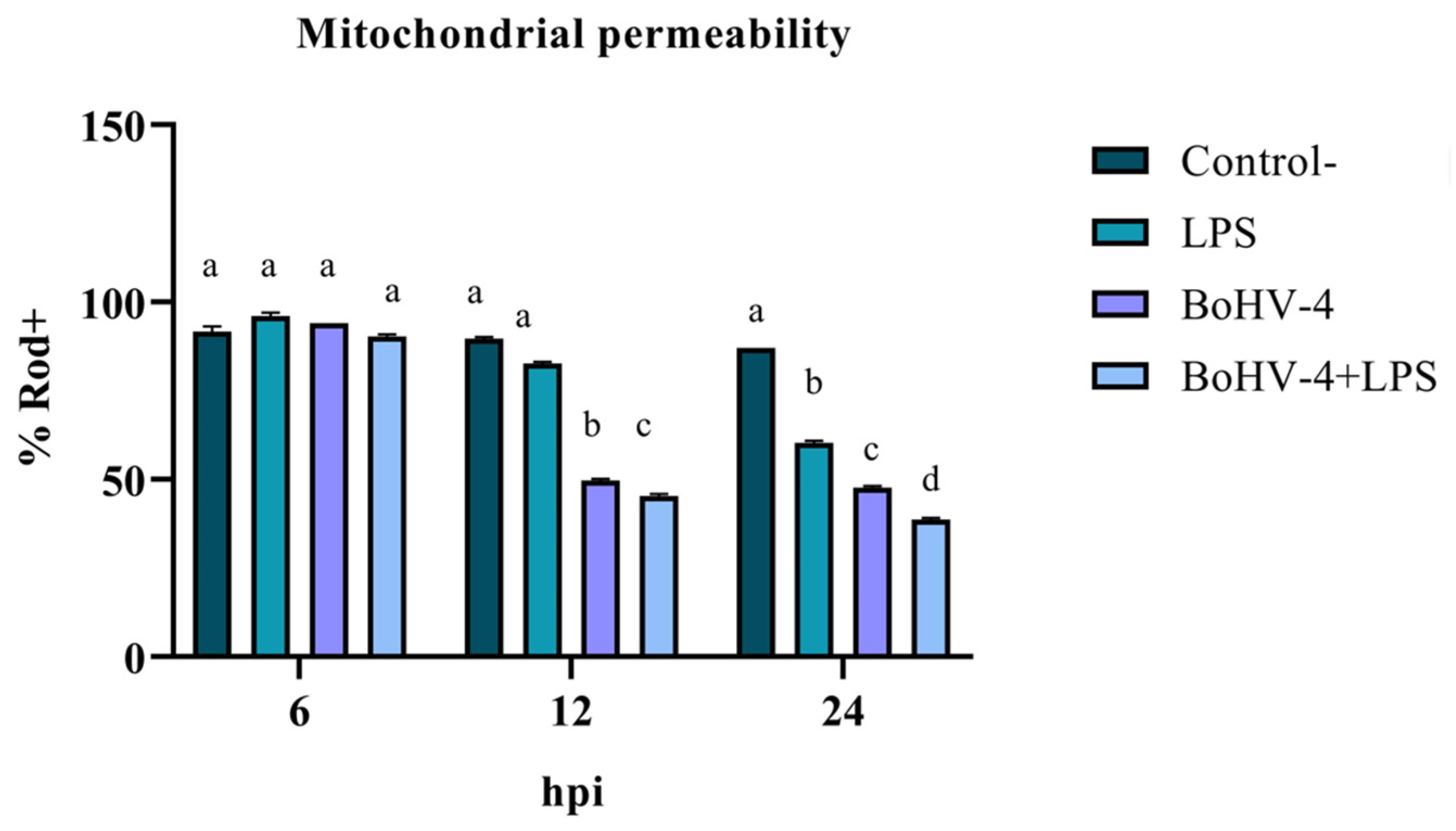
3.1. Evaluation of Early Apoptosis Using Rhodamine 123/Propidium Iodide
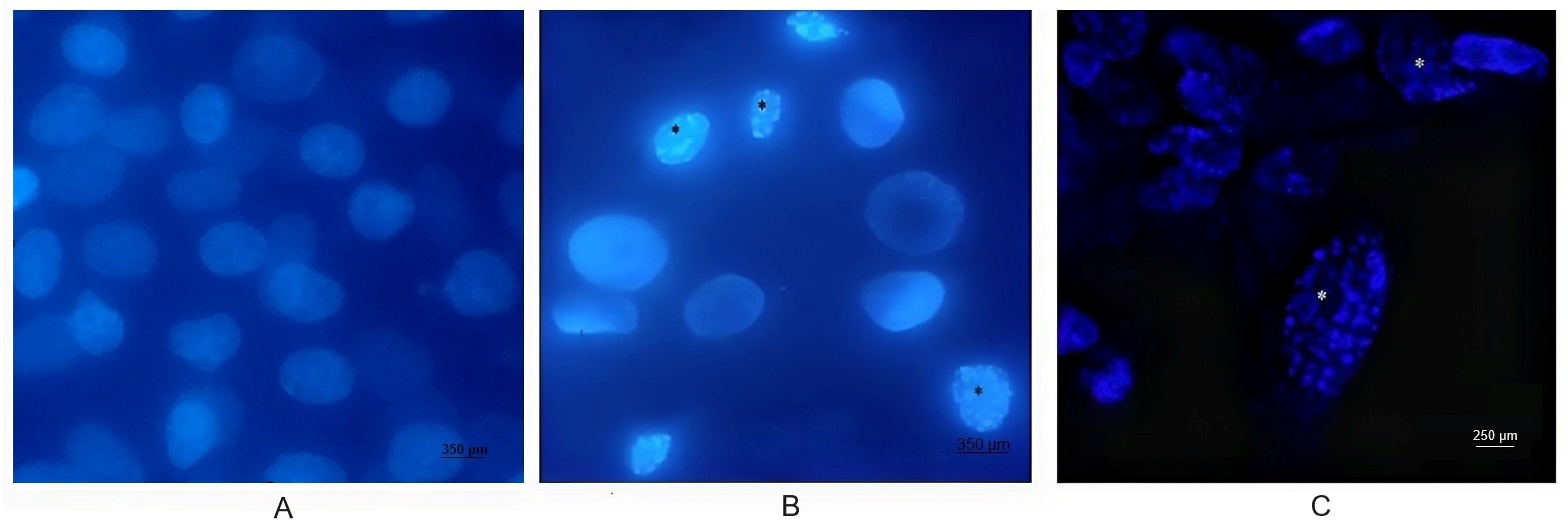
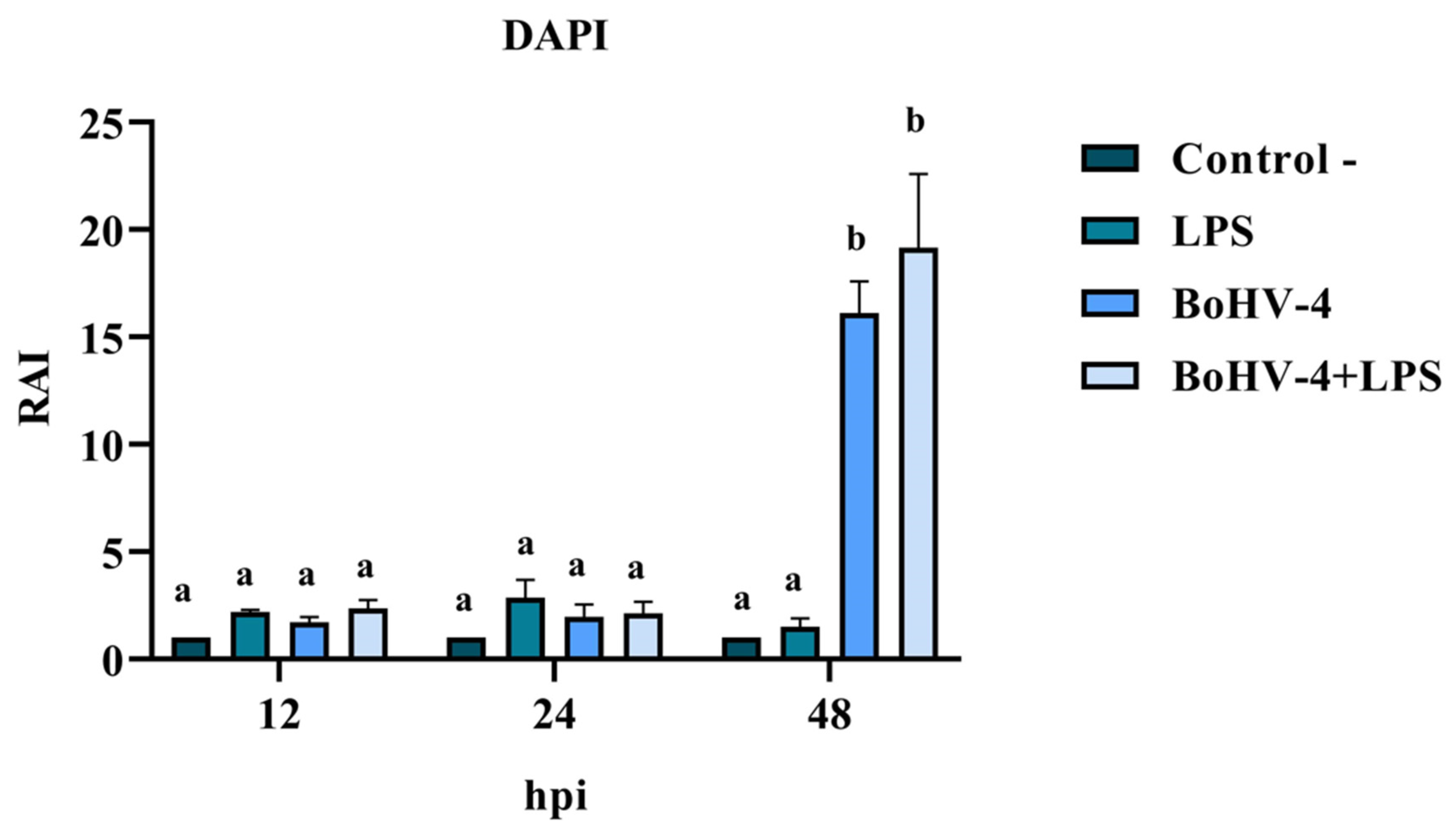
3.2. Nuclear Morphology Changes in BEC Cells
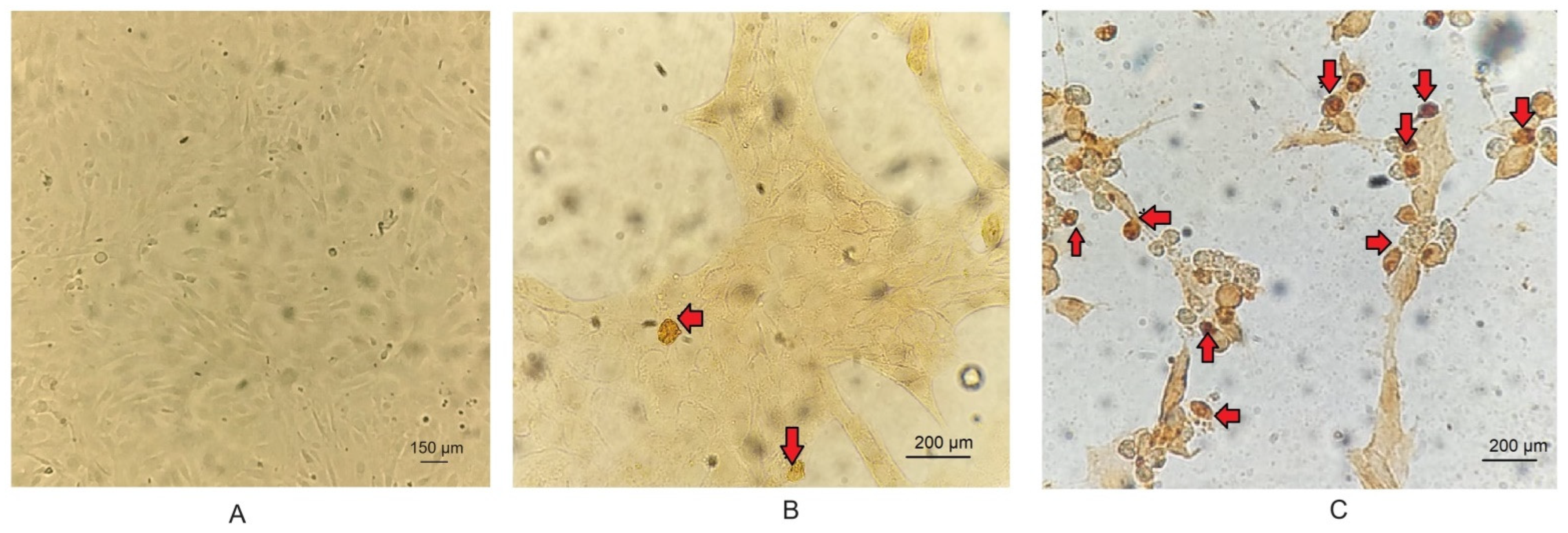
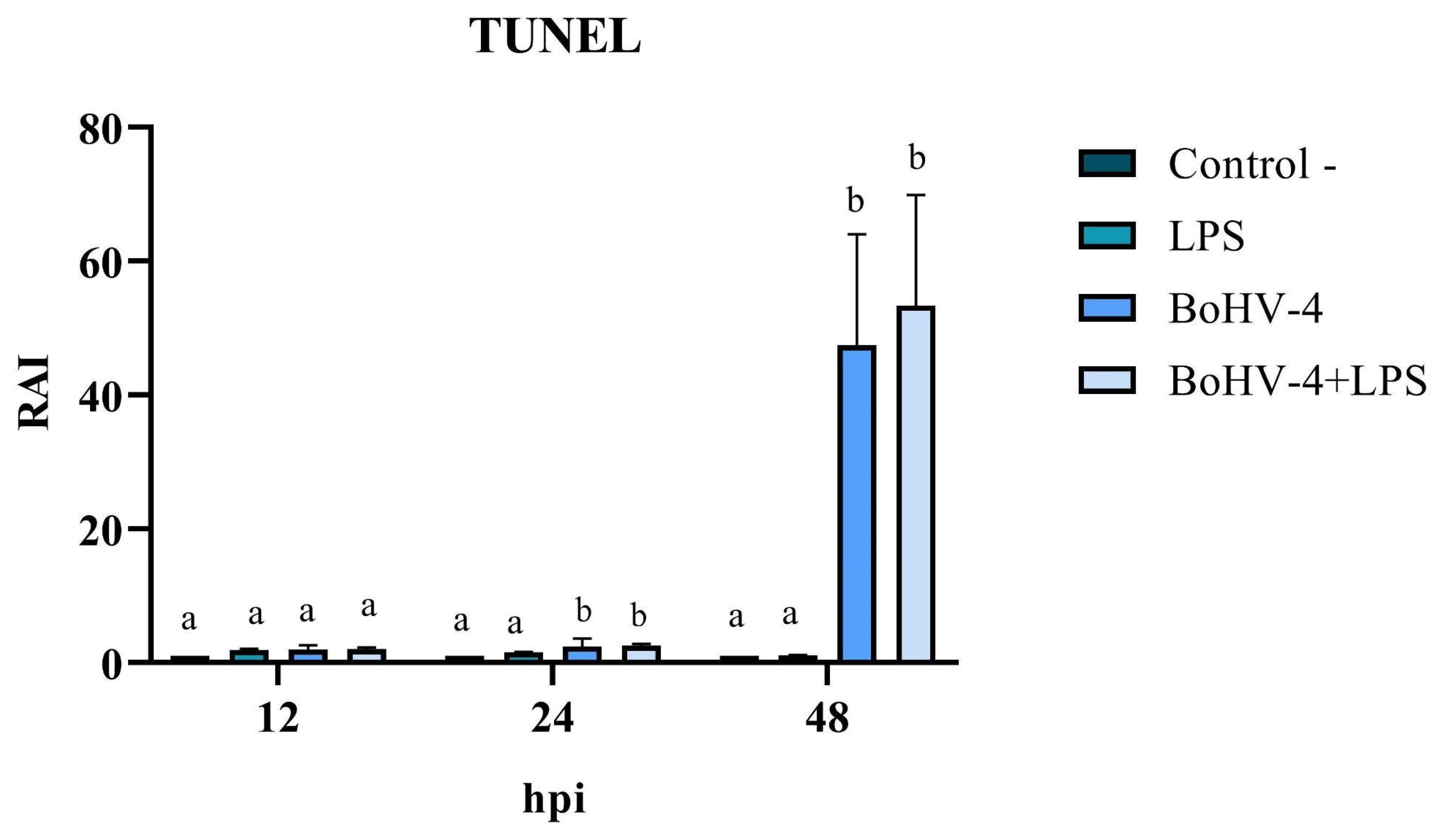
3.3. TUNEL
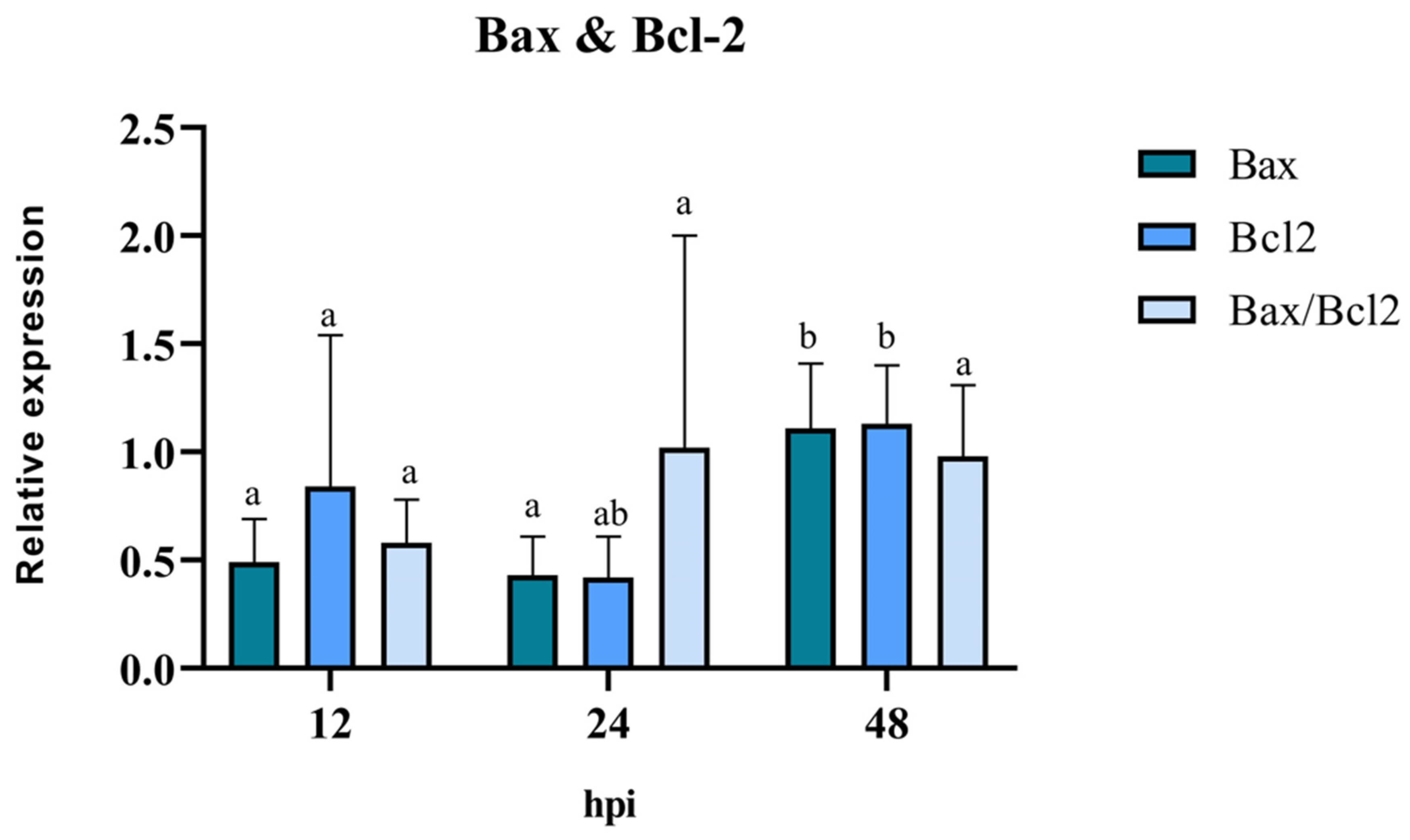
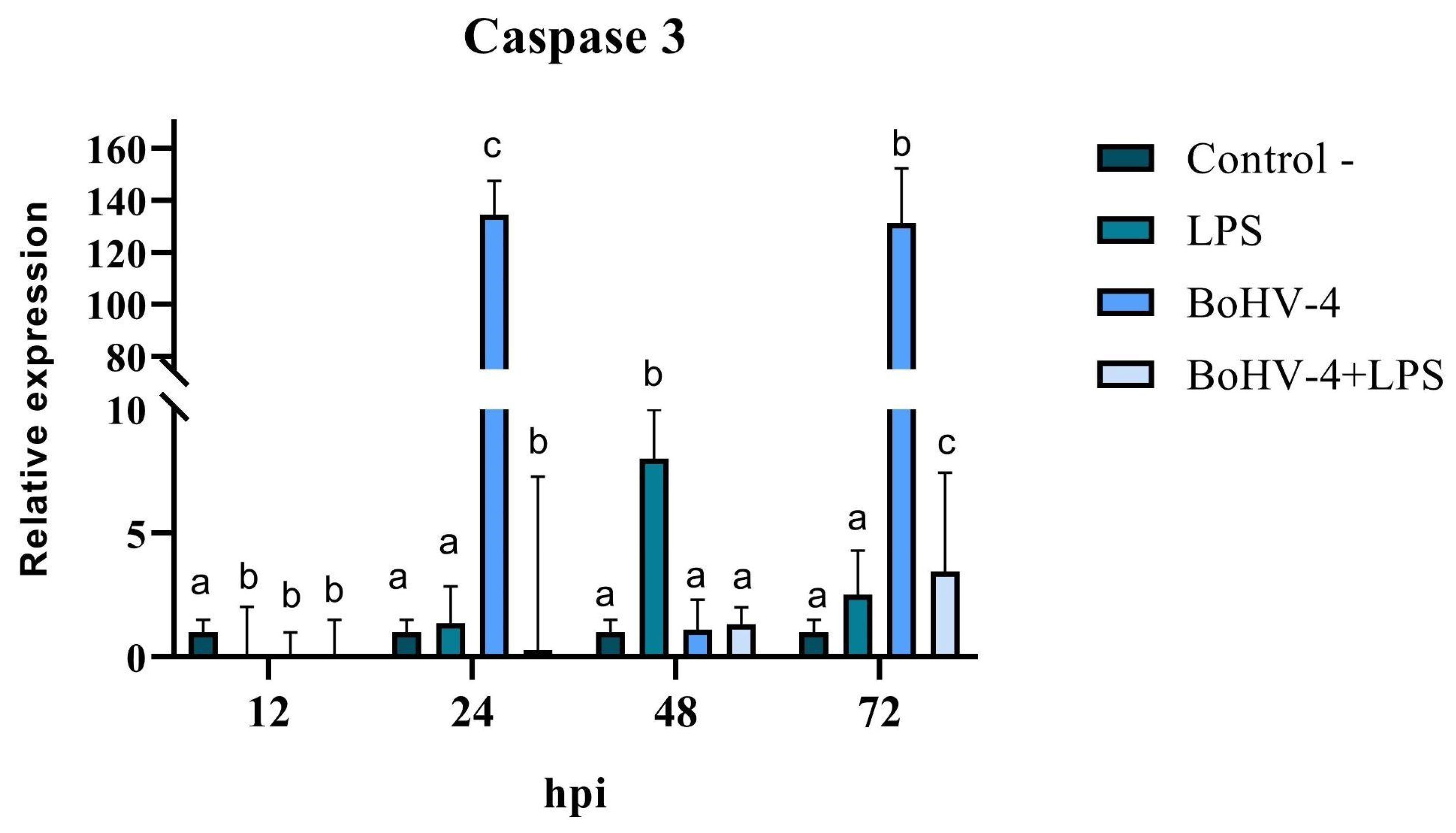
3.4. Pro/Anti-Apoptotic Potential of BoGHV4: Relative Expression of Bax, Bcl-2, and Caspase 3
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Everett, H.; McFadden, G. Apoptosis: An innate immune response to virus infection. Trends Microbiol. 1999, 7, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.G. Viruses, apoptosis, and neuroinflammation—A double-edged sword. J. Neurovirol. 2015, 21, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.A.; Norris, P.S.; Ware, C.F. To kill or be killed: Viral evasion of apoptosis. Nat. Immunol. 2002, 3, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Sarid, R.; Sato, T.; Bohenzky, R.A.; Russo, J.J.; Chang, Y. Kaposi’s sarcoma-associated herpesvirus encodes a functional bcl-2 homologue. Nat. Med. 1997, 3, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, M. PM2.5 induces autophagy and apoptosis through endoplasmic reticulum stress in human endothelial cells. Sci. Total Environ. 2020, 710, 136397. [Google Scholar] [CrossRef] [PubMed]
- Thome, M.; Schneider, P.; Hofmann, K.; Fickenscher, H.; Meinl, E.; Neipel, F.; Mattmann, C.; Burns, K.; Bodmer, J.L.; Schröter, M.; et al. Viral FLICE-inhibitory proteins (FLIPs) prevent apoptosis induced by death receptors. Nature 1997, 386, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Kofod-Olsen, E.; Ross-Hansen, K.; Schleimann, M.H.; Jensen, D.K.; Møller, J.M.; Bundgaard, B.; Mikkelsen, J.G.; Höllsberg, P. U20 is responsible for human herpesvirus 6B inhibition of tumor necrosis factor receptor-dependent signaling and apoptosis. J. Virol. 2012, 86, 11483–11492. [Google Scholar] [CrossRef] [PubMed]
- Morán, P.; Manrique, J.; Pérez, S.; Romeo, F.; Odeón, A.; Jones, L.; Verna, A. Analysis of the anti-apoptotic v-Bcl2 and v-Flip genes and effect on in vitro programmed cell death of Argentinean isolates of bovine gammaherpesvirus 4 (BoGHV4). Microb. Pathog. 2020, 144, 104170. [Google Scholar] [CrossRef]
- Nguyen, M.L.; Blaho, J.A. Apoptosis during herpes simplex virus infection. Adv. Virus Res. 2007, 69, 67–97. [Google Scholar] [CrossRef]
- Chastant-Maillard, S. Impact of Bovine Herpesvirus 4 (BoGHV4) on Reproduction. Transbound. Emerg. Dis. 2015, 62, 245–251. [Google Scholar] [CrossRef]
- Frazier, K.; Pence, M.; Mauel, M.J.; Liggett, A.; Hines, M.E., 2nd; Sangster, L.; Lehmkuhl, H.D.; Miller, D.; Styer, E.; West, J.; et al. Endometritis in postparturient cattle associated with bovine herpesvirus-4 infection: 15 cases. J. Vet. Diagn. Investig. 2001, 13, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Jacca, S.; Franceschi, V.; Colagiorgi, A.; Sheldon, M.; Donofrio, G. Bovine endometrial stromal cells support tumor necrosis factor alpha-induced bovine herpesvirus type 4 enhanced replication. Biol. Reprod. 2013, 88, 135. [Google Scholar] [CrossRef] [PubMed]
- Donofrio, G.; Herath, S.; Sartori, C.; Cavirani, S.; Flammini, C.F.; Sheldon, I.M. Bovine herpesvirus 4 is tropic for bovine endometrial cells and modulates endocrine function. Reproduction 2007, 134, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Donofrio, G.; Ravanetti, L.; Cavirani, S.; Herath, S.; Capocefalo, A.; Sheldon, I.M. Bacterial infection of endometrial stromal cells influences bovine herpesvirus 4 immediate early gene activation: A new insight into bacterial and viral interaction for uterine disease. Reproduction 2008, 136, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Deim, Z.; Szeredi, L.; Egyed, L. Detection of bovine herpesvirus 4 DNA in aborted bovine fetuses. Can. J. Vet. Res. 2007, 71, 226–229. [Google Scholar] [PubMed]
- Sciortino, M.T.; Perri, D.; Medici, M.A.; Foti, M.; Orlandella, B.M.; Mastino, A. The gamma-2-herpesvirus bovine herpesvirus 4 causes apoptotic infection in permissive cell lines. Virology 2000, 277, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Cronin, J.; Goetze, L.; Donofrio, G.; Schuberth, H.J. Defining postpartum uterine disease and the mechanisms of infection and immunity in the female reproductive tract in cattle. Biol. Reprod. 2009, 81, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Pagnini, U.; Montagnaro, S.; Pacelli, F.; De Martino, L.; Florio, S.; Rocco, D.; Iovane, G.; Pacilio, M.; Gabellini, C.; Marsili, S.; et al. The involvement of oxidative stress in bovine herpesvirus type 4-mediated apoptosis. Front. Biosci. 2004, 9, 2106–2114. [Google Scholar] [CrossRef]
- Romeo, F.; Delgado, S.; Uriarte, E.L.; Storani, L.; Cuesta, L.M.; Morán, P.; Altamiranda, E.G.; Odeón, A.; Pérez, S.; Verna, A. Study of the dynamics of in vitro infection with bovine gammaherpesvirus type 4 and apoptosis markers in susceptible cells. Microb. Pathog. 2022, 169, 105645. [Google Scholar] [CrossRef]
- Tebaldi, G.; Jacca, S.; Montanini, B.; Capra, E.; Rosamilia, A.; Sala, A.; Stella, A.; Castiglioni, B.; Ottonello, S.; Donofrio, G. Virus-Mediated Metalloproteinase 1 Induction Revealed by Transcriptome Profiling of Bovine Herpesvirus 4-Infected Bovine Endometrial Stromal Cells. Biol. Reprod. 2016, 95, 12. [Google Scholar] [CrossRef]
- Romeo, F.; Louge-Uriarte, E.; Gonzalez-Altamiranda, E.; Delgado, S.; Pereyra, S.; Morán, P.; Odeón, A.; Pérez, S.; Verna, A.E. Gene expression and in vitro replication of bovine gammaherpesvirus type 4. Arch. Virol. 2021, 166, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Romeo, F.; Spetter, M.J.; Moran, P.; Pereyra, S.; Odeon, A.; Perez, S.E.; Verna, A.E. Analysis of the transcripts encoding for antigenic proteins of bovine gammaherpesvirus 4. J. Vet. Sci. 2020, 21, e5. [Google Scholar] [CrossRef] [PubMed]
- Verna, A.E.; Manrique, J.M.; Pérez, S.E.; Leunda, M.R.; Pereyra, S.B.; Jones, L.R.; Odeón, A.C. Genomic analysis of bovine herpesvirus type 4 (BoGHV4) from Argentina: High genetic variability and novel phylogenetic groups. Vet. Microbiol. 2012, 160, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Müench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Rensetti, D.E.; Marin, M.S.; Morán, P.E.; Odeón, A.C.; Verna, A.E.; Pérez, S.E. Bovine herpesvirus type 5 replication and induction of apoptosis in vitro and in the trigeminal ganglion of experimentally-infected cattle. Comp. Immunol. Microbiol. Infect. Dis. 2018, 57, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Sakumoto, R.; Hayashi, K.G.; Ogiso, T.; Kunii, H.; Shirozu, T.; Kim, S.W.; Bai, H.; Kawahara, M.; Kimura, K.; et al. Involvement of interferon-tau in the induction of apoptotic, pyroptotic, and autophagic cell death-related signaling pathways in the bovine uterine endometrium during early pregnancy. J. Reprod. Dev. 2018, 64, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Huang, J.J.; Dahms, H.U.; Zhen, J.J.; Ying, X.P. Cell damage and apoptosis in the hepatopancreas of Eriocheir sinensis induced by cadmium. Aquat. Toxicol. 2017, 190, 190–198. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Thomson, B.J. Viruses and apoptosis. Int. J. Exp. Pathol. 2001, 82, 65–76. [Google Scholar] [CrossRef]
- McGahon, A.J.; Martin, S.J.; Bissonnette, R.P.; Mahboubi, A.; Shi, Y.; Mogil, R.J.; Nishioka, W.K.; Green, D.R. The end of the (cell) line: Methods for the study of apoptosis in vitro. Methods Cell Biol. 1995, 46, 153–185. [Google Scholar] [CrossRef]
- Mesner, P.W.; Epting, C.L.; Hegarty, J.L.; Green, S.H. A timetable of events during programmed cell death induced by trophic factor withdrawal from neuronal PC12 cells. J. Neurosci. 1995, 15, 7357–7366. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, W.; Irie, S.; Sato, T.A. Morphological changes in the nucleus and actin cytoskeleton in the process of Fas-induced apoptosis in Jurkat T cells. Histochem. J. 2000, 32, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Daniel, B.; DeCoster, M.A. Quantification of sPLA2-induced early and late apoptosis changes in neuronal cell cultures using combined TUNEL and DAPI staining. Brain Res. Brain Res. Protoc. 2004, 13, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, A.M.; Hoda, F.E.; Noor, A.M. Studying the effect of LPS on cytotoxicity and apoptosis in PC12 neuronal cells: Role of Bax, Bcl-2, and Caspase-3 protein expression. Toxicol. Mech. Methods 2010, 20, 316–320. [Google Scholar] [CrossRef]
- Gillet, L.; Dewals, B.; Farnir, F.; de Leval, L.; Vanderplasschen, A. Bovine herpesvirus 4 induces apoptosis of human carcinoma cell lines in vitro and in vivo. Cancer Res. 2005, 65, 9463–9472. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romeo, F.; Delgado, S.; Yavorsky, M.; Martinez Cuesta, L.; Pereyra, S.; González Altamiranda, E.; Louge Uriarte, E.; Pérez, S.; Verna, A. Modulation of Apoptosis by Bovine Gammaherpesvirus 4 Infection in Bovine Endometrial Cells and the Possible Role of LPS in This Process. Biology 2024, 13, 249. https://doi.org/10.3390/biology13040249
Romeo F, Delgado S, Yavorsky M, Martinez Cuesta L, Pereyra S, González Altamiranda E, Louge Uriarte E, Pérez S, Verna A. Modulation of Apoptosis by Bovine Gammaherpesvirus 4 Infection in Bovine Endometrial Cells and the Possible Role of LPS in This Process. Biology. 2024; 13(4):249. https://doi.org/10.3390/biology13040249
Chicago/Turabian StyleRomeo, Florencia, Santiago Delgado, Marisol Yavorsky, Lucía Martinez Cuesta, Susana Pereyra, Erika González Altamiranda, Enrique Louge Uriarte, Sandra Pérez, and Andrea Verna. 2024. "Modulation of Apoptosis by Bovine Gammaherpesvirus 4 Infection in Bovine Endometrial Cells and the Possible Role of LPS in This Process" Biology 13, no. 4: 249. https://doi.org/10.3390/biology13040249