
Development and Application of a High-Throughput Method for the Purification and Analysis of Surface Carbohydrates from Klebsiella pneumoniae

 ,
,  , ,
, ,  , ,
, , 

Abstract
Simple Summary
Abstract
1. Introduction
2. Experimental Section
2.1. Chemicals
2.2. Culture Supernatants
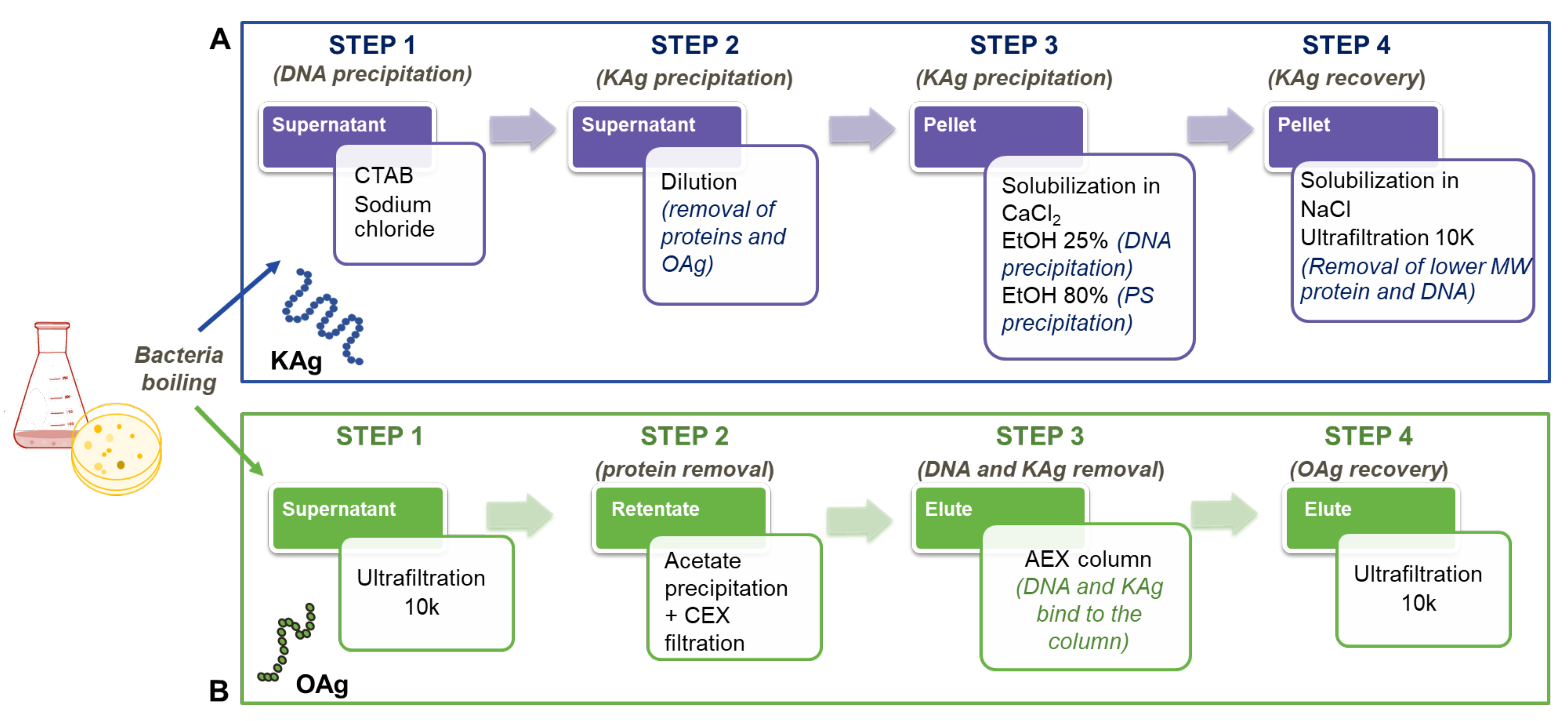
2.2.1. K-Ag Isolation
2.2.2. O-Ag Isolation
2.3. Polysaccharide Purification
2.3.1. K-Ag Purification
2.3.2. O-Ag Purification
2.4. Analytical Methods
2.4.1. Intermediate Characterization
2.4.2. High-Performance Liquid Chromatography–Size-Exclusion Chromatography (HPLC–SEC)
2.4.3. High-Performance Anion-Exchange Chromatography with Pulsed Amperometric Detection (HPAEC-PAD)
2.5. Sugar Composition and Linkage Analysis Using GLC and GLC-MS
2.6. NMR Analysis
3. Results
3.1. K-Ag Purification
3.2. OAg Purification
3.3. Purification Methods Developed Applied to the Structural Determination of New K- and O-Loci Saccharide Products
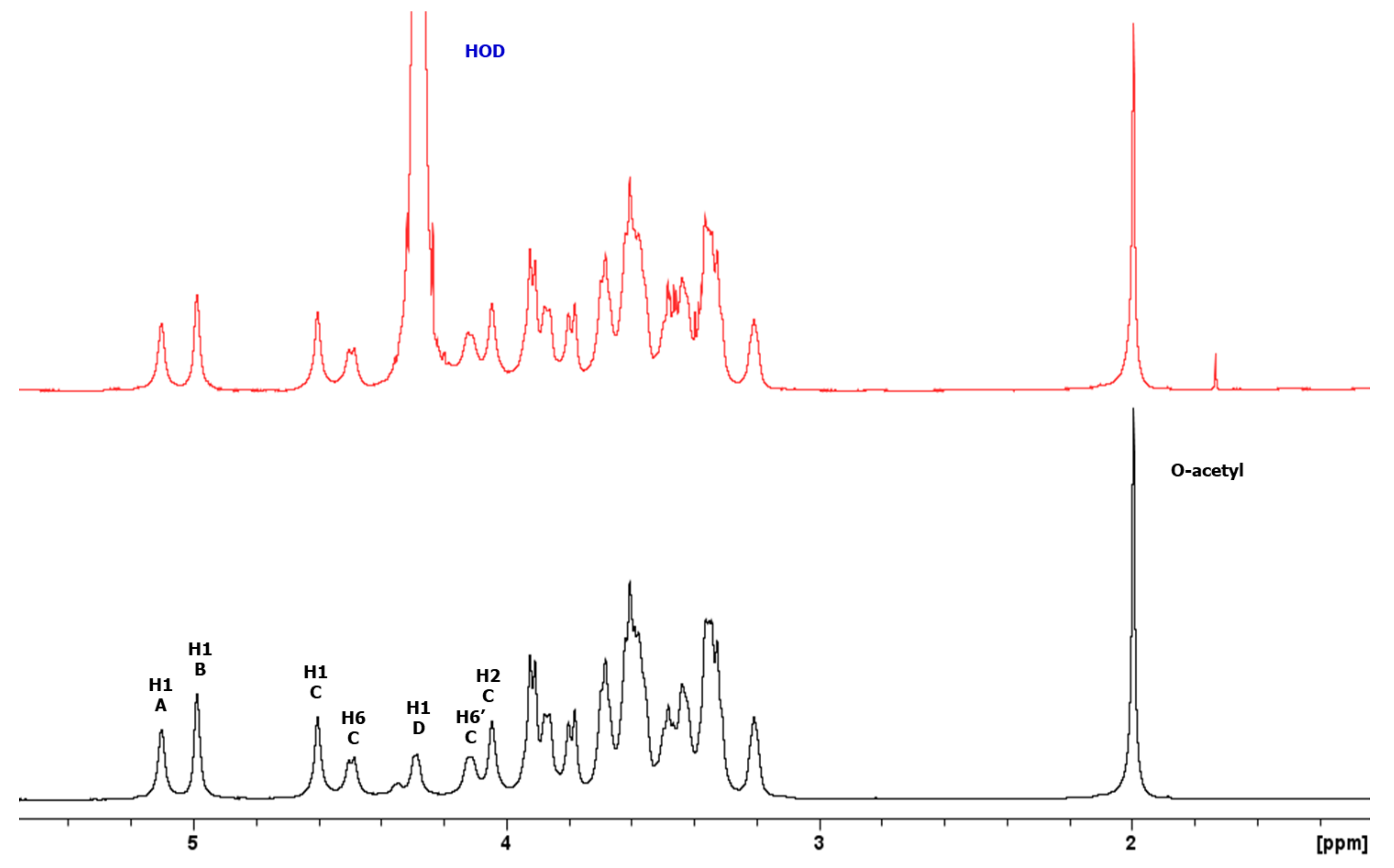
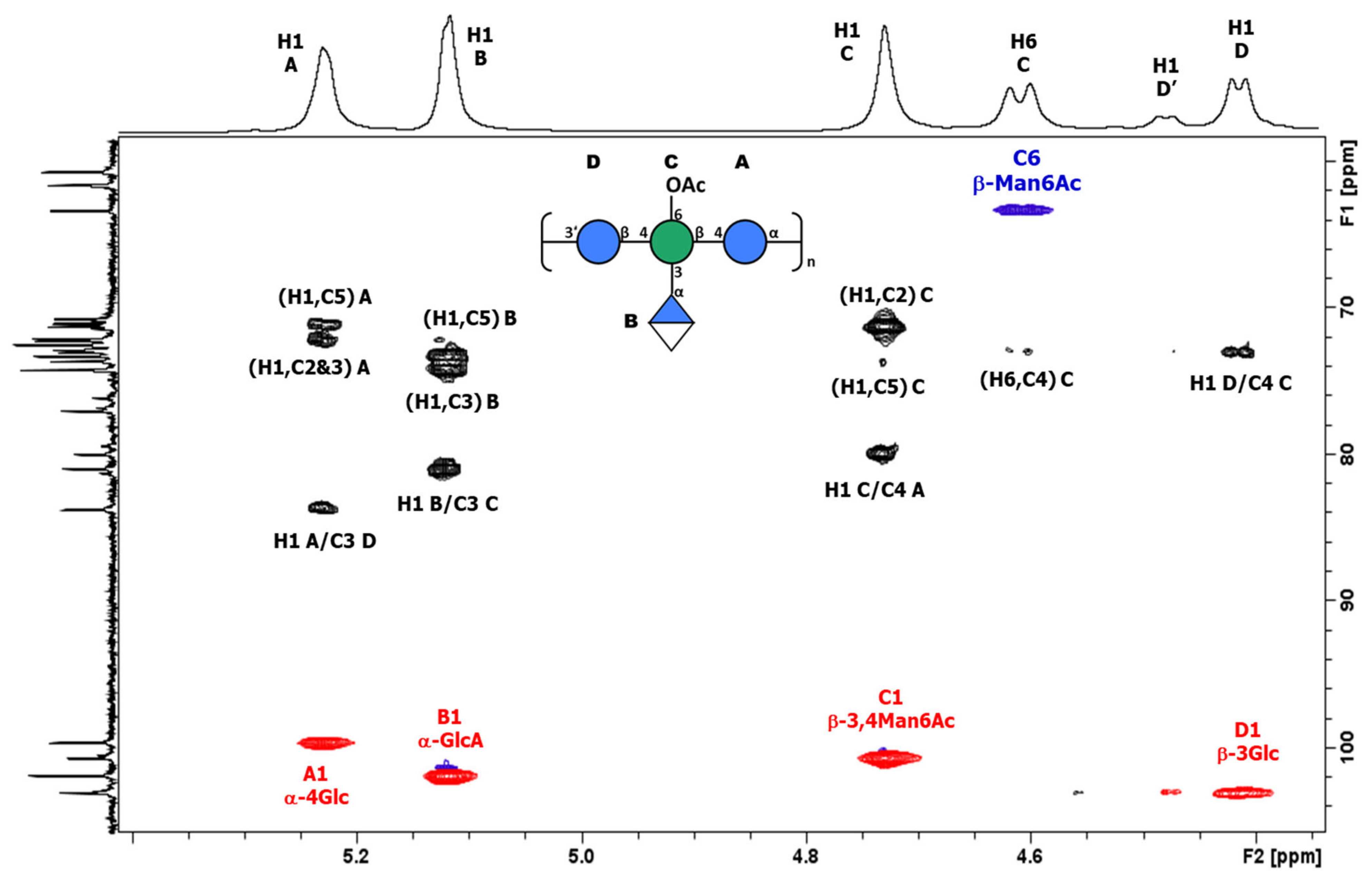
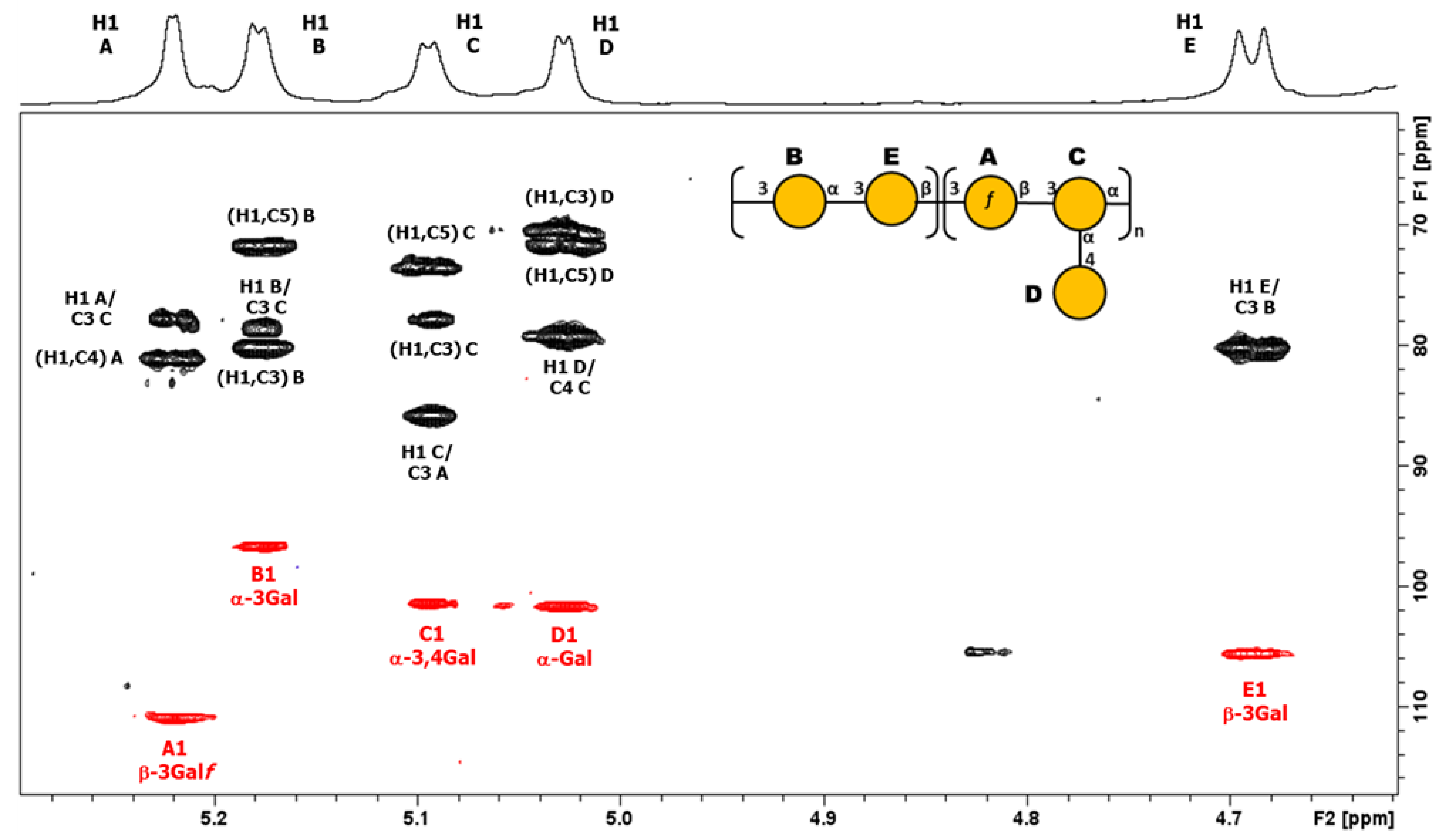
3.3.1. Structural Determination of K-Ag from Strain 4998 (KL107)

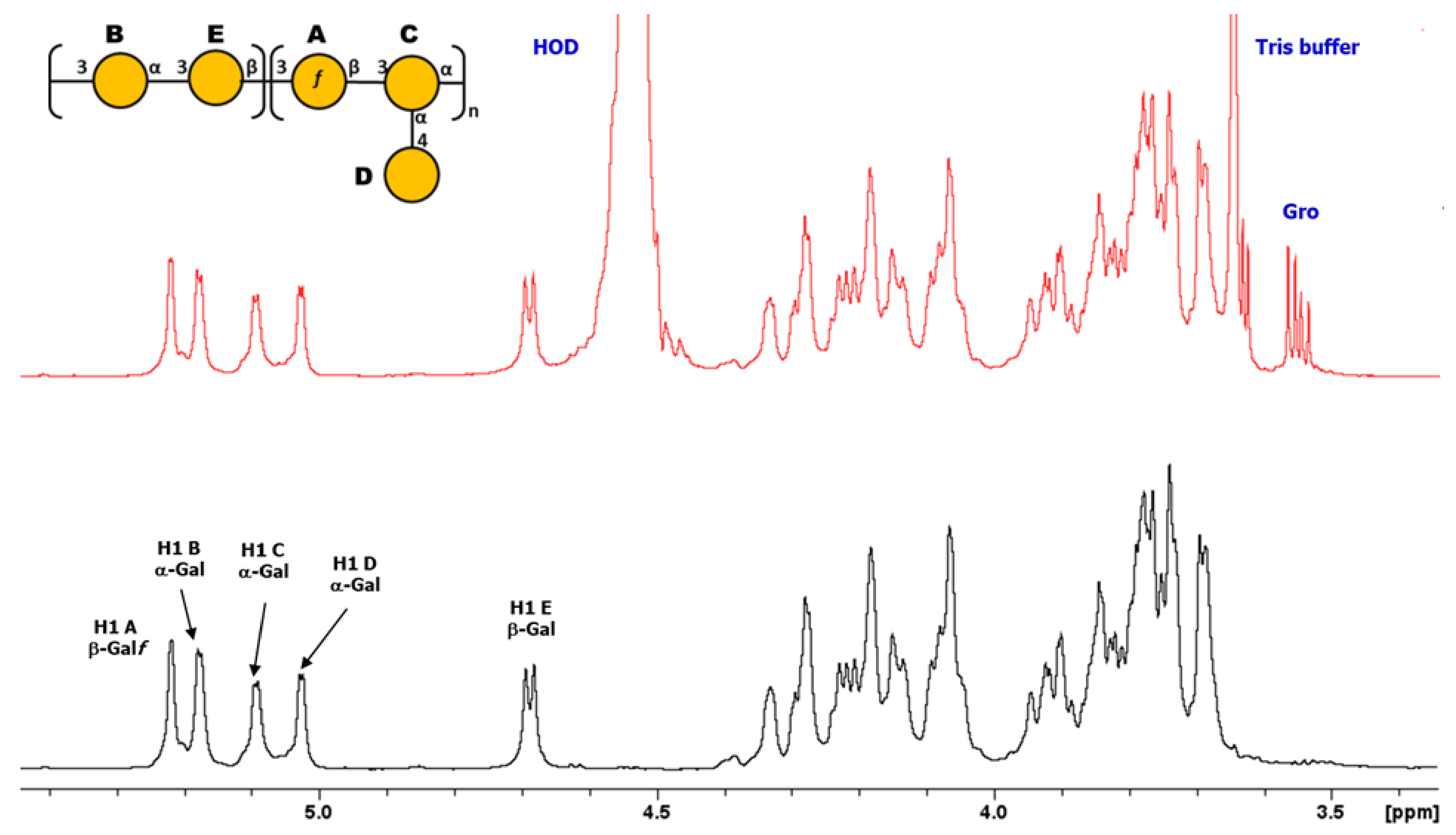
3.3.2. Structural Determination of O-Ag from Strain 7008B (OL102)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sands, K.; Carvalho, M.J.; Portal, E.; Thomson, K.; Dyer, C.; Akpulu, C.; Andrews, R.; Ferreira, A.; Gillespie, D.; Hender, T.; et al. Characterization of antimicrobial-resistant Gram-negative bacteria that cause neonatal sepsis in seven low- and middle-income countries. Nat. Microbiol. 2021, 6, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J.; et al. Multistate Point-Prevalence Survey of Health Care–Associated Infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef]
- Bassat, Q.; Blau, D.M.; Ogbuanu, I.U.; Samura, S.; Kaluma, E.; Bassey, I.A.; Sow, S.; Keita, A.M.; Tapia, M.D.; Mehta, A.; et al. Causes of Death Among Infants and Children in the Child Health and Mortality Prevention Surveillance (CHAMPS) Network. JAMA Netw. Open 2023, 6, e2322494. [Google Scholar] [CrossRef]
- Meatherall, B.L.; Gregson, D.; Ross, T.; Pitout, J.D.; Laupland, K.B. Incidence, Risk Factors, and Outcomes of Klebsiella pneumoniae Bacteremia. Am. J. Med. 2009, 122, 866–873. [Google Scholar] [CrossRef]
- Micoli, F.; Costantino, P.; Adamo, R. Potential targets for next generation antimicrobial glycoconjugate vaccines. FEMS Microbiol. Rev. 2018, 42, 388–423. [Google Scholar] [CrossRef]
- Tacconelli, N.M.E. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Choi, M.; Tennant, S.M.; Simon, R.; Cross, A.S. Progress towards the development of Klebsiella vaccines. Expert Rev. Vaccines 2019, 18, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Wick, R.R.; Gorrie, C.; Jenney, A.; Follador, R.; Thomson, N.R.; Holt, K.E. Identification of Klebsiella capsule synthesis loci from whole genome data. Microb. Genom. 2016, 2, e000102. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, R.A.; Bamert, R.S.; Tan, K.S.; Imbulgoda, U.; Barlow, C.K.; Taiaroa, G.; Pickard, D.J.; Schittenhelm, R.B.; Dougan, G.; Short, F.L.; et al. Epitopes in the capsular polysaccharide and the porin OmpK36 receptors are required for bacteriophage infection of Klebsiella pneumoniae. Cell Rep. 2023, 42, 112551. [Google Scholar] [CrossRef]
- Patro, L.P.P.; Rathinavelan, T. Targeting the Sugary Armor of Klebsiella Species. Front. Cell. Infect. Microbiol. 2019, 9, 367. [Google Scholar] [CrossRef]
- Rodrigues, C.; Sousa, C.; Lopes, J.A.; Novais, Â.; Peixe, L. A Front Line on Klebsiella pneumoniae Capsular Polysaccharide Knowledge: Fourier Transform Infrared Spectroscopy as an Accurate and Fast Typing Tool. mSystems 2020, 5, 00386. [Google Scholar] [CrossRef]
- Choi, P.H.; Jo, J.; Lin, Y.-C.; Lin, M.-H.; Chou, C.-Y.; Dietrich, L.E.P.; Tong, L. A distinct holoenzyme organization for two-subunit pyruvate carboxylase. Nat. Commun. 2016, 7, 12713. [Google Scholar] [CrossRef]
- Berti, F.; De Ricco, R.; Rappuoli, R. Role of O-Acetylation in the Immunogenicity of Bacterial Polysaccharide Vaccines. Molecules 2018, 23, 1340. [Google Scholar] [CrossRef]
- Yang, F.L.; Yang, Y.L.; Liao, P.C.; Chou, J.C.; Tsai, K.C.; Yang, A.S.; Sheu, F.; Lin, T.; Shieh, P.; Wang, J.; et al. Structure and immunological characterization of the capsular polysaccharide of a pyrogenic liver abscess caused by Klebsiella pneumoniae: Activation of macrophages through Toll-like receptor 4. J. Biol. Chem. 2011, 286, 21041–21051. [Google Scholar] [CrossRef]
- Hsu, C.-R.; Liao, C.-H.; Lin, T.-L.; Yang, H.-R.; Yang, F.-L.; Hsieh, P.-F.; Wu, S.-H.; Wang, J.-T. Identification of a capsular variant and characterization of capsular acetylation in Klebsiella pneumoniae PLA-associated type K57. Sci. Rep. 2016, 6, 31946. [Google Scholar] [CrossRef]
- Arato, V.; Raso, M.M.; Gasperini, G.; Scorza, F.B.; Micoli, F. Prophylaxis and Treatment against Klebsiella pneumoniae: Current Insights on This Emerging Anti-Microbial Resistant Global Threat. Int. J. Mol. Sci. 2021, 22, 4042. [Google Scholar] [CrossRef]
- Artyszuk, D.; Izdebski, R.; Maciejewska, A.; Kaszowska, M.; Herud, A.; Szijártó, V.; Gniadkowski, M.; Lukasiewicz, J. The Impact of Insertion Sequences on O-Serotype Phenotype and Its O-Locus-Based Prediction in Klebsiella pneumoniae O2 and O1. Int. J. Mol. Sci. 2020, 21, 6572. [Google Scholar] [CrossRef]
- Clarke, B.R.; Ovchinnikova, O.G.; Kelly, S.D.; Williamson, M.L.; Butler, J.E.; Liu, B.; Wang, L.; Gou, X.; Follador, R.; Lowary, T.L.; et al. Molecular basis for the structural diversity in serogroup O2-antigen polysaccharides in Klebsiella pneumoniae. J. Biol. Chem. 2018, 293, 4666–4679. [Google Scholar] [CrossRef]
- Stojkovic, K.; Szijártó, V.; Kaszowska, M.; Niedziela, T.; Hartl, K.; Nagy, G.; Lukasiewicz, J. Identification of d-Galactan-III As Part of the Lipopolysaccharide of Klebsiella pneumoniae Serotype O1. Front. Microbiol. 2017, 8, 684. [Google Scholar] [CrossRef]
- Guachalla, L.M.; Stojkovic, K.; Hartl, K.; Kaszowska, M.; Kumar, Y.; Wahl, B.; Paprotka, T.; Nagy, E.; Lukasiewicz, J.; Nagy, G.; et al. Discovery of monoclonal antibodies cross-reactive to novel subserotypes of K. pneumoniae O3. Sci. Rep. 2017, 7, 6635. [Google Scholar] [CrossRef]
- Vinogradov, E.; Frirdich, E.; MacLean, L.L.; Perry, M.B.; Petersen, B.O.; Duus, J.Ø.; Whitfield, C. Structures of lipopolysaccharides from Klebsiella pneumoniae. Eluicidation of the structure of the linkage region between core and polysaccharide O chain and identification of the residues at the non-reducing termini of the O chains. J. Biol. Chem. 2002, 277, 25070–25081. [Google Scholar] [CrossRef]
- Bellich, B.; Lagatolla, C.; Rizzo, R.; D’Andrea, M.M.; Rossolini, G.M.; Cescutti, P. Determination of the capsular polysaccharide structure of the Klebsiella pneumoniae ST512 representative strain KPB-1 and assignments of the glycosyltransferases functions. Int. J. Biol. Macromol. 2020, 155, 315–323. [Google Scholar] [CrossRef]
- Buffet, A.; Rocha, E.P.C.; Rendueles, O. Nutrient conditions are primary drivers of bacterial capsule maintenance in Klebsiella. Proc. R. Soc. B Biol. Sci. 2021, 288, 20202876. [Google Scholar] [CrossRef]
- Micoli, F.; Rondini, S.; Gavini, M.; Pisoni, I.; Lanzilao, L.; Colucci, A.; Giannelli, C.; Pippi, F.; Sollai, L.; Pinto, V.; et al. A scalable method for O-antigen purification applied to various Salmonella serovars. Anal. Biochem. 2013, 434, 136–145. [Google Scholar] [CrossRef]
- Kakehi, K.; Honda, S. Silyl Ethers of Carbohydrates. In Analysis of Carbohydrates by GLC and MS; CRC Press: Boca Raton, FL, USA, 2021; pp. 43–85. [Google Scholar]
- Harris, P.J.; Henry, R.J.; Blakeney, A.B.; Stone, B.A. An improved procedure for the methylation analysis of oligosaccharides and polysaccharides. Carbohydr. Res. 1984, 127, 59–73. [Google Scholar] [CrossRef]
- Albersheim, P.; Nevins, D.J.; English, P.D.; Karr, A. A method for the analysis of sugars in plant cell-wall polysaccharides by gas-liquid chromatography. Carbohydr. Res. 1967, 5, 340–345. [Google Scholar] [CrossRef]
- Sweet, D.P.; Shapiro, R.H.; Albersheim, P. Quantitative analysis by various g.l.c. response-factor theories for partially methylated and partially ethylated alditol acetates. Carbohydr. Res. 1975, 40, 217–225. [Google Scholar] [CrossRef]
- Micoli, F.; Giannelli, C.; Di Benedetto, R. O-Antigen Extraction, Purification, and Chemical Conjugation to a Carrier Protein. Methods Mol. Biol. 2021, 2183, 267–304. [Google Scholar]
- Gormus, B.J.; Wheat, R.W. Polysaccharides of type 6 Klebsiella. J. Bacteriol. 1971, 108, 1304–1309. [Google Scholar] [CrossRef]
- Scott, J.E. Aliphatic ammonium salts in the assay of acidic polysaccharides from tissues. Methods Biochem. Anal. 1960, 8, 145–197. [Google Scholar]
- Nath, K.; Chakraborty, A.K. Studies of the primary structure of the capsular polysaccharide from Klebsiella serotype K15. Carbohydr. Res. 1987, 161, 91–96. [Google Scholar] [CrossRef]
- Niemann, H.; Kwiatkowski, B.; Westphal, U.; Stirm, S. Klebsiella serotype 25 capsular polysaccharide: Primary structure and depolymerization by a bacteriophage-borne glycanase. J. Bacteriol. 1977, 130, 366–374. [Google Scholar] [CrossRef]
- Wick, R.R.; Heinz, E.; Holt, K.E.; Wyres, K.L. Kaptive web: User-Friendly capsule and lipopolysaccharide serotype prediction for Klebsiella genomes. J. Clin. Microbiol. 2018, 56, e00197-18. [Google Scholar] [CrossRef]
- Corsaro, M.M.; De Castro, C.; Naldi, T.; Parrilli, M.; Tomás, J.M.; Regué, M. 1H and 13C NMR characterization and secondary structure of the K2 polysaccharide of Klebsiella pneumoniae strain 52145. Carbohydr. Res. 2005, 340, 2212–2217. [Google Scholar] [CrossRef]
- Ovchinnikova, O.G.; Treat, L.P.; Teelucksingh, T.; Clarke, B.R.; Miner, T.A.; Whitfield, C.; Walker, K.A.; Miller, V.L. Hypermucoviscosity Regulator RmpD Interacts with Wzc and Controls Capsular Polysaccharide Chain Length. mBio 2023, 14, e0080023. [Google Scholar] [CrossRef]
- Taylor, A.W.; Blau, D.M.; Bassat, Q.; Onyango, D.; Kotloff, K.L.; El Arifeen, S.; Mandomando, I.; Chawana, R.; Baillie, V.L.; Akelo, V.; et al. Initial findings from a novel population-based child mortality surveillance approach: A descriptive study. Lancet Glob. Health 2020, 8, e909–e919. [Google Scholar] [CrossRef]
- Thomson, K.M.; Dyer, C.; Liu, F.; Sands, K.; Portal, E.; Carvalho, M.J.; Barrell, M.; Boostrom, I.; Dunachie, S.; Farzana, R.; et al. Effects of antibiotic resistance, drug target attainment, bacterial pathogenicity and virulence, and antibiotic access and affordability on outcomes in neonatal sepsis: An international microbiology and drug evaluation prospective substudy (BARNARDS). Lancet Infect. Dis. 2021, 21, 1677–1688. [Google Scholar] [CrossRef] [PubMed]
- Russell, N.J.; Stöhr, W.; Plakkal, N.; Cook, A.; Berkley, J.A.; Adhisivam, B.; Agarwal, R.; Ahmed, N.U.; Balasegaram, M.; Ballot, D.; et al. Patterns of antibiotic use, pathogens, and prediction of mortality in hospitalized neonates and young infants with sepsis: A global neonatal sepsis observational cohort study (NeoOBS). PLoS Med. 2023, 20, e1004179. [Google Scholar] [CrossRef] [PubMed]
- Kumar, C.K.; Sands, K.; Walsh, T.R.; O’Brien, S.; Sharland, M.; Lewnard, J.A.; Hu, H.; Srikantiah, P.; Laxminarayan, R. Global, regional, and national estimates of the impact of a maternal Klebsiella pneumoniae vaccine: A Bayesian modeling analysis. PLoS Med. 2023, 20, e1004239. [Google Scholar] [CrossRef]
- Cryz, S.J.; Fürer, E.; Germanier, R. Purification and vaccine potential of Klebsiella capsular polysaccharides. Infect. Immun. 1985, 50, 225–230. [Google Scholar] [CrossRef]
- Gormus, B.J.; Wheat, R.W.; Porter, J.F. Occurrence of Pyruvic Acid in Capsular Polysaccharides from Various Klebsiella Species. J. Bacteriol. 1971, 107, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Biemans, R.; Micoli, F.; Romano, M.R. Glycoconjugate vaccines, production and characterization. Recent Trends Carbohydr. Chem. 2020, 2, 285–313. [Google Scholar]
- Micoli, F.; Del Bino, L.; Alfini, R.; Carboni, F.; Romano, M.R.; Adamo, R. Glycoconjugate vaccines: Current approaches towards faster vaccine design. Expert Rev. Vaccines 2019, 18, 881–895. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Intermediates | Sugar Step Yield % | Protein % | DNA % | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| K2 | K15 | K64 | K64† | K2 | K15 | K64 | K64 † | K2 | K15 | K64 | K64 † | |
| Step 1: Supernatant after DNA precipitation with CTAB | 87 * | 75 * | 103 * | 89 * | 12.2 | 9.3 | 194 | 48.3 | 82.0 | 7.7 | 10.6 | 12.1 |
| Step 2: Pellet solubilized in CaCl2/NaCl # | 88 | 75 | 62 | 92 | 6.8 | 7.1 | 11.8 | 5.0 | 62.5 | 5.6 | 1.8 | 3.0 |
| Step 3: Pellet post EtOH precipitation | 80 | 56 | 94 | 58 | 5.5 | 5.8 | 2.8 | 3.8 | 3.6 | 1.8 | 1.1 | 5.1 |
| Step 4: Final product post 10 K and 0.22 μm filtration | 71 | 96 | 107 | 102 | 2.0 | 1.6 | 0.6 | 1.1 | 0.4 | 0.7 | 0.1 | 0.7 |
| KAg Type | Extraction Method | Amount Obtained * | Molecular Weight | OAc Level % |
|---|---|---|---|---|
| K2 | H2O | 29 mg | 530 kDa | ~55% |
| AcOH | 36 mg | 309 kDa | ~45% | |
| K62 | H2O | 68 mg | 357 kDa | na |
| AcOH | 74 mg | 160 kDa | na | |
| K64 | H2O | 11 mg | 1390 kDa | ~65% |
| AcOH | 12 mg | 391 kDa | ~63% | |
| K15 | AcOH | 50 mg | 58 kDa | ~97% |
| Strain | K-Locus | Structure | K-Ag Purity % | |
|---|---|---|---|---|
| NCTC9135 | K15 |  | [32] | 98% |
| NCTC9145 | K25 |  | [33] | 99% |
| 12641B | K102 | Unknown | 99% | |
| 17648 | K105 | 99% | ||
| 27870B | K108 | 97% | ||
| 19679 | K112 | 99% | ||
| 24745-2 | K116 | 97% | ||
| 10405 | K117 | 98% | ||
| 773 | K122 | 97% | ||
| 30405 | K128 | 98% | ||
| 34473 | K136 | 97% | ||
| Strain | O-locus | Structure | O-Ag Purity % | |
| 7008B | O1v2 |  | 99% | |
| 1217 | O3a |  | [20] | 99% |
| 24649 | O3b |  | [20] | 99% |
| 25827 | O4 |  | [21] | 98% |
| 29867 | O5 |  | [18] | 98% |
 Glucose (Glc);
Glucose (Glc);  Galactose (Gal);
Galactose (Gal);  Mannose (Man);
Mannose (Man);  Glucuronic Acid (GlcA);
Glucuronic Acid (GlcA);  Ribose (Ribf).
Ribose (Ribf).Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nonne, F.; Molfetta, M.; Nappini, R.; La Guidara, C.; Di Benedetto, R.; Mfana, S.; Bellich, B.; Raso, M.M.; Gasperini, G.; Alfini, R.; et al. Development and Application of a High-Throughput Method for the Purification and Analysis of Surface Carbohydrates from Klebsiella pneumoniae. Biology 2024, 13, 256. https://doi.org/10.3390/biology13040256
Nonne F, Molfetta M, Nappini R, La Guidara C, Di Benedetto R, Mfana S, Bellich B, Raso MM, Gasperini G, Alfini R, et al. Development and Application of a High-Throughput Method for the Purification and Analysis of Surface Carbohydrates from Klebsiella pneumoniae. Biology. 2024; 13(4):256. https://doi.org/10.3390/biology13040256
Chicago/Turabian StyleNonne, Francesca, Mariagrazia Molfetta, Rebecca Nappini, Chiara La Guidara, Roberta Di Benedetto, Siwaphiwe Mfana, Barbara Bellich, Maria Michelina Raso, Gianmarco Gasperini, Renzo Alfini, and et al. 2024. "Development and Application of a High-Throughput Method for the Purification and Analysis of Surface Carbohydrates from Klebsiella pneumoniae" Biology 13, no. 4: 256. https://doi.org/10.3390/biology13040256
APA StyleNonne, F., Molfetta, M., Nappini, R., La Guidara, C., Di Benedetto, R., Mfana, S., Bellich, B., Raso, M. M., Gasperini, G., Alfini, R., Cescutti, P., Berlanda Scorza, F., Ravenscroft, N., Micoli, F., & Giannelli, C. (2024). Development and Application of a High-Throughput Method for the Purification and Analysis of Surface Carbohydrates from Klebsiella pneumoniae. Biology, 13(4), 256. https://doi.org/10.3390/biology13040256