


Toxicity of Ammonia Stress on the Physiological Homeostasis in the Gills of Litopenaeus vannamei under Seawater and Low-Salinity Conditions



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Shrimp Materials
2.2. Ammonia Stress Experiment and Sampling
2.3. Histological Analysis
2.4. Biochemical Analysis
2.5. Gene Expression Analysis
2.6. Statistical Analysis
3. Results
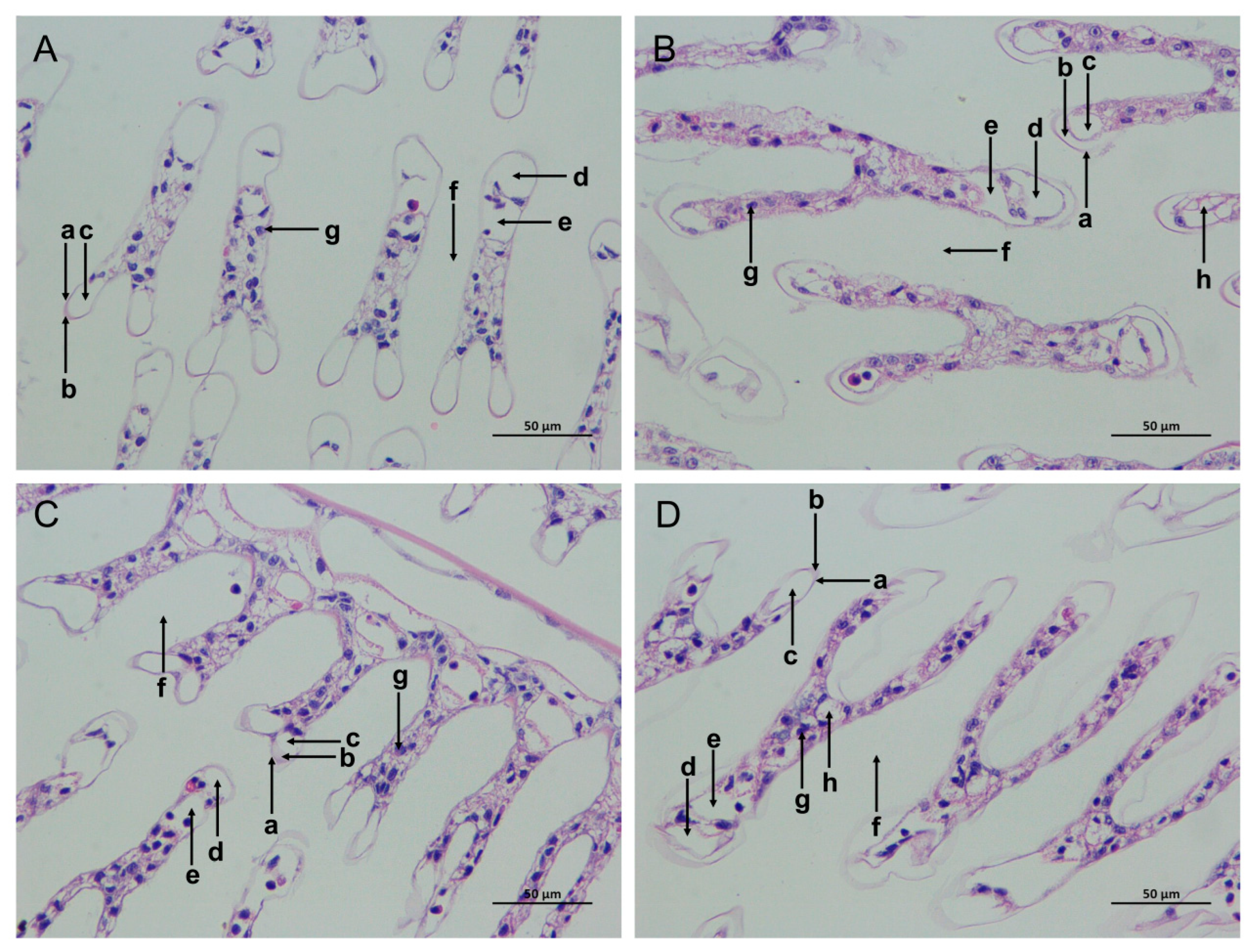
3.1. The Histological Morphology Changes in the Shrimp Gills
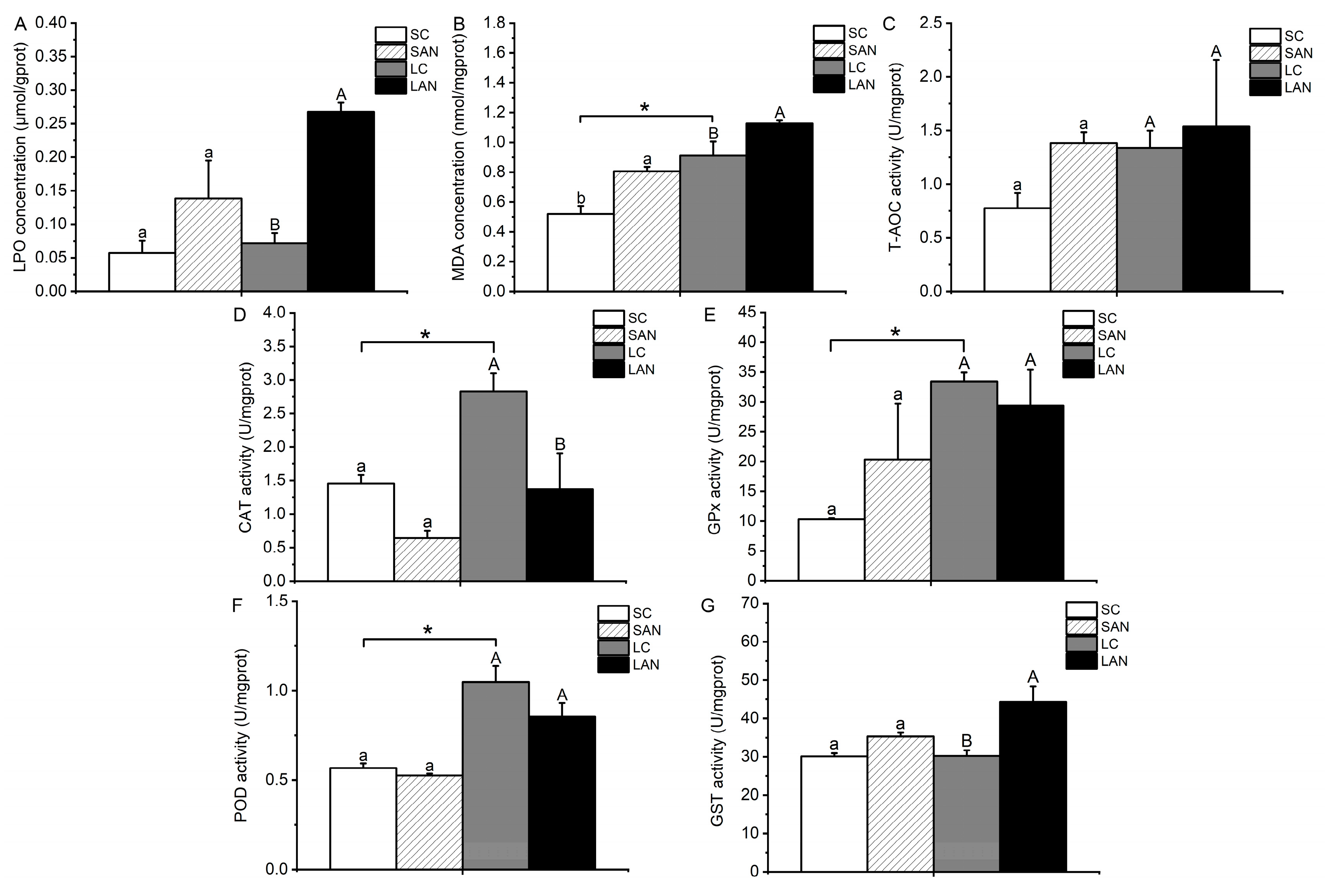
3.2. Changes in Oxidative Stress Biochemical Parameters in the Shrimp Gills
3.3. Changes in Antioxidant-Relevant Gene Expression in the Shrimp Gills
3.4. Changes in ER Stress-Relevant Genes’ Expression in the Shrimp Gills
3.5. Changes in Apoptosis-Relevant Genes’ Expression in the Shrimp Gills
3.6. Changes in Detoxification-Relevant Genes’ Expression in the Shrimp Gills
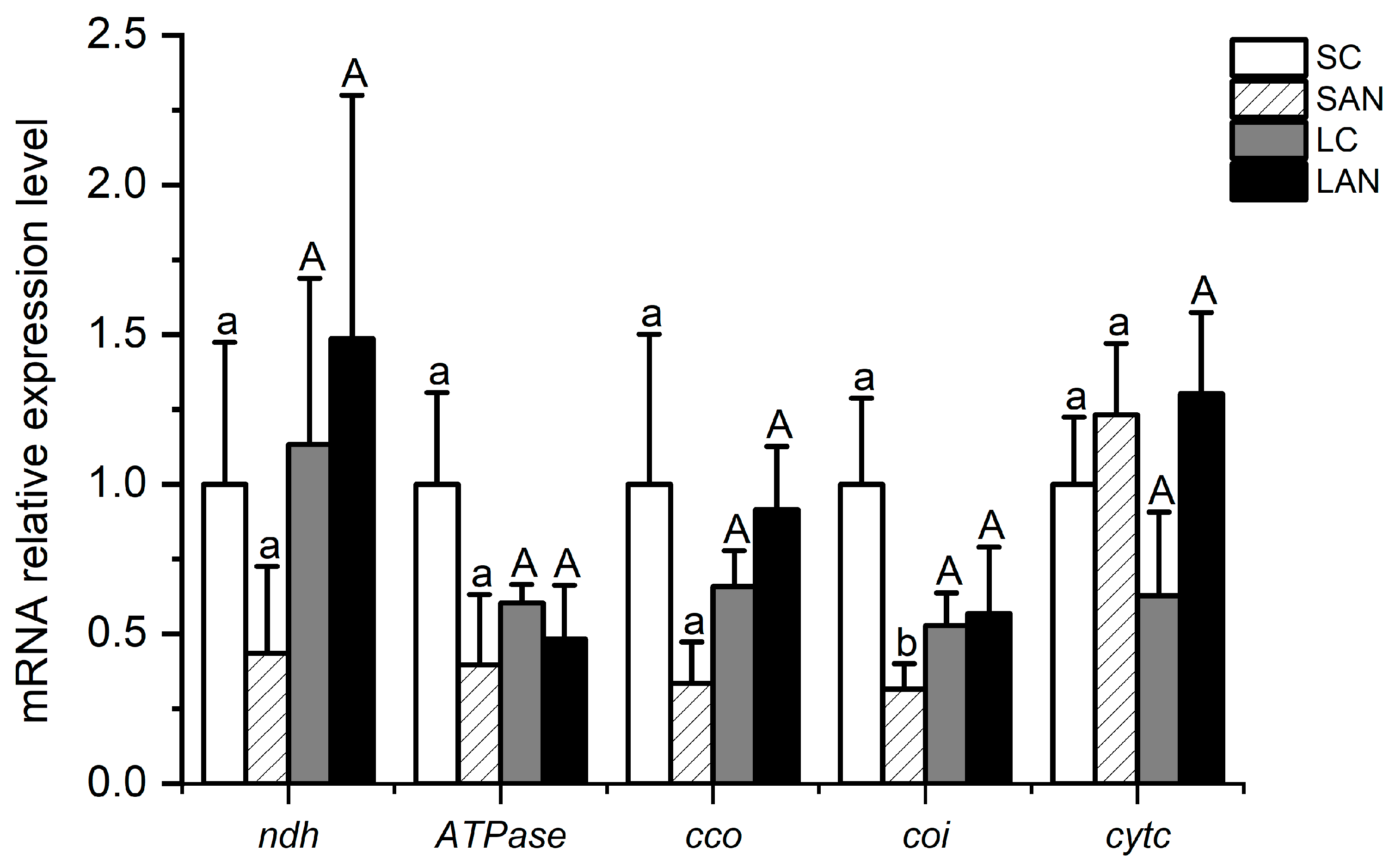
3.7. Changes in Energy Metabolism-Related Genes’ Expression in the Shrimp Gills
3.8. Changes in Osmoregulation-Relevant Genes’ Expression in the Shrimp Gills
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ogle, J.T.; Beaugez, K.; Lotz, J.M. Effects of salinity on survival and growth of postlarval Penaeus vannamei. Gulf Res. Rep. 1992, 8, 415–421. [Google Scholar] [CrossRef]
- Duan, Y.; Liu, Q.; Wang, Y.; Zhang, J.; Xiong, D. Impairment of the intestine barrier function in Litopenaeus vannamei exposed to ammonia and nitrite stress. Fish Shellfish. Immunol. 2018, 78, 279–288. [Google Scholar] [CrossRef]
- Yan, X.; Chen, Y.; Dong, X.; Tan, B.; Liu, H.; Zhang, S.; Chi, S.; Yang, Q.; Liu, H.; Yang, Y. Ammonia toxicity induces oxidative stress, inflammatory response and apoptosis in hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ E. lanceolatu). Front. Mar. Sci. 2021, 8, 667432. [Google Scholar] [CrossRef]
- Chang, Z.W.; Chiang, P.C.; Cheng, W.; Chang, C.C. Impact of ammonia exposure on coagulation in white shrimp, Litopenaeus vannamei. Ecotoxicol. Environ. Saf. 2015, 118, 98–102. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Z.; Wu, Z.; Peng, R.; Jiang, X.; Han, Q.; Jiang, M. Effect of ammonia nitrogen on the detoxification metabolic pathway of cuttlefish (Sepia pharaonis). Aquaculture 2022, 553, 738133. [Google Scholar] [CrossRef]
- Cheng, S.-Y.; Shieh, L.-W.; Chen, J.-C. Changes in hemolymph oxyhemocyanin, acid–base balance, and electrolytes in Marsupenaeus japonicus under combined ammonia and nitrite stress. Aquat. Toxicol. 2013, 130–131, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, S.; Yu, Y.; Sun, M.; Xiang, J.; Li, F. Effects of ammonia stress on the hemocytes of the Pacific white shrimp Litopenaeus vannamei. Chemosphere 2020, 239, 124759. [Google Scholar] [CrossRef]
- Zhao, M.; Yao, D.; Li, S.; Zhang, Y.; Aweya, J.J. Effects of ammonia on shrimp physiology and immunity: A review. Rev. Aquac. 2020, 12, 2194–2211. [Google Scholar] [CrossRef]
- Mekata, T.; Sudhakaran, R.; Okugawa, S.; Kono, T.; Sakai, M.; Itami, T. Molecular cloning and transcriptional analysis of a newly identified anti-lipopolysaccharide factor gene in kuruma shrimp, Marsupenaeus japonicus. Lett. Appl. Microbiol. 2010, 50, 112–119. [Google Scholar] [CrossRef]
- Kathyayani, S.A.; Poornima, M.; Sukumaran, S.; Nagavel, A.; Muralidhar, M. Effect of ammonia stress on immune variables of Pacific white shrimp Penaeus vannamei under varying levels of pH and susceptibility to white spot syndrome virus. Ecotoxicol. Environ. Saf. 2019, 184, 109626. [Google Scholar] [CrossRef]
- Liang, Z.; Liu, R.; Zhao, D.; Wang, L.; Sun, M.; Wang, M.; Song, L. Ammonia exposure induces oxidative stress, endoplasmic reticulum stress and apoptosis in hepatopancreas of pacific white shrimp (Litopenaeus vannamei). Fish Shellfish. Immunol. 2016, 54, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Cui, Y.; Wang, R.; Chen, Y.; Zhao, N.; Wang, C.; Wang, Z.; Li, Y. Combined effects of high salinity and ammonia-N exposure on the energy metabolism, immune response, oxidative resistance and ammonia metabolism of the Pacific white shrimp Litopenaeus vannamei. Aquac. Rep. 2021, 20, 100648. [Google Scholar] [CrossRef]
- Ou, H.; Liang, J.; Liu, J. Effects of acute ammonia exposure on oxidative stress, endoplasmic reticulum stress and apoptosis in the kuruma shrimp (Marsupenaeus japonicus). Aquac. Rep. 2022, 27, 101383. [Google Scholar] [CrossRef]
- Shan, H.; Geng, Z.; Ma, S.; Wang, T. Comparative study of the key enzymes and biochemical substances involved in the energy metabolism of Pacific white shrimp, Litopenaeus vannamei, with different ammonia-N tolerances. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 221, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Sui, Z.; Wei, C.; Wang, X.; Zhou, H.; Liu, C.; Mai, K.; He, G. Nutrient sensing signaling and metabolic responses in shrimp Litopenaeus vannamei under acute ammonia stress. Ecotoxicol. Environ. Saf. 2023, 253, 114672. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Nan, F.H. Effect of ambient ammonia on ammonia-N excretion and ATPase activity of Penaeus chinensis. Aquat. Toxicol. 1992, 23, 1–10. [Google Scholar]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P.; Gilmour, K.M.; Perry, S.F.; Kolosov, D.; Kelly, S.P.; Gerber, L.; Jensen, F.B.; Madsen, S.S.; et al. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.H. The fish gill: Site of action and model for toxic effects of environmental pollutants. Environ. Health. Perspect. 1987, 71, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Xiong, D.; Wang, Y.; Li, H.; Dong, H.; Zhang, J. Toxic effects of ammonia and thermal stress on the intestinal microbiota and transcriptomic and metabolomic responses of Litopenaeus vannamei. Sci. Total. Environ. 2021, 754, 141867. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Xing, Y.; Zhu, X.; Duan, Y.; Huang, J.; Nan, Y.; Zhang, J. Toxic effects of nitrite and microplastics stress on histology, oxidative stress, and metabolic function in the gills of Pacific white shrimp, Litopenaeus vannamei. Mar. Pollut. Bull. 2023, 187, 114531. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, J.; Wang, Y.; Liu, Q.; Xiong, D. Nitrite stress disrupts the structural integrity and induces oxidative stress response in the intestines of Pacific white shrimp Litopenaeus vannamei. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2018, 329, 43–50. [Google Scholar] [CrossRef]
- Castillo, C.; Hernández, J.; Valverde, I.; Pereira, V.; Sotillo, J.; Alonso, M.L.; Benedito, J. Plasma malonaldehyde (MDA) and total antioxidant status (TAS) during lactation in dairy cows. Res. Veter. Sci. 2006, 80, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Atli, G.; Canli, M. Response of antioxidant system of freshwater fish Oreochromis niloticus to acute and chronic metal (Cd, Cu, Cr, Zn, Fe) exposures. Ecotoxicol. Environ. Saf. 2010, 73, 1884–1889. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, J.; Dong, H.; Wang, Y.; Liu, Q.; Li, H. Oxidative stress response of the black tiger shrimp Penaeus monodon to Vibrio parahaemolyticus challenge. Fish Shellfish. Immunol. 2015, 46, 354–365. [Google Scholar] [CrossRef]
- Yu, C.; Xiao, J.-H. The Keap1-Nrf2 system: A mediator between oxidative stress and aging. Oxidative Med. Cell. Longev. 2021, 2021, 6635460. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Fu, M.; Zhao, C.; Zhou, F.; Yang, Q.; Qiu, L. Characterization and function analysis of Hsp60 and Hsp10 under different acute stresses in black tiger shrimp, Penaeus monodon. Cell Stress Chaperon. 2016, 21, 295–312. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Zhang, R.-R.; Zhao, X.-F.; Wang, J.-X. A thioredoxin response to the WSSV challenge on the Chinese white shrimp, Fenneropenaeus chinensis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 151, 92–98. [Google Scholar] [CrossRef]
- Hetz, C. The unfolded protein response: Controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 89–102. [Google Scholar] [CrossRef]
- Walter, P.; Ron, D. The unfolded protein response: From stress pathway to homeostatic regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef]
- Al-Masri, A. Apoptosis and long non-coding RNAs: Focus on their roles in Heart diseases. Pathol. Res. Pract. 2023, 251, 154889. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Kim, C.N.; Yang, J.; Jemmerson, R.; Wang, X. Induction of apoptotic program in cell-free extracts: Requirement for dATP and cytochrome c. Cell 1996, 86, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Miura, M. Regulatory roles of JNK in programmed cell death. J. Biochem. 2004, 136, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Nan, Y.; Zhu, X.; Huang, J.; Zhang, Z.; Xing, Y.; Yang, Y.; Xiao, M.; Duan, Y. Toxic effects of triclocarban on the histological morphology, phys-iological and immune response in the gills of the black tiger shrimp Penaeus monodon. Mar. Environ. Res. 2023, 192, 106245. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, S.; Yang, G.; Wang, X.; Liu, F.; Li, Y.; Chen, Y.; Zhou, T.; Xie, D.; Liu, Y.; et al. Energy metabolism: A critical target of cardiovascular injury. Biomed. Pharmacother. 2023, 165, 115271. [Google Scholar] [CrossRef] [PubMed]
- An, F.; Chang, W.; Song, J.; Zhang, J.; Li, Z.; Gao, P.; Wang, Y.; Xiao, Z.; Yan, C. Reprogramming of glucose metabolism: Metabolic alterations in the progression of osteosarcoma. J. Bone Oncol. 2024, 44, 100521. [Google Scholar] [CrossRef]
- Gopal, K.; Abdualkader, A.M.; Li, X.; Greenwell, A.A.; Karwi, Q.G.; Altamimi, T.R.; Saed, C.; Uddin, G.M.; Darwesh, A.M.; Jamieson, K.L.; et al. Loss of muscle PDH induces lactic acidosis and adaptive anaplerotic compensation via pyruvate-alanine cycling and glutaminolysis. J. Biol. Chem. 2023, 299, 105375. [Google Scholar] [CrossRef]
- Kay, J.; Weitzman, P.D.J. Krebs Citric Acid Cycle: Half a Century and Still Turning; Biochemical Society; Portland Press Ltd.: Colchester, UK, 1987; Volume 54, pp. 1–198. [Google Scholar]
- Zhang, Y.; Jiao, X.; Liu, J.; Feng, G.; Luo, X.; Zhang, M.; Zhang, B.; Huang, L.; Long, Q. A new direction in Chinese herbal medicine ameliorates for type 2 diabetes mellitus: Focus on the potential of mitochondrial respiratory chain complexes. J. Ethnopharmacol. 2024, 321, 117484. [Google Scholar] [CrossRef]
- Maclean, A.E.; Hayward, J.A.; Huet, D.; van Dooren, G.G.; Sheiner, L. The mystery of massive mitochondrial complexes: The apicomplexan respiratory chain. Trends Parasitol. 2022, 38, 1041–1052. [Google Scholar] [CrossRef]
- Fernández-Vizarra, E.; Ugalde, C. Cooperative assembly of the mitochondrial respiratory chain. Trends Biochem. Sci. 2022, 47, 999–1008. [Google Scholar] [CrossRef]
- Fernández-Vizarra, E.; López-Calcerrada, S.; Sierra-Magro, A.; Pérez-Pérez, R.; Formosa, L.E.; Hock, D.H.; Illescas, M.; Peñas, A.; Brischigliaro, M.; Ding, S.; et al. Two independent respiratory chains adapt OXPHOS performance to glycolytic switch. Cell Metab. 2022, 34, 1792–1808.e6. [Google Scholar] [CrossRef] [PubMed]
- Shamraj, O.I.; Lingrel, J.B. A putative fourth Na+, K+-ATPase alpha-subunit gene is expressed in testis. Proc. Natl. Acad. Sci. USA 1994, 91, 12952–12956. [Google Scholar] [CrossRef]
- Post, R.L.; Jolly, P.C. The linkage of sodium, potassium, and ammonium active transport across the human erythrocyte mem-brane. Biochim. Biophys. Acta 1957, 25, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Esbaugh, A.J.; Perry, S.F.; Bayaa, M.; Georgalis, T.; Nickerson, J.; Tufts, B.L.; Gilmour, K.M. Cytoplasmic carbonic anhydrase isozymes in rainbow trout Oncorhynchus mykiss: Comparative physiology and molecular evolution. J. Exp. Biol. 2005, 208, 1951–1961. [Google Scholar] [CrossRef] [PubMed]
- Echevarría, M.; Windhager, E.E.; Frindt, G. Selectivity of the renal collecting duct water channel aquaporin-3. J. Biol. Chem. 1996, 271, 25079–25082. [Google Scholar] [CrossRef]
- Yang, B.; Verkman, A.S. Water and glycerol permeabilities of aquaporins 1–5 and MIP determined quantitatively by expression of epitope-tagged constructs in Xenopus oocytes. J. Biol. Chem. 1997, 272, 16140–16146. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nan, Y.; Xiao, M.; Duan, Y.; Yang, Y. Toxicity of Ammonia Stress on the Physiological Homeostasis in the Gills of Litopenaeus vannamei under Seawater and Low-Salinity Conditions. Biology 2024, 13, 281. https://doi.org/10.3390/biology13040281
Nan Y, Xiao M, Duan Y, Yang Y. Toxicity of Ammonia Stress on the Physiological Homeostasis in the Gills of Litopenaeus vannamei under Seawater and Low-Salinity Conditions. Biology. 2024; 13(4):281. https://doi.org/10.3390/biology13040281
Chicago/Turabian StyleNan, Yuxiu, Meng Xiao, Yafei Duan, and Yukai Yang. 2024. "Toxicity of Ammonia Stress on the Physiological Homeostasis in the Gills of Litopenaeus vannamei under Seawater and Low-Salinity Conditions" Biology 13, no. 4: 281. https://doi.org/10.3390/biology13040281
APA StyleNan, Y., Xiao, M., Duan, Y., & Yang, Y. (2024). Toxicity of Ammonia Stress on the Physiological Homeostasis in the Gills of Litopenaeus vannamei under Seawater and Low-Salinity Conditions. Biology, 13(4), 281. https://doi.org/10.3390/biology13040281