
Obstetrical Constraints and the Origin of Extended Postnatal Brain Maturation in Hominin Evolution
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Data
2.2. Statistical Analyses
2.3. Pelvic Meshes
2.4. Fetal Model
2.5. Inclusion of Fossil Dyads in the Canonical Discriminant Analyses
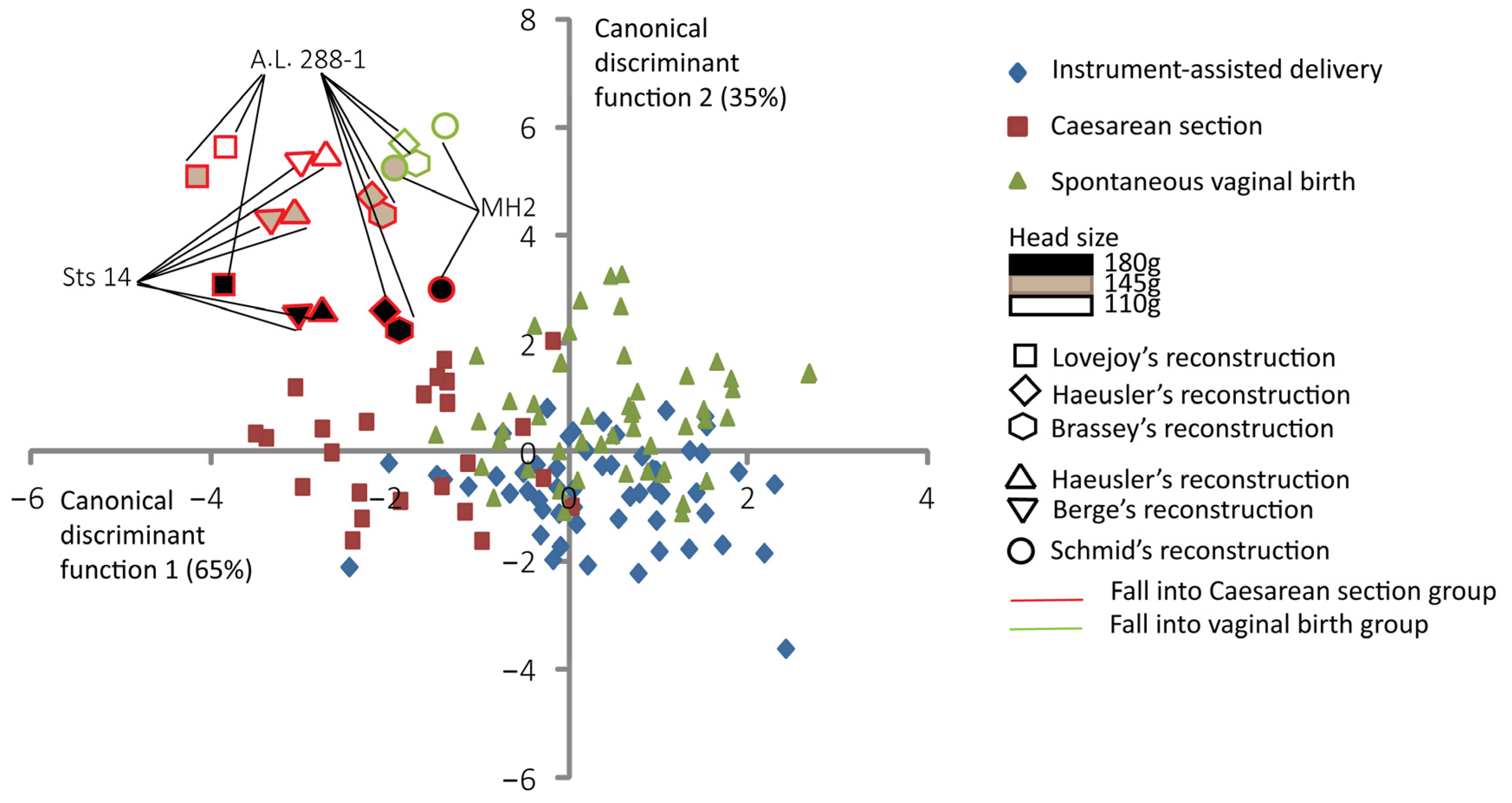
3. Results
4. Discussion
4.1. Discriminating Delivery Outcomes with Pelvic and Fetal Variables
4.2. Specific Pelvic Pattern Related to Inlet Arrest
4.3. Implication for Life History in Early Hominins
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, B.H.; Tompkins, L.R. Toward a life history of the Hominidae. Annu. Rev. Anthropol. 1995, 24, 257–279. [Google Scholar] [CrossRef]
- Hartwig, W.C. Perinatal life history traits in New World monkeys. Am. J. Primatol. 1996, 40, 99–130. [Google Scholar] [CrossRef]
- Hinde, K.; Milligan, L.A. Primate Milk: Proximate mechanisms and ultimate perspectives. Evol. Anthropol. 2011, 20, 9–23. [Google Scholar] [CrossRef]
- Portmann, A. Die Tragzeiten der Primaten und die Dauer der Schwangerschaft beim Menschen: Ein Problem der vergleichenden Biologie. Rev. Suisse Zool. 1941, 48, 511–518. [Google Scholar]
- Martin, R.D. Human brain evolution in an ecological context. In Proceedings of the 52nd James Arthur Lecture on the Evolution of the Human Brain; American Museum of Natural History, New York, NY, USA, 27 April 1982. [Google Scholar]
- Rosenberg, K.R. The evolution of human infancy: Why it helps to be helpless. Annu. Rev. Anthropol. 2021, 50, 423–440. [Google Scholar] [CrossRef]
- Coqueugniot, H.; Hublin, J.-J.; Veillon, F.; Houët, F.; Jacob, T. Early brain growth in Homo erectus and implications for cognitive ability. Nature 2004, 431, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Hrdy, S.B. Evolutionary Context of Human Development. In Attachment and Bonding: A New Synthesis; The MIT Press: Cambridge, MA, USA, 2006; pp. 9–32. [Google Scholar]
- Berge, C.; Goularas, D. A new reconstruction of Sts 14 pelvis (Australopithecus africanus) from computed tomography and three-dimensional modeling techniques. J. Hum. Evol. 2010, 58, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Tague, R.G.; Lovejoy, C.O. The obstetric pelvis of AL 288-1 (Lucy). J. Hum. Evol. 1986, 15, 237–255. [Google Scholar] [CrossRef]
- Frémondière, P.; Thollon, L.; Marchal, F.; Fornai, C.; Webb, N.M.; Haeusler, M. Dynamic finite-element simulations reveal early origin of complex human birth pattern. Commun. Biol. 2022, 5, 377. [Google Scholar] [CrossRef]
- Zhang, J.; Troendle, J.; Reddy, U.M.; Laughon, S.K.; Branch, D.W.; Burkman, R.; Landy, H.J.; Hibbard, J.U.; Haberman, S.; Ramirez, M.M.; et al. Contemporary cesarean delivery practice in the United States. Am. J. Obstet. Gynecol. 2010, 203, 326.e1–326.e10. [Google Scholar] [CrossRef]
- Dolea, C.; AbouZahr, C. Global burden of obstructed labour in the year 2000. In Global Burden of Disease 2000; World Health Organ: Geneva, Switzerland, 2003; pp. 1–17. [Google Scholar]
- Harper, L.M.; Odibo, A.O.; Stamilio, D.M.; Macones, G.A. Radiographic measures of the mid pelvis to predict cesarean delivery. Am. J. Obstet. Gynecol. 2013, 208, 460.e1–460.e6. [Google Scholar] [CrossRef] [PubMed]
- Lenhard, M.S.; Johnson, T.R.; Weckbach, S.; Nikolaou, K.; Friese, K.; Hasbargen, U. Pelvimetry revisited: Analyzing cephalopelvic disproportion. Eur. J. Radiol. 2010, 74, e107–e111. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.A.; Thurnau, G.R. Efficacy of the fetal-pelvic index in nulliparous women at risk for fetal-pelvic disproportion. Am. J. Obstet. Gynecol. 1992, 166, 810–814. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, U.; Taipale, P.; Heinonen, S. Fetal pelvic index to predict cephalopelvic disproportion—A retrospective clinical cohort study. Acta Obstet. Gynecol. Scand. 2015, 94, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Ruff, C.B. Morphological adaptation to climate in modern and fossil hominids. Am. J. Phys. Anthropol. 1994, 37, 65–107. [Google Scholar] [CrossRef]
- Betti, L.; von Cramon-Taubadel, N.; Manica, A.; Lycett, S.J. The interaction of neutral evolutionary processes with climatically-driven adaptive changes in the 3D shape of the human os coxae. J. Hum. Evol. 2014, 73, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.C.K. Between Scylla and Charybdis: Renegotiating resolution of the ‘obstetric dilemma’ in response to ecological change. Phil. Trans. R. Soc. B 2015, 370, 20140067. [Google Scholar] [CrossRef] [PubMed]
- Fischer, B.; Mitteroecker, P. Covariation between human pelvis shape, stature, and head size alleviates the obstetric dilemma. Proc. Natl. Acad. Sci. USA 2015, 112, 5655–5660. [Google Scholar] [CrossRef] [PubMed]
- Torres-Tamayo, N.; Martelli, S.; Schlager, S.; García-Martínez, D.; Sanchis-Gimeno, J.A.; Mata-Escolano, F.; Nalla, S.; Ogihara, N.; Oishi, M.; Bastir, M. Assessing thoraco-pelvic covariation in Homo sapiens and Pan troglodytes: A 3D geometric morphometric approach. Am. J. Phys. Anthropol. 2020, 173, 514–534. [Google Scholar] [CrossRef]
- DeSilva, J.M.; Laudicina, N.M.; Rosenberg, K.R.; Trevathan, W.R. Neonatal shoulder width suggests a semirotational, oblique birth mechanism in Australopithecus afarensis. Anat. Rec. 2017, 300, 890–899. [Google Scholar] [CrossRef]
- Weaver, T.D.; Hublin, J.J. Neandertal birth canal shape and the evolution of human childbirth. Proc. Natl. Acad. Sci. USA 2009, 106, 8151–8156. [Google Scholar] [CrossRef] [PubMed]
- Haeusler, M.; Grunstra, N.D.S.; Martin, R.D.; Krenn, V.A.; Fornai, C.; Webb, N.M. The obstetrical dilemma hypothesis: There’s life in the old dog yet. Biol. Rev. 2021, 96, 2031–2057. [Google Scholar] [CrossRef]
- Grunstra, N.D.S.; Betti, L.; Fischer, B.; Haeusler, M.; Pavlicev, M.; Stansfield, E.; Trevathan, W.; Webb, N.M.; Wells, J.C.K.; Rosenberg, K.R.; et al. There is an obstetrical dilemma: Misconceptions about the evolution of human childbirth and pelvic form. Am. J. Biol. Anthropol. 2023, 181, 535–544. [Google Scholar] [CrossRef]
- Abitbol, M.M. The shapes of the female pelvis. Contributing factors. J. Reprod. Med. 1996, 41, 242–250. [Google Scholar]
- Gunz, P.; Neubauer, S.; Maureille, B.; Hublin, J.J. Brain development after birth differs between Neanderthals and modern humans. Curr. Biol. 2010, 20, R921–R922. [Google Scholar] [CrossRef]
- DeSilva, J.M.; Lesnik, J.J. Brain size at birth throughout human evolution: A new method for estimating neonatal brain size in hominins. J. Hum. Evol. 2008, 55, 1064–1074. [Google Scholar] [CrossRef] [PubMed]
- Washburn, S.L. Tools and human evolution. Sci. Am. 1960, 203, 63–75. [Google Scholar] [CrossRef]
- Montagu, A. Neonatal and infant immaturity in man. JAMA 1961, 178, 56–57. [Google Scholar] [CrossRef]
- Gunz, P.; Neubauer, S.; Falk, D.; Tafforeau, P.; Le Cabec, A.; Smith, T.M.; Kimbel, W.H.; Spoor, F.; Alemseged, Z. Australopithecus afarensis endocasts suggest ape-like brain organization and prolonged brain growth. Sci. Adv. 2020, 6, eaaz4729. [Google Scholar] [CrossRef]
- Schaal, J.-P.; Maillet, R.; Riethmuller, D. Mécanique Obstétricale. In Traité d’obstétrique; Médecine-Sciences Flammarion: Paris, France, 2003; p. 1154. [Google Scholar]
- Tague, R.G. Commonalities in dimorphism and variability in the anthropoid pelvis, with implications for the fossil record. J. Hum. Evol. 1991, 21, 153–176. [Google Scholar] [CrossRef]
- Abitbol, M.M. Reconstruction of the Sts 14 (Australopithecus africanus) pelvis. Am. J. Phys. Anthropol. 1995, 96, 143–158. [Google Scholar] [CrossRef]
- Frémondière, P.; Thollon, L.; Adalian, P.; Delotte, J.; Marchal, F. Which foetal-pelvic variables are useful for predicting caesarean section and instrumental assistance. Med. Princ. Pract. 2017, 26, 359–367. [Google Scholar] [CrossRef]
- Abitbol, M.M. Ontogeny and evolution of pelvic diameters in anthropoid primates and in Australopithecus afarensis (AL 288-1). Am. J. Phys. Anthropol. 1991, 85, 135–148. [Google Scholar] [CrossRef]
- Tague, R.G. Variation in pelvic size between males and females. Am. J. Phys. Anthropol. 1989, 80, 59–71. [Google Scholar] [CrossRef]
- Frémondière, P.; Marchal, F.; Thollon, L.; Saliba-serre, B. Change in head shape of newborn infants in the week following birth: Contributing factors. J. Pediatr. Neurol. 2019, 17, 168–175. [Google Scholar] [CrossRef]
- Clarke, R.J.; Pickering, T.R.; Heaton, J.L.; Kuman, K. The earliest South African hominids. Annu. Rev. Anthropol. 2021, 50, 125–143. [Google Scholar] [CrossRef]
- Arsuaga, J.L.; Carretero, J.M. Multivariate analysis of the sexual dimorphism of the hip bone in a modern human population and in early hominids. Am. J. Phys. Anthropol. 1994, 93, 241–257. [Google Scholar] [CrossRef]
- Lovejoy, C.O. A reconstruction of the pelvis of AL-288 (Hadar Formation, Ethiopia). Am. J. Biol. Anthropol. 1979, 50, 460. [Google Scholar]
- Kibii, J.M.; Churchill, S.E.; Schmid, P.; Carlson, K.J.; Reed, N.D.; De Ruiter, D.J.; Berger, L.R. A partial pelvis of Australopithecus sediba. Science 2011, 333, 1407–1411. [Google Scholar] [CrossRef]
- Johanson, D.C.; Taieb, M.; Coppens, Y. Pliocene hominids from the Hadar formation, Ethiopia (1973–1977): Stratigraphic, chronologic, and paleoenvironmental contexts, with notes on hominid morphology and systematics. Am. J. Phys. Anthropol. 1982, 57, 373–402. [Google Scholar] [CrossRef]
- Cook, D.C.; Buikstra, J.E.; DeRousseau, C.J.; Johanson, D.C. Vertebral pathology in the Afar australopithecines. Am. J. Phys. Anthropol. 1983, 60, 83–101. [Google Scholar] [CrossRef]
- Haeusler, M.; Schmid, P. Comparison of the pelves of Sts 14 and AL 288-1: Implications for birth and sexual dimorphism in australopithecines. J. Hum. Evol. 1995, 29, 363–383. [Google Scholar] [CrossRef]
- Brassey, C.A.; O’Mahoney, T.G.; Chamberlain, A.T.; Sellers, W.I. A volumetric technique for fossil body mass estimation applied to Australopithecus afarensis. J. Hum. Evol. 2018, 115, 47–64. [Google Scholar] [CrossRef]
- Robinson, J.T. Early Hominid Posture and Locomotion; University of Chicago Press: Chicago, IL, USA, 1972; p. 361. [Google Scholar]
- Haeusler, M.; Ruff, C.B. Pelves. In Hominin Postcranial Remains from Sterkfontein, South Africa, 1936–1995; Zipfel, B., Richmond, B.G., Ward, C.V., Eds.; Oxford University Press: Oxford, UK, 2020; pp. 181–201. [Google Scholar] [CrossRef]
- Pickering, R.; Herries, A.I.R. A new multidisciplinary age of 2.61–2.07 Ma for the Sterkfontein member 4 australopiths. In Hominin Postcranial Remains from Sterkfontein, South Africa, 1936–1995; Zipfel, B., Richmond, B.G., Ward, C.V., Eds.; Oxford University Press: Oxford, UK, 2020; pp. 21–30. [Google Scholar]
- Berger, L.R.; de Ruiter, D.J.; Churchill, S.E.; Schmid, P.; Carlson, K.J.; Dirks, P.H.G.M.; Kibii, J.M. Australopithecus sediba: A new species of Homo-like australopith from South Africa. Science 2010, 328, 195–204. [Google Scholar] [CrossRef]
- Maharaj, D. Assessing cephalopelvic disproportion: Back to the basics. Obstet. Gynecol. Surv. 2010, 65, 387–395. [Google Scholar] [CrossRef]
- Ashton-Miller, J.A.; DeLancey, J.O.L. On the biomechanics of vaginal birth and common sequelae. Annu. Rev. Biomed. Eng. 2009, 11, 163–176. [Google Scholar] [CrossRef]
- Stewart, R. Pelvic Dimensions and the outcome of trial labour in Shona and Zulu primigravidas. S. Afr. Med. J. 1979, 55, 847–851. [Google Scholar]
- Abitbol, M.M. Evolution of the ischial spine and of the pelvic floor in the Hominoidea. Am. J. Biol. Anthropol. 1988, 75, 53–67. [Google Scholar] [CrossRef]
- Berge, C.; Orban-Segebarth, R.; Schmid, P. Obstetrical interpretation of the australopithecine pelvic cavity. J. Hum. Evol. 1984, 13, 573–587. [Google Scholar] [CrossRef]
- Laudicina, N.M.; Rodriguez, F.; DeSilva, J.M. Reconstructing birth in Australopithecus sediba. PLoS ONE 2019, 14, e0221871. [Google Scholar] [CrossRef]
- Claxton, A.G.; Hammond, A.S.; Romano, J.; Oleinik, E.; DeSilva, J.M. Virtual reconstruction of the Australopithecus africanus pelvis Sts 65 with implications for obstetrics and locomotion. J. Hum. Evol. 2016, 99, 10–24. [Google Scholar] [CrossRef]
- DeSilva, J.; Lesnik, J. Chimpanzee neonatal brain size: Implications for brain growth in Homo erectus. J. Hum. Evol. 2006, 51, 207–212. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Abbreviation | References |
|---|---|---|
| maximal transverse diameter of the inlet | mati | [33] |
| inlet antero-posterior diameter | iap | [33] |
| inlet sacral breadth | isb | [34] |
| posterior inlet diameter | pi | [35] |
| pectineal angle | pa | [36] |
| midplane angle | ma | [36] |
| midplane antero-posterior diameter | map | [37] |
| interspinous diameter | isp | [33] |
| posterior midplane diameter | pm | [34] |
| midplane sacral breadth | msb | [36] |
| subpubic angle | spa | [38] |
| transverse outlet diameter | tout | [33] |
| posterior outlet diameter | po | [37] |
| outlet antero-posterior diameter | oap | [33] |
| sacral chord length | scl | [33] |
| sacral chord subtense | scs | [33] |
| inlet–midplane angle | ima | [33] |
| submentobregmatic diameter | smb | [33] |
| suboccipitobregmatic diameter | sob | [33] |
| suboccipitofrontal diameter | sof | [33] |
| mentovertical diameter | mv | [33] |
| mento-occipital diameter | moc | [33] |
| occipitofrontal diameter | of | [33] |
| biparietal diameter | bip | [33] |
| bitemporal diameter | bit | [33] |
| right tragion–bregma diameter | rtb | [39] |
| left tragion–bregma diameter | ltb | [39] |
| suboccipitobregmatic circumference | sobc | [39] |
| head circumference | hc | [39] |
| biacromial diameter | bia | [36] |
| sternum–thoracic vertebral diameter | stv | [33] |
| abdominal circumference | abdc | [36] |
| abdominal sagittal diameter | abds | [36] |
| transverse abdominal diameter | abdt | [36] |
| bitrochanterian diameter | bih | [36] |
| birthweight | wght |
| Group | Predicted Group Membership | |||
|---|---|---|---|---|
| Caesarean | Spont. Vag. 1 | Instrument | Total | |
| Caesarean | 19 (79%) | 3 (13%) | 2 (8%) | 24 |
| Spont. Vag. | 4 (8%) | 32 (63%) | 15 (29%) | 51 |
| Instrument | 5 (9%) | 7 (12%) | 44 (79%) | 56 |
| “Dyads” | Predicted Group Membership | |||
|---|---|---|---|---|
| Pelvic Reconstructions | Head Size | Caesarean | Spont. Vag. 1 | Instrument |
| A.L. 288-1 Lovejoy [10] | 180 g | 94% | 5% | 1% |
| A.L. 288-1 Lovejoy [10] | 145 g | 91% | 8% | 1% |
| A.L. 288-1 Lovejoy [10] | 110 g | 85% | 15% | 0% |
| A.L. 288-1 Haeusler [44] | 180 g | 77% | 23% | 0% |
| A.L. 288-1 Haeusler [44] | 145 g | 67% | 33% | 0% |
| A.L. 288-1 Haeusler [44] | 110 g | 44% | 56% | 0% |
| A.L. 288-1 Brassey [46] | 180 g | 96% | 44% | 0% |
| A.L. 288-1 Brassey [46] | 145 g | 93% | 7% | 0% |
| A.L. 288-1 Brassey [46] | 110 g | 84% | 16% | 0% |
| Sts 14 Haeusler [44] | 180 g | 71% | 28% | 1% |
| Sts 14 Haeusler [44] | 145 g | 61% | 39% | 0% |
| Sts 14 Haeusler [44] | 110 g | 37% | 63% | 0% |
| Sts 14 Berge [9] 2 | 180 g | 92% | 8% | 0% |
| Sts 14 Berge [9] 2 | 145 g | 88% | 12% | 0% |
| Sts 14 Berge [9] 2 | 110 g | 83% | 17% | 0% |
| MH2 Schmid [50] | 180 g | 82% | 18% | 0% |
| MH2 Schmid [50] | 145 g | 75% | 25% | 0% |
| MH2 Schmid [50] | 110 g | 47% | 53% | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frémondière, P.; Haeusler, M.; Thollon, L.; Webb, N.M.; Marchal, F. Obstetrical Constraints and the Origin of Extended Postnatal Brain Maturation in Hominin Evolution. Biology 2024, 13, 398. https://doi.org/10.3390/biology13060398
Frémondière P, Haeusler M, Thollon L, Webb NM, Marchal F. Obstetrical Constraints and the Origin of Extended Postnatal Brain Maturation in Hominin Evolution. Biology. 2024; 13(6):398. https://doi.org/10.3390/biology13060398
Chicago/Turabian StyleFrémondière, Pierre, Martin Haeusler, Lionel Thollon, Nicole M. Webb, and François Marchal. 2024. "Obstetrical Constraints and the Origin of Extended Postnatal Brain Maturation in Hominin Evolution" Biology 13, no. 6: 398. https://doi.org/10.3390/biology13060398
APA StyleFrémondière, P., Haeusler, M., Thollon, L., Webb, N. M., & Marchal, F. (2024). Obstetrical Constraints and the Origin of Extended Postnatal Brain Maturation in Hominin Evolution. Biology, 13(6), 398. https://doi.org/10.3390/biology13060398