
The Response of Soil Respiration to Temperature and Humidity in the Thermokarst Depression Zone of the Headwater Wetlands of Qinghai Lake

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
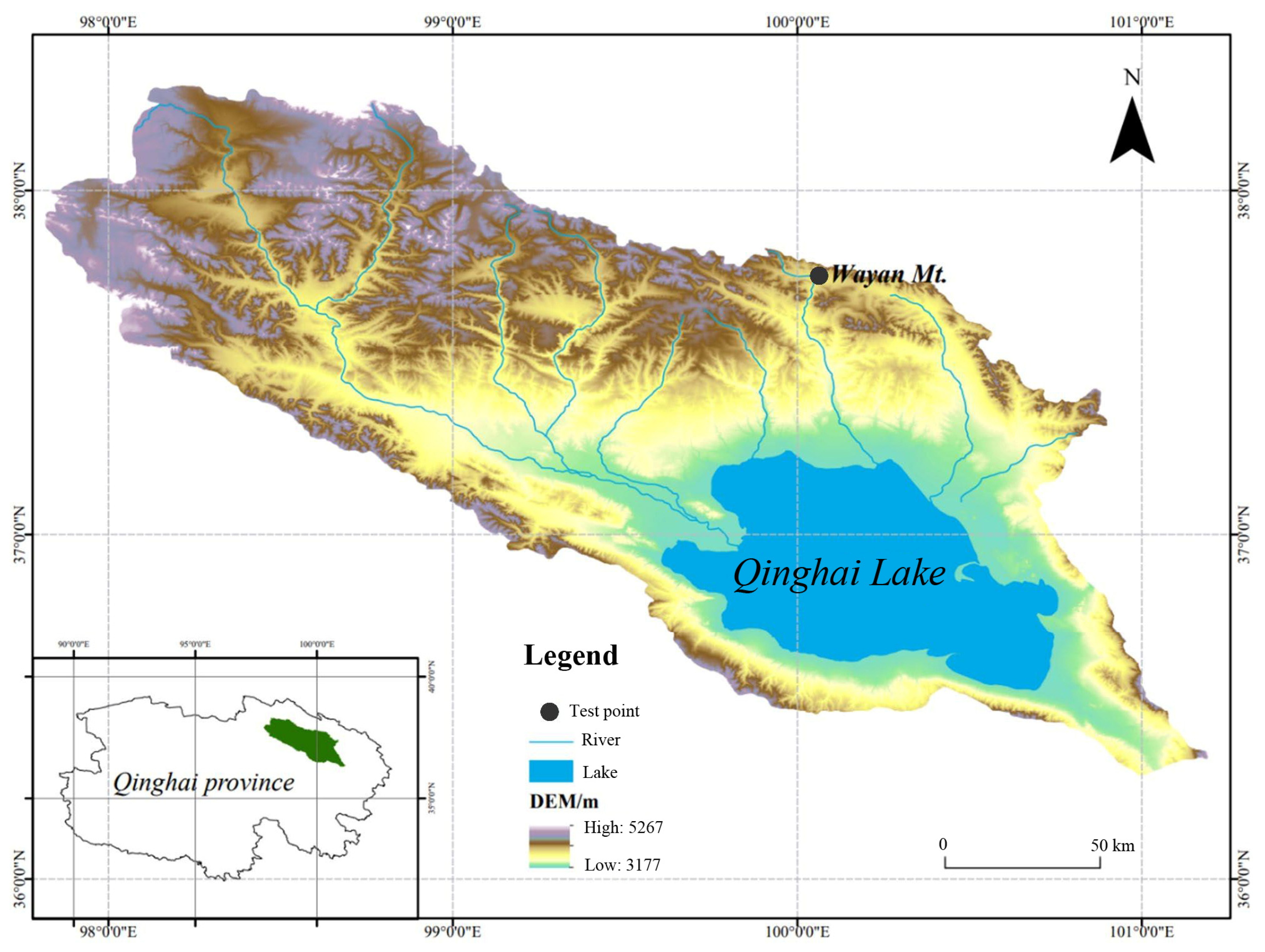
2.1. Experimental Site Description
2.2. Soil Respiration Measurement and Data Collection
2.3. Determination of Aboveground and Belowground Biomass and Soil Organic Matter
2.4. Statistical Analysis
2.5. Data Processing
2.5.1. Commonly Used Functional Equations for the Relationship between Soil Respiration Rate and Soil Temperature (Modified)
2.5.2. Common Functional Models for the Relationship between Soil Respiration Rate and Soil Volumetric Water Content
3. Results
3.1. Dynamic Characteristics of Soil Respiration in Thermokarst Areas
3.2. Soil Physicochemical Properties and Biomass in Thermokarst Areas
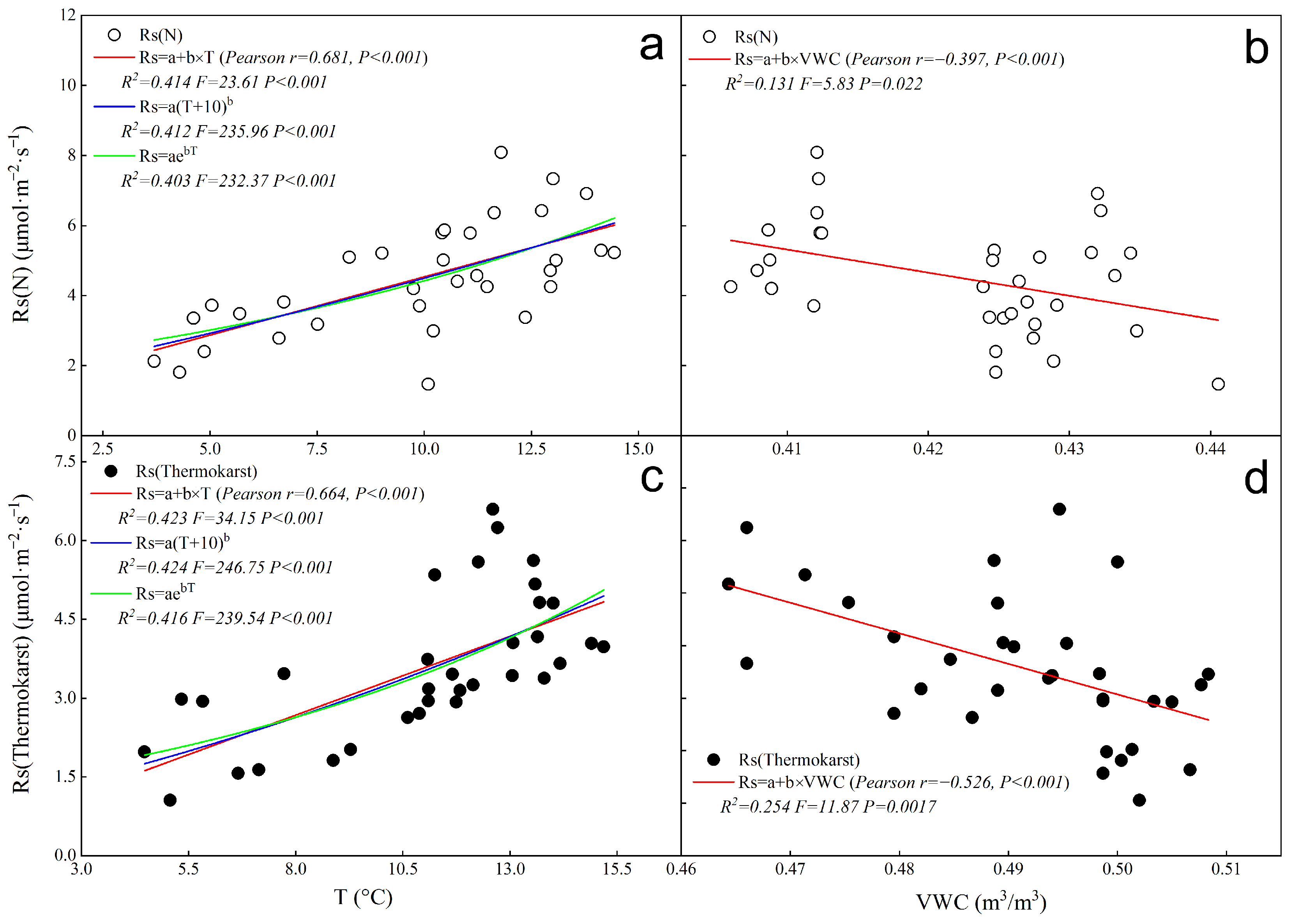
3.3. Fitting of Single-Factor Driven Models of Rs with Temperature and Humidity in the Thermokarst Subsidence Zone
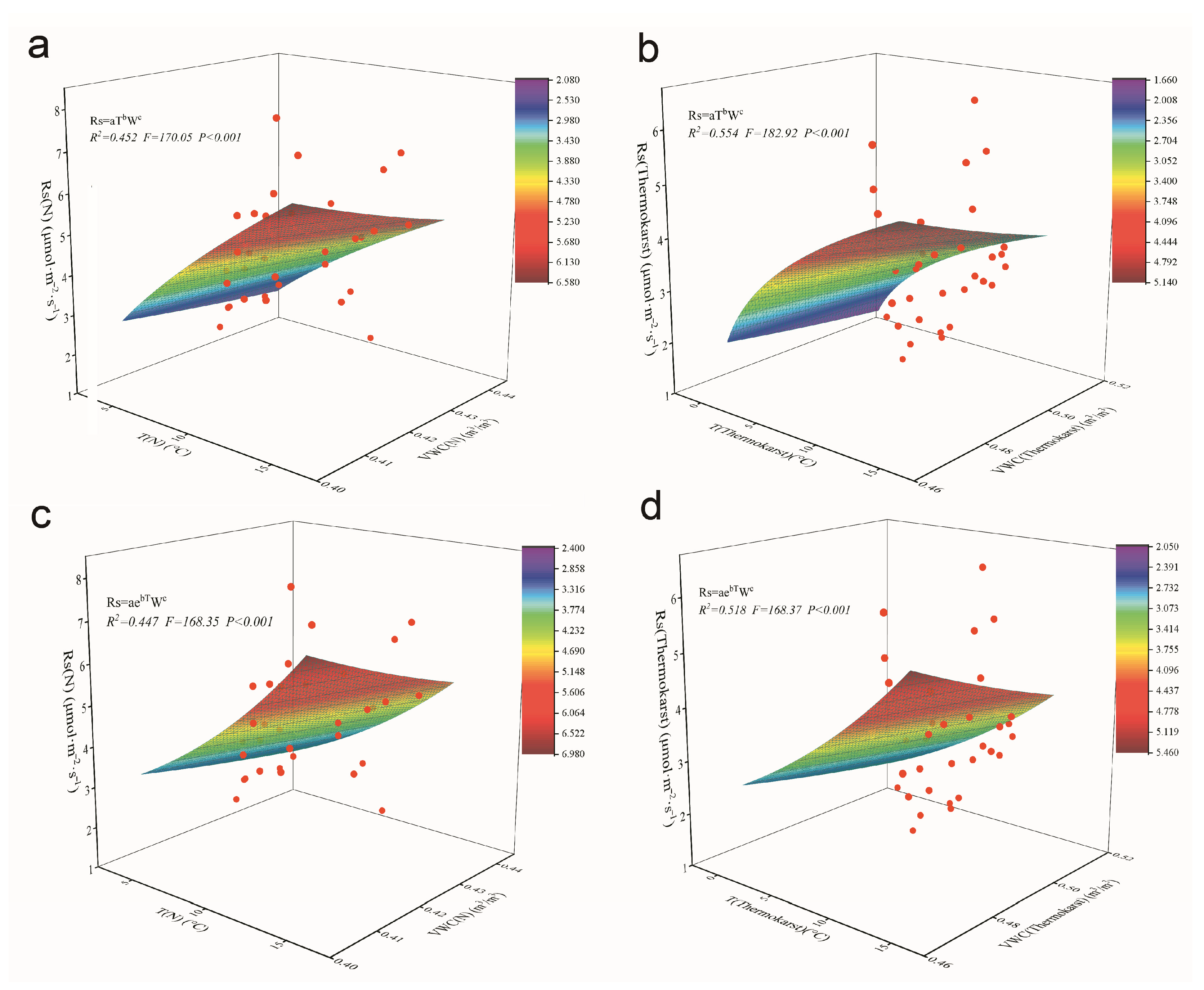
3.4. Fitting of Dual-Factor Driven Models of Rs with Temperature and Humidity in the Thermokarst Subsidence Zone
3.5. The Impact of Diurnal Variations in Thermokarst Regions on Soil Ecological Indicators
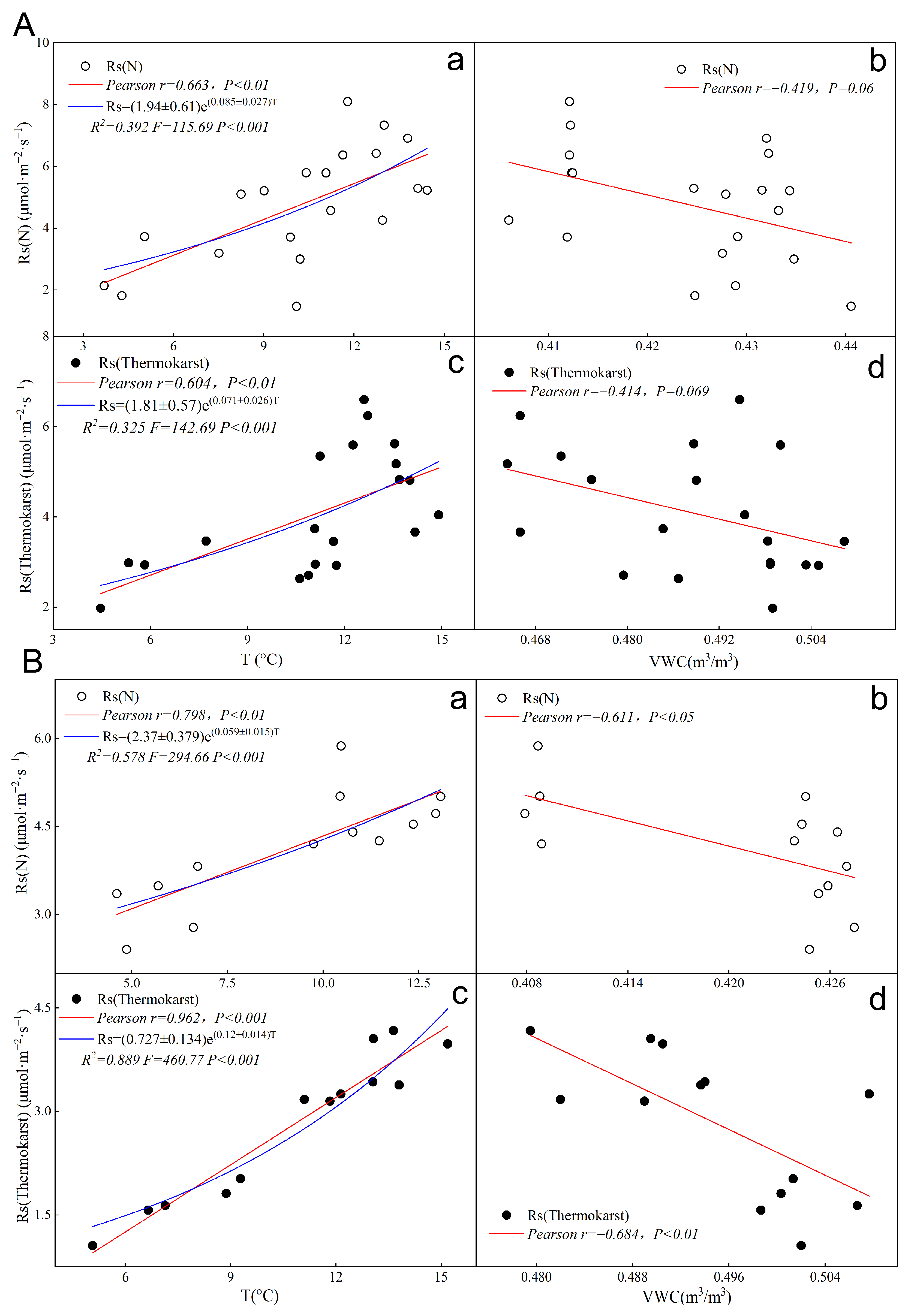
3.6. Fitting Single-Factor Models for Soil Respiration in Thermokarst and Natural Zones
4. Discussion
5. Conclusions
- (1)
- Temperature and Rs correlation: There is a significant positive correlation between soil temperature and Rs, with temperature demonstrating a higher explanatory power for Rs variations compared to volumetric water content.
- (2)
- Water content and Rs correlation: Volumetric water content shows a significant negative correlation with Rs. Notably, the inhibitory effect of water content on Rs is more pronounced in thermokarst depression zones than under natural conditions.
- (3)
- Driving factors for Rs: Single-factor models indicated that temperature-driven models have greater explanatory power for Rs variations than humidity-driven models. However, dual-factor models, which incorporate both temperature and humidity, provide an even more accurate explanation of Rs dynamics, particularly in thermokarst depression zones.
- (4)
- Day–night variations: During the day, temperature predominantly influences Rs under natural conditions, while increased water content inhibits Rs. At night, the correlation between temperature and Rs in thermokarst depression zones strengthens significantly, with a pronounced negative impact from increased water content.
- (5)
- Temperature sensitivity (Q10): The Q10 values for Rs in thermokarst depression zones (3.32) are higher than under natural conditions (1.80), indicating a greater sensitivity of Rs to temperature changes at night in these zones.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grosse, G.; Romanovsky, V.E.; Mann, D.H.; Jones, B. Spatial distribution of thermokarst terrain in Arctic Alaska. Geomorphology 2016, 273, 116–133. [Google Scholar] [CrossRef]
- Kang, S.; Xu, Y.; You, Q.; Flügel, W.-A.; Pepin, N.; Yao, T. Review of climate and cryospheric change in the Tibetan Plateau. Environ. Res. Lett. 2010, 5, 15101. [Google Scholar] [CrossRef]
- Morgenstern, A.; Overduin, P.P.; Gunther, F.; Stettner, S.; Ramage, J.; Schirrmeister, L.; Grigoriev, M.N.; Grosse, G. Thermo-erosional valleys in Siberian ice-rich permafrost. Permafr. Periglac. Process. 2021, 32, 59–75. [Google Scholar] [CrossRef]
- Sun, X.; Ren, G.; Ren, Y.; Lin, W.; Zhang, P.; Zhang, S.; Xue, X. Asian climate warming since 1901: Observation and simulation. Clim. Res. 2023, 91, 67–82. [Google Scholar] [CrossRef]
- Wan, C.; Gibson, J.J.; Shen, S.; Yi, Y.; Yi, P.; Yu, Z. Using stable isotopes paired with tritium analysis to assess thermokarst lake water balances in the Source Area of the Yellow River, northeastern Qinghai-Tibet Plateau, China. Sci. Total Environ. 2019, 689, 1276–1292. [Google Scholar] [CrossRef] [PubMed]
- Abbott, B.W.; Jones, J.B.; Godsey, S.E.; Larouche, J.R.; Bowden, W.B. Patterns and persistence of hydrologic carbon and nutrient export from collapsing upland permafrost. Biogeosciences 2015, 12, 3725–3740. [Google Scholar] [CrossRef]
- Mu, C.C.; Abbott, B.W.; Zhao, Q.; Su, H.; Wang, S.F.; Wu, Q.B.; Zhang, T.J.; Wu, X.D. Permafrost collapse shifts alpine tundra to a carbon source but reduces N2O and CH4 release on the northern Qinghai-Tibetan Plateau. Geophys. Res. Lett. 2017, 44, 8945–8952. [Google Scholar] [CrossRef]
- Chen, X.; Mu, C.; Jia, L.; Li, Z.; Wu, X. High-resolution dataset of thermokarst lakes on the Qinghai-Tibetan Plateau. Earth Syst. Sci. Data Discuss. 2021, 2021, 1–23. [Google Scholar] [CrossRef]
- Luo, J.; Niu, F.; Lin, Z.; Liu, M.; Yin, G.; Gao, Z. Abrupt increase in thermokarst lakes on the central Tibetan Plateau over the last 50 years. Catena 2022, 217, 106497. [Google Scholar] [CrossRef]
- Zhong, W. Investigation on Deformation of Thermokarst Terrain in Permafrost Regions over Heihe River Basin in Qilian Mountains. Master’s Thesis, Lanzhou Universiry, Lanzhou, China, 2019. [Google Scholar]
- Chen, X. Distribution and Regional Changes of Thermokarst lakes on the Qinghai-Tibet Plateau. Master’s Thesis, Lanzhou Universiry, Lanzhou, China, 2021. [Google Scholar]
- Ren, Z.; Cao, S.; Chen, T.; Zhang, C.; Yu, J. Bacterial functional redundancy and carbon metabolism potentials in soil, sediment, and water of thermokarst landscapes across the Qinghai-Tibet Plateau: Implications for the fate of permafrost carbon. Sci. Total Environ. 2022, 852, 158340. [Google Scholar] [CrossRef]
- Wang, G.; Peng, Y.; Chen, L.; Abbott, B.W.; Ciais, P.; Kang, L.; Liu, Y.; Li, Q.; Peñuelas, J.; Qin, S.; et al. Enhanced response of soil respiration to experimental warming upon thermokarst formation. Nat. Geosci. 2024, 17, 532–538. [Google Scholar] [CrossRef]
- Tian, P.; Cao, L.; Li, J.; Pu, R.; Liu, Y.; Zhang, H.; Wang, C. Ecosystem Stability Assessment of Yancheng Coastal Wetlands, a World Natural Heritage Site. Land 2022, 11, 564. [Google Scholar] [CrossRef]
- Janse, J.H.; van Dam, A.A.; Hes, E.M.A.; de Klein, J.J.M.; Finlayson, C.M.; Janssen, A.B.G.; van Wijk, D.; Mooij, W.M.; Verhoeven, J.T.A. Towards a global model for wetlands ecosystem services. Curr. Opin. Environ. Sustain. 2019, 36, 11–19. [Google Scholar] [CrossRef]
- Zhang, N.; Chen, K.; Wang, S.; Qi, D.; Zhou, Z.; Xie, C.; Liu, X. Dynamic Response of the cbbL Carbon Sequestration Microbial Community to Wetland Type in Qinghai Lake. Biology 2023, 12, 1503. [Google Scholar] [CrossRef]
- Zhang, N.; Chen, K.; Wang, X.; Ji, W.; Yang, Z.; Wang, X.; Li, J. Response Mechanism of cbbM Carbon Sequestration Microbial Community Characteristics in Different Wetland Types in Qinghai Lake. Biology 2024, 13, 333. [Google Scholar] [CrossRef]
- Bai, T.; Wang, P.; Qiu, Y.; Zhang, Y.; Hu, S. Nitrogen availability mediates soil carbon cycling response to climate warming: A meta-analysis. Glob. Chang. Biol. 2023, 29, 2608–2626. [Google Scholar] [CrossRef]
- Nissan, A.; Alcolombri, U.; Peleg, N.; Galili, N.; Jimenez-Martinez, J.; Molnar, P.; Holzner, M. Global warming accelerates soil heterotrophic respiration. Nat. Commun. 2023, 14, 3452. [Google Scholar] [CrossRef]
- Haaf, D.; Six, J.; Doetterl, S. Global patterns of geo-ecological controls on the response of soil respiration to warming. Nat. Clim. Chang. 2021, 11, 623–627. [Google Scholar] [CrossRef]
- Song, X.; Zhu, Y.; Chen, W. Dynamics of the soil respiration response to soil reclamation in a coastal wetland. Sci. Rep. 2021, 11, 2911. [Google Scholar] [CrossRef]
- Chen, G.; Bai, J.; Wang, J.; Liu, Z.; Cui, B. Responses of soil respiration to simulated groundwater table and salinity fluctuations in tidal freshwater, brackish and salt marshes. J. Hydrol. 2022, 612, 128215. [Google Scholar] [CrossRef]
- Han, Y.; Wang, G.; Xiong, L.; Xu, Y.; Li, S. Rainfall effect on soil respiration depends on antecedent soil moisture. Sci. Total Environ. 2024, 926, 172130. [Google Scholar] [CrossRef]
- Zhu, H.; Meason, D.F.; Salekin, S.; Hu, W.; Lad, P.; Jing, Y.; Xue, J. Time stability of soil volumetric water content and its optimal sampling design in contrasting forest catchments. J. Hydrol. 2024, 636, 131344. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Li, H.; Shen, T.; Zhang, X.; Li, J.; Han, G. Grazing decreased soil organic carbon by decreasing aboveground biomass in a desert steppe in Inner Mongolia. J. Environ. Manag. 2023, 347, 119112. [Google Scholar] [CrossRef] [PubMed]
- Martani, E.; Ferrarini, A.; Serra, P.; Pilla, M.; Marcone, A.; Amaducci, S. Belowground biomass C outweighs soil organic C of perennial energy crops: Insights from a long-term multispecies trial. GCB Bioenergy 2021, 13, 459–472. [Google Scholar] [CrossRef]
- Zhao, X.; Zhu, M.; Guo, X.; Wang, H.; Sui, B.; Zhao, L. Organic carbon content and humus composition after application aluminum sulfate and rice straw to soda saline-alkaline soil. Environ. Sci. Pollut. Res. 2019, 26, 13746–13754. [Google Scholar] [CrossRef]
- Donnan, F.G. The Brauner Jubilee Volume. Nature 1925, 115, 975. [Google Scholar] [CrossRef]
- Kucera, C.L.; Kirkham, D.R. Soil Respiration Studies in Tallgrass Prairie in Missouri. Ecology 1971, 52, 912–915. [Google Scholar] [CrossRef]
- Janssens, I.A.; Lankreijer, H.; Matteucci, G.; Kowalski, A.S.; Buchmann, N.; Epron, D.; Pilegaard, K.; Kutsch, W.; Longdoz, B.; Grünwald, T.; et al. Productivity overshadows temperature in determining soil and ecosystem respiration across European forests. Glob. Chang. Biol. 2001, 7, 269–278. [Google Scholar] [CrossRef]
- Jensen, A.E.; Lohse, K.A.; Crosby, B.T.; Mora, C.I. Variations in soil carbon dioxide efflux across a thaw slump chronosequence in northwestern Alaska. Environ. Res. Lett. 2014, 9, 025001. [Google Scholar] [CrossRef]
- Ren, Z.; Luo, W. Metagenomic analysis reveals the diversity and distribution of antibiotic resistance genes in thermokarst lakes of the Yellow River Source Area. Environ. Pollut. 2022, 313, 120102. [Google Scholar] [CrossRef]
- Abbott, B.W.; Jones, J.B. Permafrost collapse alters soil carbon stocks, respiration, CH4, and N2O in upland tundra. Glob. Chang. Biol. 2016, 21, 4570–4587. [Google Scholar] [CrossRef] [PubMed]
- Lin, J. Variations and Influencing Factors of CH4 and CO2 Emissions from Thermokarst Lakes on the Qinghai-Tibet Plateau. Master’s Thesis, LanZhou University, Lanzhou, China, 2021. [Google Scholar]
- Mei, M. Characteristics and Impact Factors of Carbon Release from Thaw Slump Landforms in Qilian Mountains. Master’s Thesis, LanZhou University, Lanzhou, China, 2021. [Google Scholar]
- Junxia, D. Study on Soil Respiration in the Freeze-ThawErosion Area of Alpine Marsh Meadows on the Qinghai-Tibet Plateau- Wayan Mountain as an Example. Master’s Thesis, Qinghai Normal University, Xining, China, 2020. [Google Scholar]
- Chen, S.; Zhou, Z.; Yan, L.; Li, B. Quantitative Evaluation of Ecosystem Health in a Karst Area of South China. Sustainability 2016, 8, 975. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, L.; Guo, R.; Huang, R.; Zhou, Z.; Niu, F.; Jiao, Z. Interaction of permafrost degradation and thermokarst lakes in the Qinghai–Tibet Plateau. Geomorphology 2023, 425, 108582. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, G.; Du, H.; Berninger, F.; Mao, F.; Li, X.; Chen, L.; Cui, L.; Li, Y.; Zhu, D. Soil respiration of a Moso bamboo forest significantly affected by gross ecosystem productivity and leaf area index in an extreme drought event. PeerJ 2018, 6, e5747. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Zhu, S.; Feng, J.; Shui, W. Slope aspect affects the soil microbial communities in karst tiankeng negative landforms. BMC Ecol. Evol. 2022, 22, 54. [Google Scholar] [CrossRef] [PubMed]
- Długosz, J.; Piotrowska-Długosz, A.; Breza-Boruta, B. The effect of differences in soil water content on microbial and enzymatic properties across the soil profiles. Ecohydrol. Hydrobiol. 2023, 1642–3593. [Google Scholar] [CrossRef]
- Suh, S.; Park, S.; Shim, K.; Yang, B.; Kim, T. The Effect of Rain Fall Event on CO2 Emission in Pinus koraiensis Plantation in Mt. Taehwa. Environ. Biol. Res. 2014, 32, 389–394. [Google Scholar] [CrossRef]
- Zhang, M.; Niu, Y.; Wang, W.; Bai, S.; Luo, H.; Tang, L.; Chen, F.; Xu, Z.; Guo, X. Responses of microbial function, biomass and heterotrophic respiration, and organic carbon in fir plantation soil to successive nitrogen and phosphorus fertilization. Appl. Microbiol. Biotechnol. 2021, 105, 8907–8920. [Google Scholar] [CrossRef] [PubMed]
- Prakash, O. Lack of kinship with anaerobes is a kind of short-sightedness of agricultural and environmental microbiologists. Environ. Microbiol. Rep. 2022, 14, 330–332. [Google Scholar] [CrossRef]
- Hall, S.J.; Huang, W.; Napieralski, S.A.; Roden, E. Shared Microbial Taxa Respond Predictably to Cyclic Time-Varying Oxygen Limitation in Two Disparate Soils. Front Microbiol 2022, 13, 866828. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecological Indicators | T (°C) | VWC (m3/m3) | RS (μmol·m−2·s−1) | Above-Ground Biomass (g/m2) |
|---|---|---|---|---|
| N | 9.85 ± 1.36 a | 0.42 ± 0.04 b | 4.52 ± 0.36 a | 257.72 ± 31.23 a |
| Thermokarst | 11.03 ± 1.69 a | 0.49 ± 0.03 a | 3.59 ± 0.49 b | 186.34 ± 18.31 b |
| Ecological Indicators | T (°C) | VWC (m3/m3) | RS (μmol·m−2·s−1) | Q10 | ||||
|---|---|---|---|---|---|---|---|---|
| Day | Night | Day | Night | Day | Night | Day | Night | |
| N | 10.05 ± 2.73a | 9.01 ± 2.35a | 0.416 ± 0.021b | 0.413 ± 0.011b | 4.77 ± 0.41a | 4.15 ± 0.31a | 2.34 ± 0.36a | 1.81 ± 0.25b |
| Thermokarst | 11.11 ± 2.81a | 10.52 ± 2.62a | 0.476 ± 0.027a | 0.473 ± 0.022a | 4.09 ± 0.37a | 2.82 ± 0.22b | 2.03 ± 0.27a | 3.32 ± 0.38a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, Y.; Chen, K.; Ji, W.; Yang, Y. The Response of Soil Respiration to Temperature and Humidity in the Thermokarst Depression Zone of the Headwater Wetlands of Qinghai Lake. Biology 2024, 13, 437. https://doi.org/10.3390/biology13060437
Mao Y, Chen K, Ji W, Yang Y. The Response of Soil Respiration to Temperature and Humidity in the Thermokarst Depression Zone of the Headwater Wetlands of Qinghai Lake. Biology. 2024; 13(6):437. https://doi.org/10.3390/biology13060437
Chicago/Turabian StyleMao, Yahui, Kelong Chen, Wei Ji, and Yanli Yang. 2024. "The Response of Soil Respiration to Temperature and Humidity in the Thermokarst Depression Zone of the Headwater Wetlands of Qinghai Lake" Biology 13, no. 6: 437. https://doi.org/10.3390/biology13060437
APA StyleMao, Y., Chen, K., Ji, W., & Yang, Y. (2024). The Response of Soil Respiration to Temperature and Humidity in the Thermokarst Depression Zone of the Headwater Wetlands of Qinghai Lake. Biology, 13(6), 437. https://doi.org/10.3390/biology13060437