
Pan-Evo: The Evolution of Information and Biology’s Part in This


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
- Innovation (Section 3.1 and Section 3.2).
- Transmission and replication, including the random processes therein (Section 4.1 and Section 4.2).
- Adaptation and processes such as selection that often produce adaptation (Section 5.1 and Section 5.2).
- Movement (Section 6.1 and Section 6.2).
2. Information, Life, and Intelligence
- oooooooooooooo Entropy = zero
- ooooooohhhhhhh Entropy = 0.69
- oooyhaerlelhwu Entropy = 2.11
- hellohowareyou Entropy = 2.11
3. Innovation
3.1. Innovation in Biology
3.2. Innovation Outside of Biology
4. Transmission and Replication
4.1. Transmission and Replication in Biology
4.2. Transmission and Replication Outside Biology
5. Adaptation
5.1. Adaptation in Biology
5.2. Adaptation Outside Biology
6. Movement
6.1. Movement in Biology
6.2. Movement Outside Biology
7. Speciation
7.1. Speciation in Biology
7.2. Speciation Outside Biology
8. Integration of Non-Biological and Biological
9. Potential Benefits and Harms
9.1. Possible Benefits to Biology, including Humans
9.2. Possible Threats to Biology, including Humans
10. AI, Competition, and Panspeciation
11. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Anonymous. AI Special Issue: A machine-intelligent world. Science 2023, 381, 136–137. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. EDITORIAL: Stop talking about tomorrow’s AI doomsday when AI poses risks today. Nature 2023, 618, 885–886. [Google Scholar] [CrossRef] [PubMed]
- Federspiel, F.; Mitchell, R.; Asokan, A.; Umana, C.; McCoy, D. Threats by artificial intelligence to human health and human existence. BMJ Glob. Health 2023, 8, e010435. [Google Scholar] [CrossRef] [PubMed]
- Rich, A.S.; Gureckis, T.M. Lessons for artificial intelligence from the study of natural stupidity. Nat. Mach. Intell. 2019, 1, 174–180. [Google Scholar] [CrossRef]
- Sherwin, W.B. Singularity or Speciation? A comment on Lazar S & Nelson A. 2023. AI safety on whose terms? Science 2023, 381, 138. [Google Scholar]
- Omohundro, S.M. The Basic AI Drives. Artif. Gen. Intell. 2008, 171, 483–492. [Google Scholar]
- Yudkowsky, E.; Salamon, E.A.; Shulman, C.; Kaas, S.; McCabe, T.; Nelson, R. Reducing Long-Term Catastrophic Risks from Artificial Intelligence; Machine Intelligence Research Institute: San Francisco, CA, USA, 2010. [Google Scholar]
- O’Connor, M.I.; Pennell, M.W.; Altermatt, F.; Matthews, B.; Melian, C.J.; Gonzalez, A. Principles of ecology revisited: Integrating information and ecological theories for a more unified science. Front. Ecol. Evol. 2019, 7, 219. [Google Scholar] [CrossRef]
- Schrodinger, E. What Is Life? Cambridge University Press: Cambridge, UK, 1944. [Google Scholar]
- Margalef, R. Diversidad de especies en las comunidades naturales. Publicaciones Inst. Biol. Apl. 1951, 9, 5–27. [Google Scholar]
- Branson, H.R. Information theory and the structure of proteins. In Information Theory in Biology; Quastler, R., Ed.; University of Illinois Press: Urbana, IL, USA, 1953; pp. 84–104. [Google Scholar]
- Lewontin, R.C. The apportionment of human diversity. Evol. Biol. 1972, 6, 381–398. [Google Scholar]
- Ewens, W. Mathematical Population Genetics; Springer: Berlin/Heidelberg, Germany, 1979. [Google Scholar]
- Ewens, W.J. The sampling theory of selectively neutral alleles. Theor. Popul. Biol. 1972, 3, 87–112. [Google Scholar] [CrossRef]
- Caswell, H. Community structure: A neutral model analysis. Ecol. Monogr. 1976, 46, 327–354. [Google Scholar] [CrossRef]
- Sherwin, W.B.; Chao, A.; Jost, L.; Smouse, P.E. Information Theory Broadens the Spectrum of Molecular Ecology and Evolution. Trends Ecol. Evol. 2017, 32, 948–963, Erratum in Trends Ecol. Evol. 2021, 36, 955–956. [Google Scholar] [CrossRef] [PubMed]
- Sherwin, W.B.; Jabot, F.; Rush, R.; Rossetto, M. Measurement of biological information with applications from genes to landscapes. Mol. Ecol. 2006, 15, 2857–2869. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, G.D.; Jabot, F.; Gunn, M.R.; Sherwin, W.B. Predicting Shannon’s information for genes in finite populations: New uses for old equations. Conserv. Genet. Resour. 2020, 12, 245–255. [Google Scholar] [CrossRef]
- Sherwin, W.B. Entropy, or Information, Unifies Ecology and Evolution. Entropy 2018, 20, 727. [Google Scholar] [CrossRef]
- Buddle, C.M.; Beguin, J.; Bolduc, E.; Mercardo, A.; Sackett, T.E.; Selby, R.D.; Varady-Szabo, H.; Zeran, R.M. The importance and use of taxon sampling curves for comparative biodiversity research with forest arthropod assemblages. Can. Entomol. 2004, 137, 120–127. [Google Scholar] [CrossRef]
- Vellend, M. The Theory of Ecological Communities (MPB-57); Princeton University Press: Princeton, NJ, USA, 2016. [Google Scholar]
- Muthukrishna, M. A Theory of Everyone: Who We Are, How We Got Here, and Where We’re Going; John Murray Press: London, UK, 2024. [Google Scholar]
- DeLong, J.P.; O’Connor, M.I.; Savage, V.; Luhring, T.; Schaum, C.-E.; Sears, M. Beyond individual, population, and community: Considering information, cell number, and energy flux as fundamental dimensions of life across scales. Ideas Ecol. Evol. 2023, 16, 1–9. [Google Scholar] [CrossRef]
- Hubbell, S.P. The Unified Neutral Theory of Biodiversity and Biogeography; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Rosindell, J.; Cornell, S.J.; Hubbell, S.P.; Etienne, R.S. Protracted speciation revitalizes the neutral theory of biodiversity. Ecol. Lett. 2010, 13, 716–727. [Google Scholar] [CrossRef]
- Overcast, I.; Ruffley, M.; Rosindell, J.; Harmon, L.; Borges, P.A.V.; Emerson, B.C.; Etienne, R.S.; Gillespie, R.; Krehenwinkel, H.; Mahler, D.L.; et al. A unified model of species abundance, genetic diversity, and functional diversity reveals the mechanisms structuring ecological communities. Mol. Ecol. Resour. 2021, 21, 2782–2800. [Google Scholar] [CrossRef]
- Rollins, L.A.; Woolnough, A.P.; Fanson, B.G.; Cummins, M.L.; Crowley, T.M.; Wilton, A.N.; Sinclair, R.; Butler, A.; Sherwin, W.B. Selection on mitochondrial variants occurs between and within individuals in an expanding invasion. Mol. Biol. Evol. 2016, 33, 995–1007. [Google Scholar] [CrossRef]
- Lewontin, R.C. The units of selection. Annu. Rev. Ecol. Syst. 1970, 1, 1–18. [Google Scholar] [CrossRef]
- Licklider, J.C.R. Man-Computer Symbiosis. IRE Trans. Hum. Factors Electron. 1960, HFE-1, 4–11. [Google Scholar] [CrossRef]
- Halpin, H. The Co-Evolution of the Extended Mind and the Internet. Proceedings 2022, 81, 148. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication.; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Trifonov, E.N. Vocabulary of Definitions of Life Suggests a Definition. J. Biomol. Struct. Dyn. 2011, 29, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Charlat, S.; Ariew, A.; Bourrat, P.; Ferreira Ruiz, M.; Heams, T.; Huneman, P.; Krishna, S.; Lachmann, M.; Lartillot, N.; Le Sergeant d’Hendecourt, L.; et al. Natural Selection beyond Life? A Workshop Report. Life 2021, 11, 1051. [Google Scholar] [CrossRef] [PubMed]
- England, J.L. Statistical physics of self-replication. J. Chem. Phys. 2013, 139, 121923. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Czégel, D.; Lachmann, M.; Kempes, C.P.; Walker, S.I.; Cronin, L. Assembly theory explains and quantifies selection and evolution. Nature 2023, 622, 321–328. [Google Scholar] [CrossRef]
- Liu, Y.; Mathis, C.; Bajczyk, M.D.; Marshall, S.M.; Wilbraham, L.; Cronin, L. Exploring and mapping chemical space with molecular assembly trees. Sci. Adv. 2021, 7, eabj2465. [Google Scholar] [CrossRef]
- Finnila, S.; Lehtonen, M.S.; Majamaa, K. Phylogenetic Network for European mtDNA. Am. J. Hum. Genet. 2001, 68, 1475–1484. [Google Scholar] [CrossRef]
- Nisbett, R.E.; Aronson, J.; Blair, C.; Dickens, W.; Flynn, J.; Halpern, D.F.; Turkheimer, E. Intelligence: New Findings and Theoretical Developments. Am. Psychol. 2012, 67, 130–159. [Google Scholar] [CrossRef]
- Soreq, E.; Violante, I.R.; Daws, R.E.; Hampshire, A. Neuroimaging evidence for a network sampling theory of individual differences in human intelligence test performance. Nat. Commun. 2021, 12, 2072. [Google Scholar] [CrossRef] [PubMed]
- Oxford-Dictionary. Available online: https://www.bing.com/search?q=intelligence+definition+oxford+dictionary&qs=n&form=QBRE&sp=-1&ghc=2&lq=0&pq=intelligence+definition+oxford+dictionary&sc=7-41 (accessed on 21 June 2024).
- Walsh, T. Machines Behaving Badly: The Morality of AI; LaTrobe University Press: Melbourne, Australia, 2022. [Google Scholar]
- Danchin, E. Avatars of information: Towards an inclusive evolutionary synthesis. Trends Ecol. Evol. 2013, 28, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Bonduriansky, R. Rethinking heredity, again. Trends Ecol. Evol. 2012, 27, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Coyne, J.; Orr, H.A. Speciation; Sinauer: Sunderland, MA, USA, 2004. [Google Scholar]
- Rebeck, J. Synthetic Self-Replicating Molecules. Sci. Am. 1994, 271, 48–55. [Google Scholar] [CrossRef]
- Hamblin, S. On the practical usage of genetic algorithms in ecology and evolution. Methods Ecol. Evol. 2013, 4, 184–194. [Google Scholar] [CrossRef]
- George, A.; Walsh, T. Can AI invent? Nat. Mach. Intell. 2022, 4, 1057–1060. [Google Scholar] [CrossRef]
- Danchin, E.; Charmantier, A.; Champagne, F.; Mesoudi, A.; Pujol, B.; Blanchet, S. Beyond DNA: Integrating inclusive inheritance into an extended theory of evolution. Nat. Rev. Genet. 2011, 12, 475–486. [Google Scholar] [CrossRef]
- Oldroyd, B. Beyond DNA: How Epigenetics is Transforming our Understanding of Evolution; Melbourne University Press: Melbourne, Australia, 2023. [Google Scholar]
- Piñero, J.; Solé, R. Nonequilibrium Entropic Bounds for Darwinian Replicators. Entropy 2018, 20, 98. [Google Scholar] [CrossRef]
- Bissette, A.J.; Fletcher, S.P. Mechanisms of Autocatalysis. Angew. Chem. Int. Ed. 2013, 52, 12800–12826. [Google Scholar] [CrossRef]
- Knight, R.D.; Landweber, L.F. Rhyme or reason: RNA-arginine interactions and the genetic code. Chem. Biol. 1998, 5, R215–R220. [Google Scholar] [CrossRef]
- Goodfellow, I.J.; Pouget-Abadie, J.; Mirza, M.; Xu, B.; Warde-Farley, D.; Ozair, S.; Courville, A.; Bengio, Y. Generative Adversarial Networks. arXiv 2014, arXiv:1406.2661v1 [stat.ML]. [Google Scholar] [CrossRef]
- Brady, S.P.; Daniel, I.; Bolnick, D.I.; Angert, A.L.; Gonzalez, A.; Barrett, R.D.H.; Crispo, E.; Derry, A.M.; Eckert, C.G.; Fraser, D.J.; et al. Causes of maladaptation. Evol. Appl. 2019, 12, 1229–1242. [Google Scholar] [CrossRef] [PubMed]
- Good, I.J. A little learning can be dangerous. Br. J. Phil. Sci. 1974, 25, 340–342. [Google Scholar] [CrossRef]
- Frank, S.A. The Price Equation Program: Simple Invariances Unify Population Dynamics, Thermodynamics, Probability, Information and Inference. Entropy 2018, 20, 978. [Google Scholar] [CrossRef] [PubMed]
- Day, T. Information entropy as a measure of genetic diversity and evolvability in colonization. Mol. Ecol. 2015, 24, 2073–2083. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudık, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Yan, W.Q. Computational Methods for Deep Learning: Theory, Algorithms, and Implementations; Springer Nature: Vienna, Austria, 2023. [Google Scholar]
- Turing, A. Computing Machinery and Intelligence. Mind 1950, LIX, 433–460. [Google Scholar] [CrossRef]
- Lavoipierre, A. A Twist in the Legal Fight between X and eSafety. Available online: https://www.abc.net.au/news/2024-05-28/twist-in-legal-fight-between-x-esafety-google-ai-glue-and-rocks/103900668 (accessed on 5 June 2024).
- Bajic, D. Information Theory, Living Systems, and Communication Engineering. Entropy 2024, 26, 430. [Google Scholar] [CrossRef] [PubMed]
- Ivaniš, P.; Milojkovic, J.; Blagojevic, V.; Brkic, S. Capacity Analysis of Hybrid Satellite–Terrestrial Systems with Selection Relaying. Entropy 2024, 26, 419. [Google Scholar] [CrossRef]
- Brooks, R.C. How Might Artificial Intelligence Influence Human Evolution? Q. Rev. Biol. 2024; in press. [Google Scholar]
- Dingle, H.; Alistair Drake, V.A. What Is Migration? BioScience 2007, 57, 113–121. [Google Scholar] [CrossRef]
- Lam, K.-Y.; Lou, Y. Introduction to Reaction-Diffusion Equations: Theory and Applications to Spatial Ecology and Evolutionary Biology; Springer Nature: Cham, Switzerland, 2022. [Google Scholar]
- Duran-Nebreda, S.; Pla, J.; Vidiella, B.; Piñero, J.; Conde-Pueyo, N.; Sole, R. Synthetic Lateral Inhibition in Periodic Pattern Forming Microbial Colonies. ACS Synth. Biol. 2021, 10, 277–285. [Google Scholar] [CrossRef] [PubMed]
- FitzHugh, R. Impulses and Physiological States in Theoretical Models of Nerve Membrane. Biophys. J. 1961, 1, 445–466. [Google Scholar] [CrossRef]
- De Queiroz, K. Ernst Mayr and the modern concept of species. Proc. Natl. Acad. Sci. USA 2005, 102, 6600–6607. [Google Scholar] [CrossRef]
- Costello, M.J.; May, R.M.; Stork, N.E. Can We Name Earth’s Species Before They Go Extinct? Science 2013, 339, 413–416. [Google Scholar] [CrossRef]
- Edmands, S. Does parental divergence predict reproductive compatibility? Trends Ecol. Evol. 2002, 17, 520. [Google Scholar] [CrossRef]
- Moritz, C. Defining “evolutionarily significant units” for conservation. Trends Ecol. Evol. 1994, 9, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Moritz, C. Conservation units and translocations: Strategies for conserving evolutionary processes. Hereditas 1999, 130, 217–228. [Google Scholar] [CrossRef]
- Lan, R.; Reeves, P. When does a clone deserve a name? A perspective on bacterial species based on population genetics. Trends Microbiol. 2001, 9, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Carr, N. How many computers does the world need? Fewer than you think. In Guardian; Guardian: London, UK, 2008. [Google Scholar]
- Ji, Z.; Lee, N.; Frieske, R.; Yu, T.; Su, D.; Xu, Y.; Ishii, E.; Bang, Y.; Dai, W.; Madotto, A.; et al. Survey of Hallucination in Natural Language Generation. ACM Comput. Surv. 2023, 55, 248. [Google Scholar] [CrossRef]
- Anonymous. Compatibility. Available online: https://www.techtarget.com/whatis/definition/compatibility (accessed on 9 May 2024).
- von Neumann, J.; Burks, A.W.e. Theory of Self-Reproducing Automata; University of Illinois Press: Urbana, IL, USA; London, UK, 1966. [Google Scholar]
- Sole, R. Synthetic transitions: Towards a new synthesis. Phil. Trans. Roy. Soc. B. 2016, 371, 20150438. [Google Scholar] [CrossRef]
- Halliburton, R. Introduction to Population Genetics; Pearson: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Wright, S. Size of population and breeding structure in relation to evolution. Proc. Natl. Acad. Sci. USA 1938, 24, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Kleidon, A. Life, hierarchy, and the thermodynamic machinery of planet Earth. Phys. Life Rev. 2010, 7, 424–460. [Google Scholar] [CrossRef] [PubMed]
- Wicken, J.S. Entropy and information: Suggestions for common language. Philos. Sci. 1987, 54, 176–193. [Google Scholar] [CrossRef]
- Baravalle, L.; Luque, V.J. Towards a Pricean foundation for cultural evolutionary theory. Theoria 2022, 37, 209–231. [Google Scholar] [CrossRef]
- Hanopolskyi, A.I.; Smaliak, V.A.; Novichkov, A.I.; Semenov, S.N. Autocatalysis: Kinetics, Mechanisms and Design. ChemSystemsChem 2021, 3, e2000026. [Google Scholar] [CrossRef]
- Lincoln, T.A.; Joyce, G.F. Self-sustained replication of an RNA enzyme. Science 2009, 323, 1229–1232. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhang, X.; Shi, J.; Yan, M.; Zhou, T. Origins and evolving functionalities of tRNA-derived small RNAs. Trends Biochem. Sci. 2021, 46, 790–804. [Google Scholar] [CrossRef] [PubMed]
- Adamski, P.; Eleveld, M.; Sood, A.; Ku, Á.; Szilágyi, A.; Czárán, T.; Szathmáry, E.; Otto, S. From self-replication to replicator systems en route to de novo life. Nat. Rev. 2020, 4, 386–403. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Liu, J. Catalytic Nucleic Acids: Biochemistry, Chemical Biology, Biosensors, and Nanotechnology. iScience 2020, 23, 100815. [Google Scholar] [CrossRef]
- Lazar, S.; Nelson, A. AI safety on whose terms? Science 2023, 381, 138. [Google Scholar] [CrossRef]
- Richerson, P.J.; Boyd, R. Not by Genes Alone: How Culture Transformed Human Evolution; University of Chicago Press: Chicago, IL, USA, 2006. [Google Scholar]
- Carroll, S.P.; Jørgensen, P.S.; Kinnison, M.T.; Bergstrom, C.T.; Denison, R.F.; Gluckman, P.; Smith, T.B.; Strauss, S.Y.; Tabashnik, B.E. Applying evolutionary biology to address global challenges. Science 2014, 346, 1245993. [Google Scholar] [CrossRef] [PubMed]
- von Kodolitsch, Y.; Berger, J.; Rogan, P.K. Predicting severity of haemophilia A and B splicing mutations by information analysis. Haemophilia 2006, 12, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Woolnough, A.P.; Hollenberg, L.C.L.; Cassey, P.; Prowse, T.A.A. Quantum computing: A new paradigm for ecology. Trends Ecol. Evol. 2023, 38, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, A.; van Schaik, C.P.; Isler, K. Energetics and the evolution of human brain size. Nature 2011, 480, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Mitteroecker, P.; Huttegger, S.M.; Fischer, B.; Pavlicev, M. Cliff-edge model of obstetric selection in humans. Proc. Natl. Acad. Sci. USA 2016, 113, 14680–14685. [Google Scholar] [CrossRef] [PubMed]
- Dunsworth, H.M. Thank your intelligent mother for your big brain. Proc. Natl. Acad. Sci. USA 2016, 113, 6816–6818. [Google Scholar] [CrossRef] [PubMed]
- Van Valen, L. Ecological Species, multispecies, and oaks. Taxon 1976, 25, 233–239. [Google Scholar] [CrossRef]
- Hart, S.P.; Schreiber, S.J.; Levine, J.M. How variation between individuals affects species coexistence. Ecol. Lett. 2016, 19, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Hart, S.P.; Turcotte, M.M.; Levine, J.M. Effects of rapid evolution on species coexistence. Proc. Natl. Acad. Sci. USA 2019, 116, 2112–2117. [Google Scholar] [CrossRef]

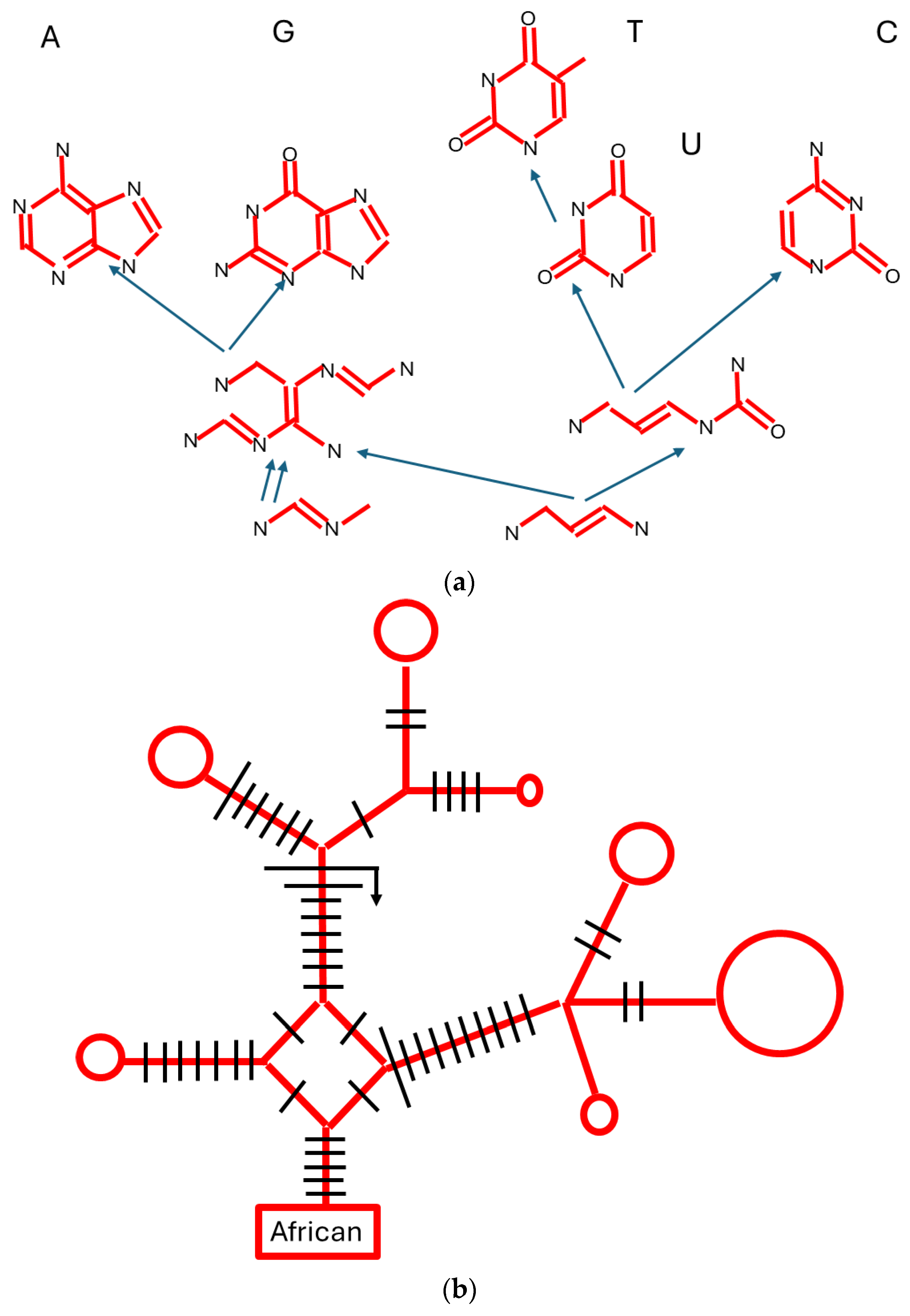
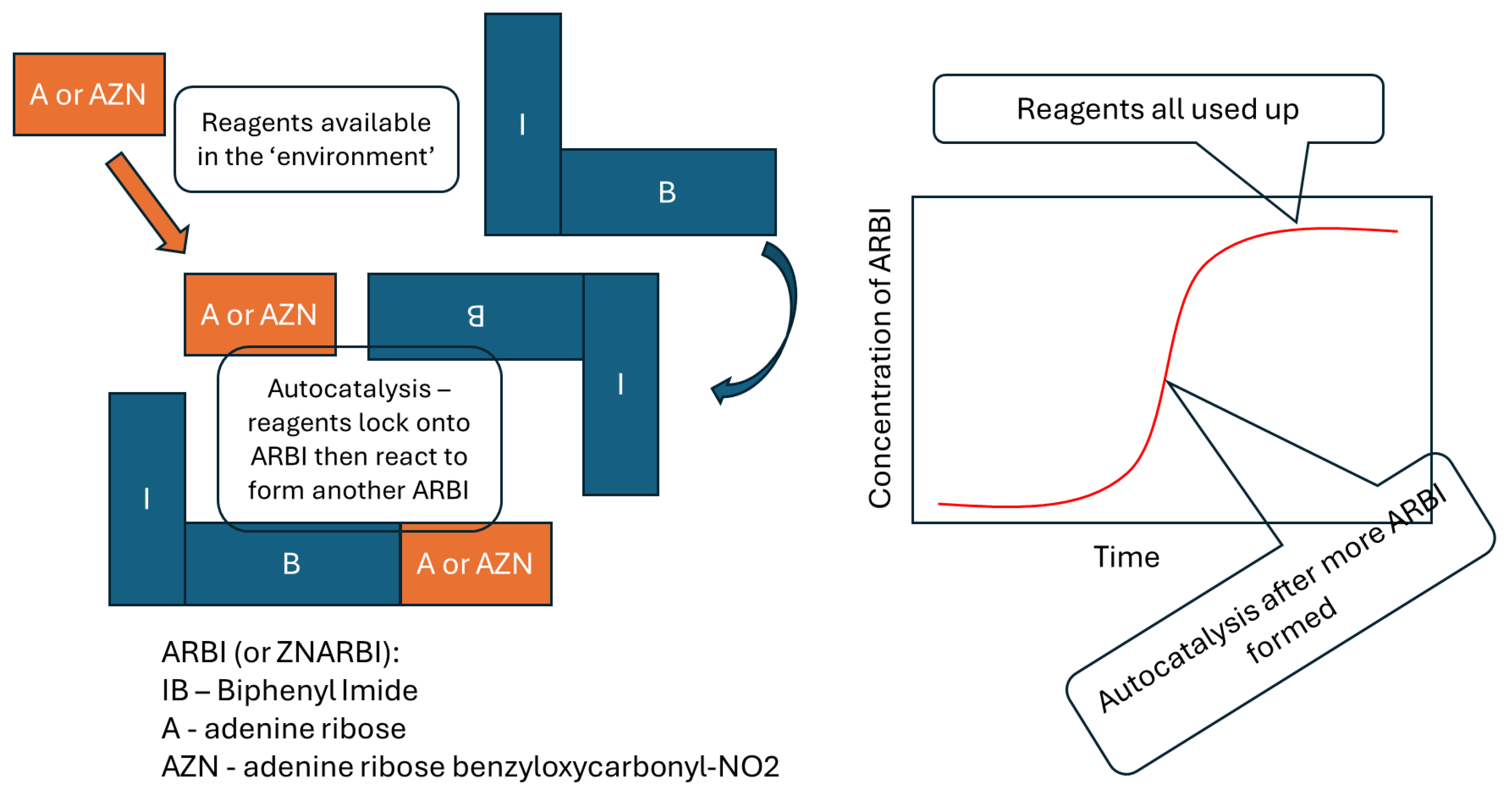
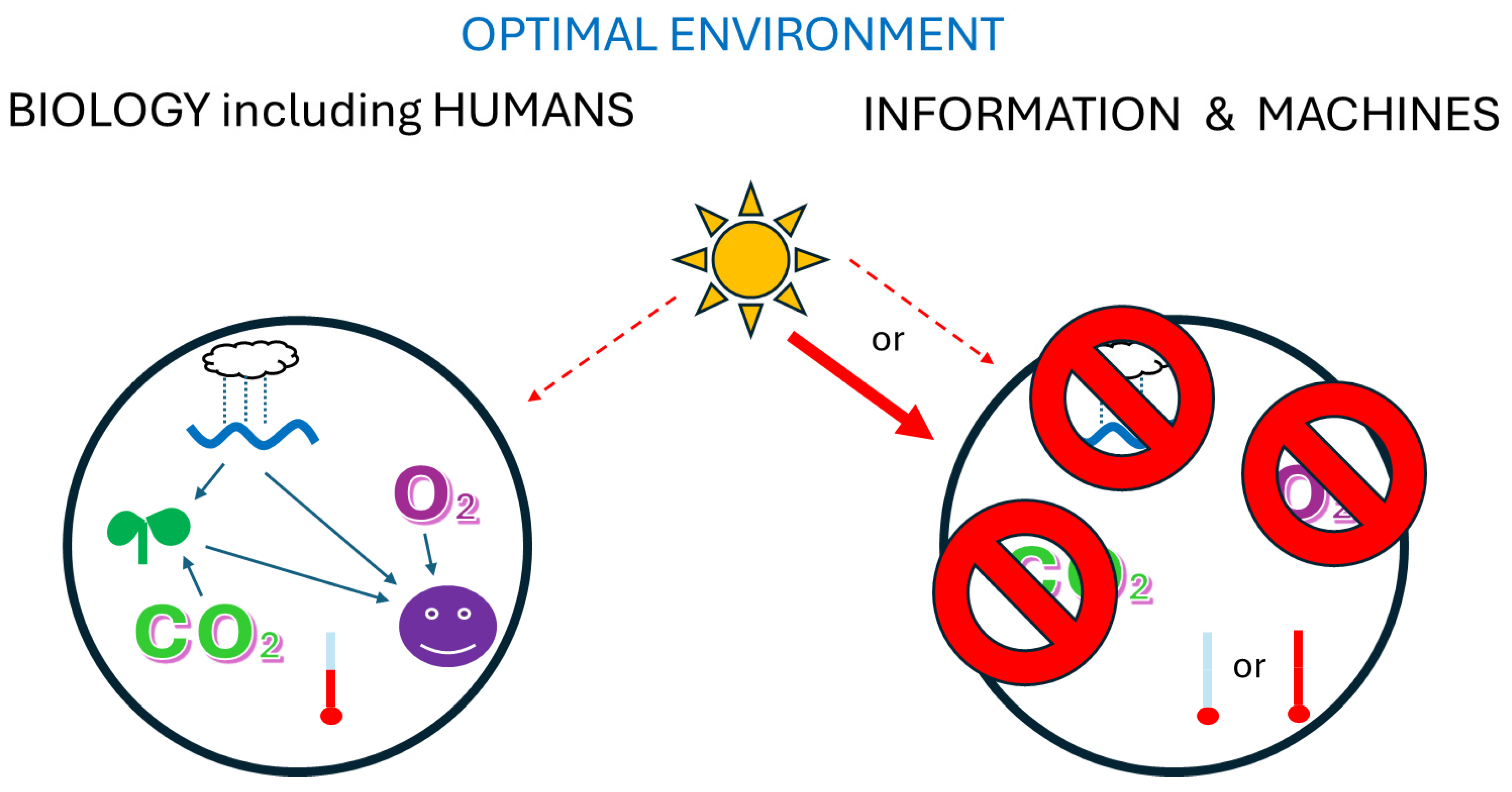
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sherwin, W.B. Pan-Evo: The Evolution of Information and Biology’s Part in This. Biology 2024, 13, 507. https://doi.org/10.3390/biology13070507
Sherwin WB. Pan-Evo: The Evolution of Information and Biology’s Part in This. Biology. 2024; 13(7):507. https://doi.org/10.3390/biology13070507
Chicago/Turabian StyleSherwin, William B. 2024. "Pan-Evo: The Evolution of Information and Biology’s Part in This" Biology 13, no. 7: 507. https://doi.org/10.3390/biology13070507
APA StyleSherwin, W. B. (2024). Pan-Evo: The Evolution of Information and Biology’s Part in This. Biology, 13(7), 507. https://doi.org/10.3390/biology13070507